

Палеонтологический журнал, 2019, № 3, стр. 88-96
Новый примитивный тероцефал (Therocephalia, Theromorpha) из средней перми Восточной Европы
Ю. А. Сучкова a, *, В. К. Голубев a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
* E-mail: js@paleo.ru
** E-mail: vg@paleo.ru
Поступила в редакцию 04.12.2018
После доработки 11.12.2018
Принята к публикации 25.12.2018
Аннотация
По материалам из типового местонахождения сундырского комплекса тетрапод Сундырь-1 (Республика Марий Эл, Россия) описан самый крупный среднепермский тероцефал Восточной Европы, новый представитель сем. Scylacosauridae – Julognathus crudelis gen. et sp. nov. В сундырском сообществе тетрапод юлогнатус занимал нишу крупнейшего хищника, образуя трофическую пару с растительноядным тапиноцефалом.
ВВЕДЕНИЕ
В 2010 г. был открыт новый этап в развитии фауны пермских тетрапод Восточной Европы – сундырский комплекс (Голубев и др., 2011). Комплекс датируется началом позднесеверодвинского века (раннепутятинское время) (Голубев и др., 2015). Он является переходным между среднепермским диноцефаловым и позднепермским териодонтовым суперкомплексами, характеризуя заключительную стадию развития диноцефаловой фауны на территории Восточно-Европейской платформы накануне северодвинского (позднегваделупского) экологического кризиса (Голубев, Сенников, 2011, 2014; Golubev, 2015; Arefiev et al., 2015; Сенников, Голубев, 2017; Голубев, Буланов, 2018).
Ранее были описаны представители водного и субдоминантного блоков сундырского сообщества11 (Буланов, Голубев, 2011; Куркин, 2017; Голубев, Буланов, 2018; Bulanov, Golubev, 2018; Ульяхин, 2018). Сундырское водное сообщество довольно разнообразно. Его формируют хрониозухиды Suchonica vladimiri Golubev, темноспондилы Dvinosaurus aff. primus Amalitzky, сеймуриаморфы Microphon exiguus Ivachnenko и Leptorophа aff. talonophora (Tchudinov), энозухидные антракозавры Enosuchus alveolatus Bulanov et Golubev. Из субдоминантного блока сундырского сообщества пока известны только галеопиды Parasuminia ivachnenkoi Kurkin. Хрониозухиды, двинозавры, микрофоны и галеопиды сближают сундырскую фауну с более молодыми комплексами териодонтового суперкомплекса. На территории Восточной Европы эти группы не известны в более древних комплексах, чем сундырский (Сенников, Голубев, 2017). Лепторофы и энозухиды, наоборот, распространены исключительно в комплексах диноцефалового суперкомплекса (Сенников, Голубев, 2017).
Доминантный блок сундырского сообщества до сих пор подробно не описан. В этом блоке нишу крупного фитофага, несомненно, занимали тапиноцефалы, возможно, близкие к ишеевским улемозавридам. Об их присутствии в сундырской фауне свидетельствуют многочисленные находки изолированных зубов характерной формы, неизвестной более ни у одной другой группы терапсид, а также крупных челюстных костей с типичным для тапиноцефалов постепенным уменьшением размеров зубов от симфиза к углам пасти и отсутствием морфологически выраженных клыков. В отношении же хищной компоненты доминантного блока сундырского сообщества долгое время не было полной определенности. Это связано с характером сохранности материала в местонахождении Сундырь-1. В этом местонахождении остатки тетрапод представлены исключительно изолированными костями, зачастую сильно поломанными и окатанными, что затрудняет их точную диагностику.
На первом этапе изучения местонахождения предполагалось, что хищниками в доминантном блоке были антеозавриды cf. Titanophoneus и сиодонтиды cf. Syodon (Kurkin, 2010; Голубев и др., 2011; Буланов, Голубев, 2011; Голубев, Сенников, 2011, 2014; Голубев, 2012, 2018). В таком составе сундырское доминантное сообщество полностью соответствовало доминантному сообществу более древнего (позднеуржумско-раннесеверодвинского) ишеевского комплекса. Позднее, по мере поступления нового материала, присутствие хищных диноцефалов в сундырском комплексе не подтвердилось, и было выдвинуто предположение, что трофическую пару растительноядному тапиноцефалу в этой фауне составляли горгонопии (Голубев и др., 2015; Golubev, 2015; Сенников, Голубев, 2017; Suchkova, 2017). Основано оно было на том, что некоторые зубы, а также покровные кости нижней челюсти и предглазничной части черепа хищных терапсид, обнаруженные в местонахождении Сундырь-1, отличались характерной для горгонопий морфологией (Сучкова, 2017а–г, 2018). Новые материалы, собранные в результате раскопок этого местонахождения в 2016–2018 гг., заставили пересмотреть и эти представления. В частности, были найдены крупные parietalia (экз. № 5388/421), узкие и сросшиеся по сагиттальному шву, что типично для тероцефалов и цинодонтов, но никогда не наблюдается у горгонопий. Также было найдено pterygoideum (экз. № 5388/270), передне-верхний край которого не имеет следов контакта с ectopterygoideum, что указывает на наличие подглазничных окон, присутствующих только у тероцефалов.
Таким образом, в сундырской коллекции были обнаружены остатки крупных тероцефалов. Учитывая новые данные, мы критически пересмотрели всю коллекцию сундырских хищных терапсид. В результате оказалось, что остатки, которые ранее определялись как принадлежащие горгонопиям, характеризуются признаками либо типичными только для тероцефалов, либо характерными как для горгонопий, так и для примитивных тероцефалов. В частности, в коллекции хорошо представлены зубные кости и имеется одна пластинчатая кость. Spleniale (экз. № 5388/127) найдено в непосредственной близости от dentale (экз. № 5388/126), имеет подходящие ему размеры и отличается от аналогичных костей горгонопий плоской формой без медиального выступа в симфизной части. На всех обнаруженных зубных костях сундырских териодонтов, если это позволяет сохранность образцов, прослеживаются морфологические признаки, характерные для примитивных тероцефалов (van den Heever, 1987, 1994; Татаринов, 2008; Abdala et al., 2008): слабая выраженность симфизных поверхностей (рыхлый симфиз) и отсутствие за ними типичного для горгонопий вдавления для вмещения передней части spleniale; покатый, вытянутый, отнесенный далеко назад подбородочный выступ; три нижнечелюстных резца (у горгонопий четыре); углообразный изгиб задне-нижнего края зубной кости; венечный отросток с широким терминальным краем, характерным для тероцефалов, но не для горгонопий.
Таким образом, было установлено, что доминирующими хищниками в сундырском комплексе были не горгонопии, а тероцефалы, по ряду особенностей морфологии черепа напоминающие горгонопий. Это морфологическое сходство с горгонопиями является отличительной особенностью многих среднепермских тероцефалов. Их называют базальными, или ранними тероцефалами (van den Heever, 1987, 1994). По Ю. ван ден Хиверу, эта группа обособляется от прочих тероцефалов наличием таких признаков, как зазубренные челюстные зубы, заднелобные кости, три нижних резца, узкие несрастающиеся сошники, обычно крупные размеры и клыки (van den Heever, 1987, 1994). Группа считается примитивной, и в составе двух семейств – короткомордых Lycosuchidae и длинномордых Scylacosauridae – противопоставляется прогрессивным Eutherocephalia (van den Heever, 1994; Abdala et al., 2008, 2014).
На территории России ранние тероцефалы долгое время были практически неизвестны (Ivakhnenko, 2011). В сборах из медистых песчаников остатки тероцефалов определены лишь предположительно по единичным находкам костей посткраниального скелета (Ефремов, 1954, с. 325–326). В ишеевском комплексе по немногочисленным фрагментарным остаткам описан тероцефал Porosteognathus efremovi Vjuschkov (Вьюшков, 1952), который по наличию зазубренности на кантах челюстных зубов должен быть отнесен к ранним тероцефалам. Еще более фрагментарны находки тероцефалов в местонахождении Малая Кинель (Оренбургская обл.). Здесь обнаружены плечевая кость (экз. ПИН, № 272/10) и клык (экз. ПИН, № 272/5), который мог принадлежать как раннему тероцефалу, так и горгонопии. Не исключено, что эти остатки принадлежат поростеогнату, поскольку фауна Малой Кинели соответствует ишеевскому комплексу (Вьюшков, 1955). Второй несомненный ранний тероцефал с территории Восточной Европы – Gorynychus masyutinae Kammerer et Masyutin – был описан по материалам из местонахождения Котельнич (Кировская обл.) (Kammerer, Masyutin, 2018).
Обнаруженные в местонахождении Сундырь-1 ранние тероцефалы представлены не менее чем двумя формами. Ниже приведено описание одной из них. Эта форма представлена в коллекции большим количеством ископаемых остатков. Она отличается более крупными размерами и удлиненной предглазничной частью черепа, что позволяет отнести ее к семейству Scylacosauridae.
МАТЕРИАЛ
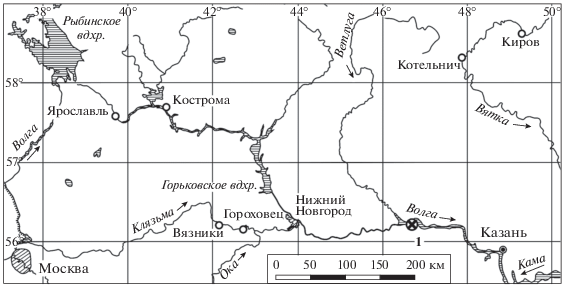
Материал происходит из местонахождения Сундырь-1, расположенного на правом берегу р. Волга (южный берег Чебоксарского водохранилища) непосредственно ниже устья р. Сундырь, в 2.5 км северо-западнее д. Токари и в 1.6 км восток-северо-восточнее с. Юльялы, в Горномарийском р-не Республики Марий Эл (рис. 1). Описание разреза и местонахождения приведено в работах В.К. Голубева (Голубев и др., 2015; Голубев, Буланов, 2018). Стратиграфическое положение костеносных слоев – средняя пермь, нижняя часть верхнесеверодвинского подъяруса (=кептенский ярус), нижний подгоризонт путятинского горизонта. Изученная коллекция ископаемых остатков хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (Москва), № 5388.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД THEROCEPHALIA
СЕМЕЙСТВО SCYLACOSAURIDAE BROOM, 1903
Род Julognathus Sutschkova et Golubev, gen. nov.
Название рода от Юл – марийское название р. Волга и gnathos греч. – челюсть.
Типовой вид – Julognathus crudelis sp. nov.
Диагноз. Крупный тероцефал, зубная формула I 5/3 C 1/1 Pc 6-7/7-9.
Видовой состав. Типовой вид.
Сравнение. От Porosteognathus отличается меньшим количеством зубов на praemaxillare (5) и maxillare (6–7) и более крупными размерами. От Alopecodon, Glanosuchus, Ictidosaurus, Pardosuchus, Pristerognathus и Scylacosaurus отличается меньшим количеством верхних резцов. От Alopecodon, Ictidosaurus, Pardosuchus и Scylacosaurus отличается отсутствием предклыковых зубов на maxillare.
Julognathus crudelis Sutschkova et Golubev, sp. nov.
Табл. X, XI (см. вклейку)
Burnetioidea fam. indet.: Голубев и др., 2015, с. 50; Сучкова, 2017г, с. 211; 2018, с. 231.
Gorgonopia fam. nov.: Сучкова, 2017а, с. 32; 2017б, с. 67; Suchkova, 2017, с. 193.
Название вида crudelis лат. – жестокий, безжалостный.
Голотип – ПИН, № 5388/51, левое dentale; Республика Марий Эл, Горномарийский р-н, местонахождение Сундырь-1; средняя пермь, верхнесеверодвинский подъярус, нижнепутятинский подгоризонт.
Описание (рис. 2, 3). Крупный тероцефал с длиной черепа до 43 см.
Рис. 2.
Julognathus crudelis gen. et sp. nov., череп сбоку. Реконструкция по голотипу ПИН, № 5388/51 и экз. ПИН, №№ 5388/58, 63, 90, 126, 316, 339, 421, 422. Длина масштабной линейки 2 см.
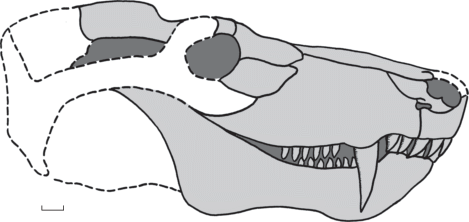
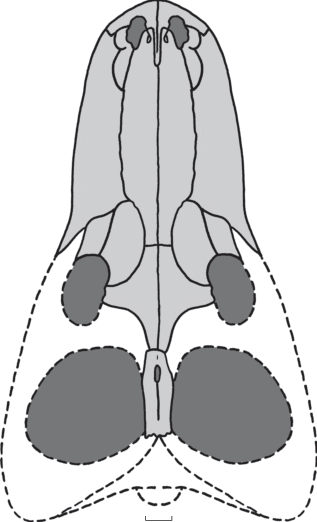
Рис. 3.
Julognathus crudelis gen. et sp. nov., череп сверху. Реконструкция по экз. ПИН, №№ 5388/58, 63, 90, 316, 339, 421, 422. Длина масштабной линейки 2 см.
Зубная кость достаточно массивная, наименьшую высоту имеет сразу позади зубного ряда, перед началом подъема к венечному отростку. Вентральный край кости после симфиза формирует площадку подбородочного выступа, слабо расширенную и покрытую мелко-пористой скульптурой. Подбородочный выступ по длине кости достигает уровня пятого-шестого заклыковых зубов. Он хорошо выражен на всех экземплярах, кроме самого маленького (экз. № 5388/124). За подбородочным выступом нижний край кости несколько вогнут вверх. Задне-нижний край dentale углообразно изогнут. В вершине угла этого изгиба развита латерально расширенная пяткообразная площадка, покрытая четкими продольными желобками. Площадка ориентирована под углом около 20° к продольной оси кости, по строению очень похожа на площадку подбородочного выступа. Резцово-клыковая область зубной кости приподнята над рядом заклыковых зубов. За ней наискосок к подбородочному выступу по лабиальной поверхности кости следует заметное вдавление – место прилегания верхнечелюстного клыка. Симфиз вытянутый, имеет субтреугольную форму, образован слабо выраженными контактными гребешками. Заклыковый зубной ряд отделен от лингвальной стороны кости продольной бороздой. По лингвальной стороне вплоть до симфизной поверхности (symphysis intermandibularis) проходит желоб sulcus mylohyoideus. Под началом подъема к венечному отростку он отдает ответвление в толщу кости. Венечный отросток крупный, поднимается под углом 30° в нижней части и 35° в верхней (экз. №№ 5388/126, 328). Венечный отросток на конце широкий, что типично для тероцефалов, его верхний край скруглен, а нижний вытянут и образует небольшой угол. Латеральная поверхность кости несет скульптуру в районе резцово-клыковой области. Она представлена мелкими отверстиями с отходящими от них короткими желобками. Далее кость практически гладкая, с редкими отверстиями для сосудов. Сзади под венечным отростком поверхность слабо прогибается, образуя небольшие валики по вентральному и дорсальному краям.
Зубной ряд включает три резца, клык и семь-девять заклыковых зубов. Почти на всех экземплярах зубы отсутствуют: выпали в результате посмертной мацерации или обломаны. Первые две резцовые альвеолы ориентированы несколько наклонно вперед и расположены очень тесно, на экземплярах с повреждением этой области стенка между ними часто разрушается. Длина резцовой области приблизительно равна диаметру клыковой альвеолы. Клыковая альвеола крупная, овальная. На экз. № 5388/125 впереди от обломанного корня действующего клыка видны остатки маленького сменного зуба. На экз. № 5388/310 клык отсутствует, альвеола неравномерно заросла костной тканью.
Заклыковые зубы тесно расположены. У самых мелких экземпляров (№№ 5388/124, 51, 310) заклыковый зубной ряд отделяется от клыка диастемой. У всех остальных, более крупных экземпляров диастема отсутствует, и начало ряда заходит на подъем к резцово-клыковой области. Размеры заклыковых зубов часто неодинаковы: на экз. № 5388/51 и № 5388/125 альвеола первого зуба ряда заметно меньше остальных в диаметре, на экз. № 5388/124 и № 5388/126 уменьшены первый и последний зубы в ряду (Сучкова, 2017б). Судя по сохранившимся заклыковым зубам на голотипе, они имели овальную шейку и несколько уплощенную с боков, зауженную к вершине коронку с мелко зазубренными передним и задним режущими кантами.
Пластинчатая кость (spleniale) прямоугольной формы, высокая, плоская, без медиального выступа в симфизной части.
Предчелюстная кость (praemaxillare) массивная, вытянутая. Вертикальные отростки (spina nasalis) тонкие и имеют узко-треугольную форму в сечении. Симфиз покрыт неглубокими гребешками в передней части и более резкими – в задней. Проксимальный край кости утолщен в области fossa dentia canini inferioris. Область причленения премаксиллярного отростка челюстной кости (processus praemaxillaris ossis maxillaris) неровная, покрыта желобами и гребнями; с малозаметным перегибом переходит в дорсальную часть контакта с maxillare. На дорсальной поверхности кости в основании spina nasalis присутствует сосудистое отверстие. Оно, очевидно, связано каналом (canalis apicalis) с передним премаксиллярным отверстием под основанием spina nasalis на латеральной стороне. Далее назад резко выделяется площадка налегания septomaxillare, расположенная несколько наклонно наружу и отделенная с внутренней стороны от вогнутой околосимфизной области высоким гребнем, соответствующим juga alveolaris incisivi. Площадка очень крупная, скульптированная хорошо выраженными контактными гребешками. Зона контакта с сошником хорошо видна на вентральной стороне кости, поверхность небной пластины кнаружи от нее вогнута, образуя подобие очень небольшого сошникового отростка. Вхождение vomer продолжается длинной контактной площадкой вперед вдоль симфиза почти до середины длины кости. Небная пластина короткая и широкая, слабо вырезанная хоаной.
Вентральная поверхность предчелюстной кости несет пять очень крупных альвеол резцов. Первая альвеола самая маленькая в диаметре, остальные крупнее, самая большая в ряду – третья. В глубине некоторых альвеол наблюдается двураздельность, отражающая, по-видимому, смену зубов (третья на экз. № 5388/58 и 4-я и 5-я на экз. № 5388/355). Отчетливых сосудистых отверстий там не просматривается. У экз. № 5388/57 рядом со стенкой третьей видной на образце альвеолы из толщи кости выдается кончик не полностью выросшего резца. Он несет мелкую зазубренность на внешнем, открытом для изучения канте.
Верхнечелюстная кость (maxillare) длинная и относительно низкая, достигает максимальной высоты в области сумки клыка, которая несколько отклоняется назад. Отростки (processus praemaxillaris и pr. jugalis) вытянутые. Тело кости уплощенное, за исключением альвеолярного заклыкового ряда и очень крупной, вздутой сумки клыка. На лицевой пластине развита скульптура, состоящая из мелких ямок с отходящими от них желобками и бороздок. Ямки концентрируются в передней части лицевой пластины над клыком, становясь более редкими по краям. В задней и задне-верхней части кости скульптура состоит в основном из узких частых бороздок, направленных преимущественно назад над processus jugalis и назад и вверх к контакту с praefrontale. Бороздки начинаются на уровне второго заклыкового зуба. Вентральный край кости неровный: на уровне внешней стенки сумки клыка он несколько выдается вниз относительно заклыкового зубного ряда, а на уровне pr. praemaxillaris – вогнут в проксимальной части и поднят над линией заклыкового зубного ряда. Особенно это заметно на экз. №№ 5388/346 и 434. Начальная его часть с внутренней стороны гладкая, образует часть ямы для нижнечелюстного клыка (fossa dentia canini inferioris), далее дистально расположены контактные гребешки для соединения с praemaxillare. Контактная поверхность ограничена сверху отгибающимся медиально-вверх гребнем, который ложится на дорсальную часть praemaxillare. Между этим гребнем, внутренней поверхностью кости и высокой передней стенкой сумки клыка образуется полость. Передний край maxillare непосредственно выше уровня контактной поверхности с предчелюстной костью образует вырезку, хорошо заметную даже на сильно поврежденных фрагментах (экз. №№ 5388/346, 138, 434, 137). Вырезка формирует со стороны maxillare септомаксиллярное отверстие (foramen septomaxillaris). Над ней по латеральной поверхности кости при хорошей сохранности видна площадка налегания лицевого отростка septomaxillare (экз. №№ 5388/138, 346, 354, 434).
Сумка клыка крупная, вздутая и широкая даже на самом маленьком экз. № 5388/427, при хорошей сохранности в передне-заднем направлении всегда двураздельная и вмещающая остатки сменного клыка. Основной клык всегда расположен впереди от сменного. Сумка клыка особенно массивна в нижней половине (экз. №№ 5388/316, 434) или трети (экз. №№ 5388/346, 354), где разрастается и формирует широкий гребень crista choanalis, вдающийся в хоану. На вентральной стороне он отделен от собственно альвеолы глубокой бороздой, доходящей до контакта с palatinum. Контакт этот также заходит на crista choanalis. Таким образом, по ширине crista choanalis перекрывает не только клык, но и один-два заклыковых зуба. На самом маленьком образце гребень небольшой, занимает приблизительно четверть высоты сумки, но все же хорошо заметен. Медиальная поверхность сумки клыка часто неравномерно выпуклая, с более выпуклой передней половиной, за которой располагался корень функционирующего клыка, и несколько уплощенной задней, за которой располагался растущий сменный клык.
Позади клыка на вентральной поверхности maxillare располагается ряд из шести–семи заклыковых зубов или альвеол. На экз. № 5388/316 первая, третья и последняя (шестая?) альвеолы пусты, во второй и предпоследней располагаются корни обломанных зубов. Закономерность в чередовании пустых и несущих зубы альвеол может объясняться примитивной альтернативной сменой зубов. На экз. № 5388/140 все зубы остались в альвеолах, обломаны, четвертая с конца (третья с начала?) альвеола, от которой сохранилась только медиальная стенка, выглядит пустой. На экз. № 5388/346 сохранились первые три альвеолы – в первой и второй зубы обломаны, третья пуста. На экз. № 5388/272 сохранилась только средняя часть зубной пластины, на которой пустые альвеолы чередуются с сохранившимися зубами (две альвеолы и два зуба). Эти зубы конические, немного уплощенные с боков к вершинкам, которые слегка загнуты назад; оба канта – передний и задний – несут мелкую зазубренность, очень похожи на заклыковые зубы на зубных костях экз. №№ 5388/51 и 5388/126.
На заднем и задне-нижнем крае сумки клыка и частично под crista choanalis на альвеолярном крае кости виден контакт с palatinum в виде гребней и желобков. Дорсальная поверхность альвеолярного края образует дно максиллярного синуса (sinus Highmori). Он узкий на всем своем протяжении, особенно в задней части. Медиальная стенка, насколько можно судить, у него не развита. Альвеолярный край кости понижается в заднем направлении, переходя в длинный processus jugalis. Его задний конец располагается в 5–6 см позади последнего заклыкового зуба (экз. №№ 5388/140, 316). С дорсальной стороны pr. jugalis имеет вид открытого желоба. Медиально он ограничен на высоту 3–5 мм продолжением альвеолярного края. Приблизительно на середине его вентральная поверхность изгибается вверх гребнем, резко сужая желоб pr. jugalis. С внешней стороны кости вентральная часть отростка отделена от латеральной глубоким желобом, который проходит назад и переходит на вентральную, а затем на медиальную стороны кости. Хорошо заметен он на экз. № 5388/316, меньше – на экз. № 5388/140. На последнем в этой области видны пара небольших отверстий сосудов.
Внешняя поверхность maxillare более мелкого и лучше сохранившегося экз. № 5388/140 несет в передней части более заметные, чем в той же области экз. № 5388/316, бугорки и ямки. С увеличением размеров верхнечелюстной кости скульптура поверхности становится менее выраженной, за счет более редкого расположения ямок с желобками и бугорков. При этом скульптура заметнее выражена на средней части лицевой пластины, постепенно исчезая к краям кости.
Самая крупная верхнечелюстная кость (экз. № 5388/354) имеет в высоту 8 см, в сравнении с экз. № 5388/316 ее реконструируемая длина могла быть 20.9 см, что по пропорциям, приведенным, например, на изображении Glanosuchus macrops (van den Heever, 1994, с. 9) дает приблизительную длину черепа в 43 см.
Теменные кости (экз. № 5388/421) крупные (длина сохранившегося фрагмента – 7.8 см, ширина до 2.4 см, высота – 3 см), узкие, сросшиеся по сагиттальному шву. Пинеальное отверстие небольшое, вытянутое и узкое, расположено в передней части parietalia. С вентральной стороны оно окружено неглубоким вдавлением. Далее назад кость образует шовную поверхность без образования здесь отогнутых пластин angulus parietalis. Кости неполностью срастаются по сагиттальному шву, а сужение после пинеального отверстия выражено очень слабо, что отличает данный вид от ишеевского Porosteognathus efremovi (Ivakhnenko, 2011).
Помимо сохранившихся в зубных и челюстных костях заклыковых зубов, в коллекции присутствует множество изолированных резцов, клыков и заклыковых зубов. Клыки варьируют по длине коронки от 0.9 до 7.2 см, они слабоизогнуты на всем протяжении, каплевидные в сечении основания, несут передний и задний канты с мелкой зазубренностью на них. Передний режущий кант немного короче заднего, а в некоторых случаях сильно сокращен (экз. № 5388/141) или отсутствует (экз. №№ 5388/112, 374). Резцы также обнаруживают закономерность в длине ряда зазубрин на режущих кантах: с одной из сторон зазубренный кант более короткий или отсутствует. Изолированные заклыковые зубы достаточно легко определить, благодаря возможности сравнения с сохранившимися зубами в зубных костях экз. № 5388/51 и № 5388/126 и челюстной кости экз. № 5388/272. Они отличаются мелкой зазубренностью и высокими шейками (3–4 мм при высоте коронки около 1 см).
Материал. Кроме голотипа, из типового местонахождения зубные кости (экз. ПИН, №№ 5388/52, 53, 124–126, 310, 328, 381), пластинчатая кость (экз. ПИН, № 5388/127), предчелюстные кости (экз. ПИН, №№ 5388/57, 58, 105, 355), челюстные кости (экз. ПИН, №№ 5388/137, 138, 140, 167, 191, 272, 316, 346, 351, 354, 359), теменной гребень (экз. ПИН, №№ 5388/ 421), резцы (экз. ПИН, №№ 5388/116, 153, 264, 281, 367, 368), клыки (экз. ПИН, №№ 5388/108, 110–115, 141, 293, 294, 308, 362, 365, 369, 373, 374), заклыковые зубы (экз. ПИН, №№ 5388/174, 175, 177, 311, 376).
СИНЭКОЛОГИЯ
Описанный из местонахождения Сундырь-1 Juloghathus crudelis – самый крупный хищник в сундырском комплексе. В структуре сундырского сообщества тетрапод он занимал нишу доминирующего хищника и, вероятно, образовывал трофическую пару с растительноядным тапиноцефалом. Таким образом, сундырский комплекс – это третий комплекс тетрапод Восточной Европы, в котором хищные тероцефалы занимали ведущие позиции в доминантном блоке. Подобная ситуация наблюдается также в котельничском и вязниковском комплексах (Сенников, Голубев, 2017). Однако в котельничском сообществе роль доминирующего хищника тероцефалы делят вместе с горгонопиями, а в вязниковском – с архозаврами–протерозухидами. В сундырском доминантном сообществе, кроме тероцефалов, другие хищники не известны. Как было отмечено выше, присутствие здесь хищных диноцефалов – титанозухий и горгонопий не подтвердилось. Эта особенность сундырского комплекса отличает его от всех комплексов Восточной Европы и Южной Африки, в которых тероцефалы никогда не были единственными крупными хищниками.
* * *
Работа выполнена при финансовой поддержке РФФИ (проекты №№ 17-04-01937, 17-04-00410, 17-54-10013) и программы фундаментальных научных исследований Президиума РАН № 22 “Эволюция органического мира и планетарных процессов”, а также за счет средств субсидии, выделенной в рамках государственной поддержки Казанского (Приволжского) федерального университета в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров (The work is performed according to the Russian Government Program of Competitive Growth of Kazan Federal University).
Список литературы
Буланов В.В., Голубев В.К. Водный блок сундырского сообщества позднепермских позвоночных Восточной Европы // Пермская система: стратиграфия, палеонтология, палеогеография, геодинамика и минеральные ресурсы. Сб. матер. Междунар. науч. конф., посвящ. 170-летию со дня открытия пермской системы (5–9 сент. 2011 г.). Пермь: Пермский гос. ун-т, 2011. С. 47–55.
Вьюшков Б.П. Об относительном возрасте Ишеевской и Северодвинской фаун наземных позвоночных перми СССР // Докл. АН СССР. 1952. Т. 83. № 6. С. 897–900.
Вьюшков Б.П. О фауне верхнепермских наземных позвоночных с реки Малая Кинель // Тр. Палеонтол. ин-та АН СССР. 1955. Т. 49. С. 176–189.
Голубев В.К. Граница среднего и верхнего отделов пермской системы на Восточно-Европейской платформе // Палеонтология и стратиграфические границы. Матер. 58 сесс. Палеонтол. об-ва (С.-Петербург, 2–6 апреля 2012 г.). СПб.: Изд-во ВСЕГЕИ, 2012. С. 43–45.
Голубев В.К. Диноцефаловый этап истории развития пермской фауны тетрапод Восточной Европы // Академик Юрий Александрович Орлов. К 120-летию со дня рождения. Сб. статей. М.: ПИН РАН, 2018. С. 111–120.
Голубев В.К., Буланов В.В. Амфибии сундырского фаунистического комплекса пермских тетрапод Восточной Европы // Палеонтол. журн. 2018. № 6. С. 50–62.
Голубев В.К., Куркин А.А., Сенников А.Г. О возрасте сундырского фаунистического комплекса пермских тетрапод Восточно-Европейской платформы // Учен. зап. Казанск. ун-та. Сер. Естеств. науки. 2015. Т. 157. Кн. 1. С. 49–58.
Голубев В.К., Куркин А.А., Сенников А.Г., Березин А.Ю. Новая фауна пермских тетрапод Восточной Европы // Палеострат-2011. Годичн. собр. секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН (Москва, 24–26 января 2011 г.). Прогр. и тез. докл. М.: ПИН РАН, 2011. С. 30–32.
Голубев В.К., Сенников А.Г. Среднепермское событие в истории фауны тетрапод Восточной Европы // Позвоночные палеозоя и мезозоя Евразии: эволюция, смена сообществ, тафономия и палеогеография. Матер. конф., посвящ. 80-летию со дня рожд. Виталия Георгиевича Очева (1931–2004) (6 декабря 2011 г., ПИН РАН, Москва). М.: ПИН РАН, 2011. С. 13–16.
Голубев В.К., Сенников А.Г. Северодвинский кризис сообщества тетрапод Восточной Европы // Диверсификация и этапность эволюции органического мира в свете палеонтологической летописи. Матер. 60 сесс. Палеонтол. об-ва, посвящ. 100-летию со дня рождения акад. Б.С. Соколова (Санкт-Петербург, 7–11 апреля 2014 г.). СПб.: ВСЕГЕИ, 2014. С. 161–163.
Ефремов И.А. Фауна наземных позвоночных в пермских медистых песчаниках Западного Приуралья // Тр. Палеонтол. ин-та АН СССР. 1954. Т. 54. С. 1–416.
Куркин А.А. Новый галеопид (Anomodontia, Galeopidae) из перми Восточной Европы // Палеонтол. журн. 2017. № 3. С. 81–85.
Сенников А.Г. Ранние текодонты Восточной Европы. М.: Наука, 1995. 141 с. (Тр. Палеонтол. ин-та РАН. Т. 263).
Сенников А.Г. Некоторые проблемы типологии сообществ наземных тетрапод // Эволюция биосферы и биоразнообразия. К 70-летию А.Ю. Розанова. М.: КМК, 2006. С. 359–372.
Сенников А.Г., Голубев В.К. Последовательность пермских фаун тетрапод Восточной Европы и пермо-триасовый экологический кризис // Палеонтол. журн. 2017. № 6. С. 30–41.
Сучкова Ю.А. Горгонопии в сундырской фауне (средняя пермь, Восточная Европа) // Современная палеонтология: классические и новейшие методы. 14 всеросс. научн. школа молодых ученых-палеонтологов, 2–4 октября 2017 г. М.: ПИН РАН, 2017a. С. 32.
Сучкова Ю.А. Особенности нижнего заклыкового зубного ряда у сундырских горгонопий // Морфогенез в индивидуальном и историческом развитии: онтогенез и формирование биологического разнообразия (Москва, 22–24 ноября 2017 г.). Тез. конф. М.: ПИН РАН, 2017б. С. 66–67.
Сучкова Ю.А. Развитие подбородочного выступа на зубных костях в онтогенезе горгонопий сундырского комплекса (поздняя пермь, Восточная Европа) // Палеострат-2017. Годичн. собр. (научн. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН (Москва, 30 января – 1 февраля 2017 г.). Тез. докл. М.: ПИН РАН, 2017в. С. 66–67.
Сучкова Ю.А. Первая находка погрызов на костях пермских тетрапод Восточной Европы // Интегративная палеонтология: перспективы развития для геологических целей. Матер. 63 сесс. Палеонтол. об-ва (С.‑Петербург, 3–7 апреля 2017 г.). СПб., 2017г. С. 210–211.
Сучкова Ю.А. Новые данные о составе фауны пермских терапсид из местонахождения Сундырь-1 (Марий Эл) // Фундаментальная и прикладная палеонтология. Матер. 64 сессии Палеонтол. об-ва при РАН (2‒6 апреля 2018 г., С.-Петербург). СПб.: ВСЕГЕИ, 2018. С. 231.
Татаринов Л.П. Инфракласс Therosauria // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1. М.: ГЕОС, 2008. С. 184–223.
Ульяхин А.В. Архаичный двинозаврид из позднепермского местонахождения Сундырь-1, Марий Эл // Палеострат-2018. Годичн. собр. (научн. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН (Москва, 29–31 января 2018 г.). Прогр. и тез. докл. М.: ПИН РАН, 2018. С. 56–57.
Abdala F., Kammerer C.F., Day M.O. et al. Adult morphology of the therocephalian Simorhinella baini from the middle Permian of South Africa and the taxonomy, paleobiogeography, and temporal distribution of the Lycosuchidae // J. Paleontol. 2014. V. 88. P. 1139–1153.
Abdala F., Rubidge B.S., van den Heever J. The oldest therocephalians (Therapsida, Eutheriodontia) and the early diversification of Therapsida // Palaeontology. 2008. V. 51. P. 1011–1024.
Arefiev M.P., Golubev V.K., Kuleshov V.N., Pokrovsky B.G. Paleogeographical reorganization of the Moscow Syneclise during the Severodvinian (Capitanian-Wuchiapingian) time based on isotopic (δ13C and δ 18O) and paleontological data, Permian Period, East European Platform // Paleontol. J. 2015. V. 49. № 11. P. 1206–1227.
Bulanov V.V., Golubev V.K. New data on enosuchid anthracosauromorphs (Amphibia) of the Middle–Late Permian of European Russia: Part 2. New taxa of Enosuchidae // Paleontol. J. 2018. V. 52. № 13. P. 1623–1632.
Golubev V.K. Dinocephalian stage in the history of the Permian tetrapod fauna of Eastern Europe // Paleontol. J. 2015. V. 49. № 12. P. 1346–1352.
Ivakhnenko M.F. Permian and Triassic therocephals (Eutherapsida) of Eastern Europe // Paleontol. J. 2011. V. 45. № 9. P. 981–1144.
Kammerer C.F., Masyutin V. A new therocephalian (Gorynychus masyutinae gen. et sp. nov.) from the Permian Kotelnich locality, Kirov Region, Russia // PeerJ. 2018. 6:e4933; https://doi.org/10.7717/peerj.4933
Kurkin A. New locality with dinocephalian fauna in the European Russia // Proc. of the 16th Conf. of the Palaeontological Society of Southern Africa (Howick, August 5–8, 2010) / Eds. Mostovski M.B., Ovechkina M.N. Pietermaritzburg, 2010. P. 55–57.
Olson E.C. Community evolution and the origin of mammals // Ecology. 1966. V. 47. № 2. P. 291–302.
Suchkova Yu. A. Gorgonopians in Permian tetrapod communities of Eastern Europe // Upper Palaeozoic Earth systems: high-precision biostratigraphy, geochronology and petroleum resources. Kazan Golovkinsky Stratigraphic Meeting – 2017 and Fourth All-Russ. Conf. “Upper Palaeozoic of Russia”. Kazan, 19–23 September, 2017. Abstract vol. Kazan: Kazan Univ. Press, 2017. P. 193–194.
Van den Heever J.A. The comparative and functional cranial morphology of the early Therocephalia (Amniota: Therapsida). PhD thesis. Univ. Stellenbosch, 1987. 576 p.
Van den Heever J.A. The cranial anatomy of the early Therocephalia (Amniota: Therapsida) // Ann. Univ. Stellenbosch. 1994. V. 1. P. 1–59.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал