

Палеонтологический журнал, 2019, № 4, стр. 93-106
Новые материалы по представителям мелких гиенодонов (Hyaenodontinae, Creodonta) из палеогена Монголии
А. В. Лавров *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: lavrov@paleo.ru
Поступила в редакцию 31.03.2017
После доработки 30.09.2017
Принята к публикации 14.02.2019
Аннотация
По горизонтальной ветви нижней челюсти описан новый вид Hyaenodontinae, самый мелкий азиатский вид – Hyaenodon pumilus sp. nov. из верхнего эоцена свиты Эргилин-Дзо Юго-Восточной Монголии. Описаны новые экземпляры H. chunkhtensis Dashzeveg, 1985 из местонахождений Хоер-Дзан (свита Эргилин-Дзо, верхний эоцен) и Татал-гол (свита Шанд-Гол, верхний олигоцен). Экземпляры H. chunkhtensis могут быть разделены на два морфотипа.
Гиенодонтины (Hyaenodontinae) были широко распространены в позднем эоцене – олигоцене Северной Америки, Европы и Азии, а в Китае дожили до раннего миоцена (Wang et al., 2005). Гиенодонтины разнообразны по размерам тела и габитусу лицевого отдела. По размерам они охватывают диапазон от куницы (“Hyaenodon” microdon) до медведя (“Hyaenodon” gigas). Особенности строения внутреннего уха Hyaenodontinae показывают, что они заслуживают выделения в отдельное подсемейство (Polly, 1996). Согласно Д. Поли, Hyaenodontinae включают роды Protohyaenodon Stock, 1933 и Neohyaenodon Thorpe, 1922, а также типовой род Hyaenodon Leidy, 1869 [типовой вид H. leptorhynchus (Laizer et Parieu, 1838)], объединяющий европейские виды. Вероятное обособленное систематическое положение европейских видов Hyaenodontinae подтверждается тем, что они имеют иной порядок прорезания постоянных зубов (раннее прорезание С1 и позднее прорезание Р3; Balst et al., 2011). Среди представителей Hyaenodontinae выделяется группа карликовых видов, характеризующихся мелкими размерами: от современной куницы (Martes martes L., 1758) до серой лисицы (Urocyon cinereoargenteus Schreber, 1775). Реконструируемая кондилобазальная длина черепа мелких гиенодов находится в диапазоне 80–130 мм. Из Северной Америки описано три вида мелких гиенодонов (Protohyaenodon venturae Mellett, 1977, P. microdon Mellett, 1977 и Hyaenodon raineyi Gustafson, 1986). Один вид известен из Европы (H. filholi Schlosser, 1977). В Азии до настоящего времени было известно два вида мелких гиенодонов (H. eminus Matthew et Granger, 1925 и H. chunkhtensis Dashzeveg, 1985). Всего насчитывается шесть видов мелких гиенодонов. При крайней фрагментарности находок морфологическое разнообразие карликовых видов высоко. Поэтому в настоящее время они не могут быть объединены в один род.
Карликовые формы традиционно относились к роду Hyaenodon. Например, Д. Дашзевэг (Dashzeveg, 1985) описал азиатского карликового гиенодона как H. chunkhtensis, Э. Густафсон (Gustafson, 1986) описал новый вид американского карликового гиенодона как H. raineyi. В диагноз Neohyaenodon и Protohyaenodon было включено отношение длины М2 к длине верхнего щечного зубного ряда Р1–М2. Для Neohyaenodon отношение LM2/LP1–M2 было установлено равным около 23%, для Protohyaenodon – около 19% (Mellett, 1977). Для Hyaenodon raineyi отношение LM2/LP1–M2 составляет 17.4–19.4% (у P. venturae – 18.4%). Д. Мелле исследовал образцы H. raineyi и отнес его к Protohyaenodon, но не включил этот вид в свою монографию (Mellett, 1977). Для азиатского H. chunkhtensis значение отношения LM2/LP1–M2 неизвестно, и мы относим его к типовому роду, согласно первоописанию (Dashzeveg, 1985). Единственный европейский представитель карликовых гиенодонов H. filholi отличается от остальных карликовых форм двухкорневым Р1 (это коррелирует с его удлиненным лицевым отделом) и иным порядком прорезания постоянных зубов (Balst et al., 2011).
Первый карликовый гиенодон Азии H. eminus был описан по двум фрагментам нижней челюсти и изолированному М2 (Matthew, Granger, 1925). В отличие от остальных карликовых форм, у этого вида Р1 не только однокорневой, но и сильно редуцирован. По размерам H. eminus (LM1–M3 – 24.2 мм; LP1–M3 – 58.0 мм) почти на треть крупнее, чем H. chunkhtensis. Позже была описана полная горизонтальная ветвь нижней челюсти H. eminus c сохранившейся альвеолой клыка и полным М3 (Dashzeveg, 1964). Было установлено, что между С1 и Р1 имеется большая диастема. Все находки вида происходят из отложений верхнего эоцена (Berkey, Grander, 1923; Dashzeveg, 1964, 1985). H. chunkhtensis Dashzeveg, 1985 был описан из олигоцена Монголии. Его челюсть почти вдвое массивнее (по отношению высоты горизонтальной ветви к длине щечного зубного ряда P1–M3), чем у H. eminus, а горизонтальная ветвь не имеет выраженного сужения по направлению к С1.
Hyaenodon eminus и Protohyaenodon venturae – самые крупные представители карликовых гиенодонтин. При этом H. eminus имеет грацильную низкую нижнюю челюсть, а нижняя челюсть P. venturae высокая и очень массивная. Реконструируемая нами кондилобазальная длина черепа (L1) у H. eminus – около 130 мм, у P. venturae – 110–120 мм. Реконструируемая L1 для Hyaenodon raineyi – около 105–110 мм. Эти размеры соответствуют длине черепа современной серой лисицы (Urocyon cinereoargenteus, Canidae): L1 = 110–130 мм (Fritzell, Haroldson, 1982). Самые мелкие гиенодоны Protohyaenodon microdon и Hyaenodon pumilus sp. nov. имели размеры нижней челюсти, равные челюсти современной Martes martes (Mustelidae, Carnivora): у них L1 была около 70–80 мм. Для сравнения, у Protohyaenodon mustelinus Scott, 1895 – самого мелкого американского гиенодона, не относящегося к карликовым формам – L1 более 157 мм.
Экземпляры, описываемые в этой статье, были собраны в ходе работ ССМПЭ в 1980-е гг. в южной части Монголии (рис. 1). Образцы из местонахождения Хоер-Дзан собраны под руководством В.Ю. Решетова. Экз. ПИН, № 3110/1217 (рис. 2) был обнаружен на местонахождении Хоер-Дзан (рис. 1) в нижней части разреза (белая толща), соответствующей по возрасту верхнему эоцену. Экз. ПИН, № 3110/618 найден автором в 1989 г. в районе северного обо́ столовой горы Хоер-Дзан в белой толще. На несколько лет раньше там же был обнаружен фрагмент нижней челюсти с Р3–Р4 Hyaenodon chunkhtensis (экз. ПИН, № 3110/552). Фрагмент левой ветви нижней челюсти с Р4–М2 и альвеолой Р3 (экз. ПИН, № 3935/538) был найден на местонахождении Татал-Гол непосредственно над вторым слоем базальтов в 1972 г. (сборы Е.Н. Курочкина, ПИН РАН). Эта находка относится к отложениям свиты Шанд-Гол, биозона С, возраст 26–27 млн лет, поздний олигоцен (Daxner-Höck et al., 2010). В данной работе приводятся описания всех этих образцов, так как материал по карликовым гиенодонам Азии исключительно редок.
Рис. 1.
Схема местонахождений находок азиатских видов мелких гиенодонов: 1 – Хоер-Дзан (верхний эоцен); 2 – Их-Дзан (верхний эоцен); 3 – Татал-Гол (олигоцен); 4 – Шункт (нижний олигоцен).
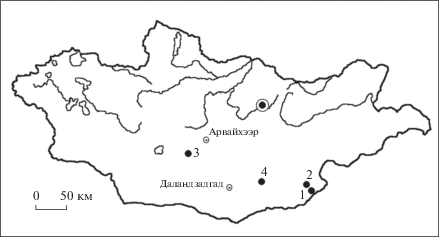
Рис. 2.
Hyaenodon pumilus sp. nov., голотип ПИН, № 3110/1217, правая ветвь нижней челюсти с С1–М3; Монголия, местонахождение Хоер-Дзан; верхний эоцен: а – с латеральной стороны; б – с медиальной стороны; в – сверху.
МАТЕРИАЛ И МЕТОДЫ
Использованный материал: Hyaenodon pumilus sp. nov.: голотип ПИН, № 3110/1217; H. chunkhtensis: экз. ПИН, №№ 3110/552; 3110/618; 3935/552; экз. PST, № 21-1; H. filholi: экз. ГГМ, №№ X-520/17; X-520/7; X-520/8; X-520/11; № X-520/14; X‑520/15; X-520/18; X-520/19; H. venturae: экз. USNM, № 214643; экз. СIT-LACM, № 1140 (слепок); экз. USNM, № 214643; H. microdon: экз. AMNH, № 9623; экз. USNM, № 187569; Neohyaenodon vetus: экз. LACM, № 1243; Protohyaenodon crucians: экз. AMNH, № 647; экз. FAM, № 75652 (слепок); экз. FAM, № 75565 (слепок); Hyaenodon cf. chunkhtensis: экз. SHG, № S/29 (слепок).
Cокращения: ПИН – Палеонтологический ин-т им. А.А. Борисяка РАН (Москва, Россия); ГГМ – Государственный геологический музей им. В.И. Вернадского (Москва, Россия); FAM – коллекция Фрика, Американский музей естественной истории (Нью-Йорк, США); USNM – Национальный музей естественной истории, отдел палеобиологии (Вашингтон, США); CIT-LACM – Палеонтологическая коллекция позвоночных Калифорнийского технологического ин-та (Лос-Анджелес, США); PST – коллекция Отдела палеонтологии и стратиграфии Геологического ин-та Академии наук Монголии (Улан-Батор, Монголия); MPC-M – коллекция ископаемых млекопитающих Монгольского палеонтологического центра (Улан-Батор, Монголия); SHG – коллекция местонахождения Шанд-Гол Центральной Монголии, Музей естественной истории, Вена, Австрия.
Обозначения измерений: L – длина; W – ширина; Ltal – длина талонида; h – высота; hm1 – высота М1; Lm1 – длина М1; lpa – длина параконида; lpr – длина протоконида; L1 – кондилобазальная длина черепа; m. – musculus; for. – foramen. Все измерения приведены в миллиметрах (мм), высота зубов измерялась с вентрального края от границы эмали до вершины самого высокого конуса. Высота М3 измерялась по паракониду и по протокониду. KhD – коэффициент массивности нижней челюсти, где h –высота; D – dentale. KhM1 – высота нижней челюсти перед М1 (между Р4 и М1). В работе использованы результаты вычисления индексов моляров М1 и М2, выполненные Б. Ланж-Бадре (Lange-Badre, 1979).
Автор выражает свою глубокую благодарность А.К. Агаджаняну (ПИН РАН) за чтение рукописи и внесенные замечания; Р. Эмри (USNM, Paleobilogy Department, США) за бесценную помощь в работе; Д. Нагель (Ин-т палеонтологии, Университет Вены, Австрия) за присланные слепки образцов; М.Н. Кандинову и И.А. Стародубцевой (отдел фондов ГГМ им. В.И. Вернадского) за предоставленный материал из фондов музея и консультации. Работа выполнена при поддержке грантов РФФИ–08-04-00483, 09-04-01303, 11-04-00933А, 11–06–12030-ОФИ-М-2011, и ОФИ 13-06-5-ОФИ-М-2013, а также программы I.21П “Биоразнообразие природных систем”, программы фундаментальных исследований Отделения биологических наук РАН “Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга”.
ОТРЯД CREODONTA
СЕМЕЙСТВО HYAENODONTIDAE LEIDY, 1869
ПОДСЕМЕЙСТВО HYAENODONTINAE LEIDY, 1869
Триба Hyaenodontini Leidy, 1869
Род Hyaenodon Leidy, 1869
Hyaenodon pumilus Lavrov, sp. nov.
Табл. X , фиг. 1 (см. вклейку)
Название вида pumilus лат. – карликовый.
Голотип – ПИН, № 3110/1217, правая горизонтальная ветвь нижней челюсти (венечный, угловой и сочленовный отростки отбиты), содержащая C1, P2, P4–M3, альвеолы I2-I3, P1 и P3; Монголия, Восточно-Гобийский (Дорногоби) аймак, местонахождение Хоер-Дзан (рис. 1); верхний эоцен, свита Эргилин-Дзо.
Описание (рис. 2). Горизонтальная ветвь нижней челюсти принадлежит взрослому, но еще молодому животному: М3 полностью прорезался, но все зубы слабо стерты, включая и М1 (заметны следы умеренного износа). Нижняя челюсть грацильная, сжата латеромедиально, без выраженных костных валиков вдоль ее длинной оси, дорсальный и вентральный края горизонтальной ветви почти прямые (рис. 2). Горизонтальная ветвь почти прямая, она ниже, чем у H. filholi. Размеры горизонтальной ветви нижней челюсти описываемого образца близки к таковым у современной Martes martes (Mustelidae, Carnivora). Масcетерный гребень хорошо выражен, в сохранившейся вентральной части венечного отростка узкий, тонкий и низкий, подобно этому гребню у P. venturae. Массетерная ямка глубокая, имеет типичную для гиенодонов треугольную форму. Симфиз субгоризонтальный, синдосмозный, подобно другим мелким видам гиенодонтин (за исключением P. microdon). Симфиз длиннее, чем у H. filholi и достигает середины P3 (у H. filholi задний край симфиза достигает только переднего края P3). Подбородочных отверстий два: переднее – округлой формы и расположено под альвеолой Р1, второе отверстие овальной формы, расположено под задним корнем Р3. Парастилиды моляров развернуты в лингвальном направлении. Отклонение направления парастилидов от сагиттальной оси уменьшается от M1 к M3. На медиальной поверхности горизонтальной ветви под протоконидом M3 отмечается шероховатая поверхность места прикрепления дистального апоневроза внутренней порции m. temporalis. Площадка прикрепления имеет форму равнобедренного треугольника, направленного вершиной вниз, шириной 2 мм и высотой около 4 мм.
C1 массивный, он слабо наклонен вперед по сравнению с клыком H. filholi, по форме и степени наклона в челюсти похож на C1 y H. chunkhtensis. Ряд премоляров компактный, без диастем. P1 однокорневой (подобно Р1 большинства других мелких видов), судя по альвеоле, он ассиметричный и был сильно наклонен вперед; он плотно прилегает кС1 и Р2. P2 ассиметричный, его талонид короче, чем у H. chunkhtensis; коронка зуба заметно расширяется каудолингвально, лингвальный цингулид хорошо развит; P3, судя по альвеоле, лишь немного длиннее P2, подобно H. chunkhtensis. P4 сильно сжат лабиолингвально, его коронка слабо расширяется в заднем направлении, наибольшая ширина зуба в задней части талонида; парастилид имеет вид приостренного конуса высотой около 0.5 мм, главный конус слабо наклонен назад и имеет хорошо выраженные режущие лезвия по своему переднему и заднему краю. У M1 протоконид выше и длиннее параконида: они разделены глубокой хищнической вырезкой; талонид короткий, режущий. Фасетка стирания от окклюзии с протоконом М1 хорошо выражена. В отличие от H. chunkhtensis и H. filholi, тригонид М1 низкий по отношению высоты зуба к его длине. Из-за этого М1 по форме имеет определенное сходство с DP4 других гиенодонов.
Талонид М1 крупный, в отличие от других Hyaenodontinae. Он имеет подобие рудиментарного бассейна, ограниченного посткристидом. Длина талонида равна 1.0 мм. Его длина по отношению к длине зуба визуально кажется существенно больше по сравнению с М1 H. chunkhtensis. При том, что длина М1 у последнего вида крупнее (табл. 1 ), длина талонида у него меньше (Ltal = 0.8 мм). M2 почти повторяет форму M1, но крупнее его; форма зуба сходна с M2 Paenohyaenoides (Hyainailourinae), который также имеет асимметричный тригонид (среди Hyaenodontidae, наиболее близкий по форме M2 у Protohyaenodon venturae, экз. USNM, № 214643): параконид ниже и короче протоконида; высота зуба больше, чем его длина; имеется второе режущее лезвие протоконида на его задней поверхности, обращенной к талониду. Желобок на передней поверхности зуба для талонида предстоящего зуба небольшой. Наибольшая ширина зуба приходится на задний край тригонида. Талонид режущий, имеет хорошо различимый посткристид. Отношение длины талонида М2 к длине зуба у Hyaenodon pumilus sp. nov. (Ltal/LМ2=0.196) близко к H. chunkhtensis. M3 имеет типичную для Hyaenodontinae форму: параконид равен по высоте протокониду. Протоконид сильно удлинен: соотношение длин протостилида и парастилида (lpr/lpa) равно 1.96, т.е. соотношение размеров составляет почти 2 : 1. У нового вида на М3 режущее лезвие по переднему краю параконида отсутствует, талонид отсутствует (нет даже рудиментарного талонидного выступа – небольшого пологого бугорка у основания протоконида). При этом задняя поверхность протоконида имеет выраженное режущее лезвие. Желобок на передней поверхности зуба для талонида предстоящего зуба выражен хорошо, он гораздо глубже, чем на М2. Лезвие параконида развернуто под небольшим углом к продольной оси зуба, и вершина параконида расположена лингвальнее хищнической вырезки. Зуб размещен в челюсти наклонно: он наклонен вперед – часть протоконида находится на основании венечного отростка (рис. 2). На Р2, Р4 и М1 хорошо заметен лингвальный цингулид. Лабиальный цингулид хорошо развит на передней части главного конуса Р4. Он также заметен под параконидом М1.
Таблица 1.
Размеры зубов Hyaenodon pumilus sp. nov. и Hyaenodon chunkhtensis, в мм
| Таксоны | Экземпляры | C1 | P1 | P2 | P3 | P4 | M1 | M2 | M3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | L | W | L | W | L | W | L | W | L | W | ||
| Hyaenodon pumilus sp. nov. | ПИН, № 3110/1217 (голотип) | 4.5 | 3.8 | 3.4* | 2.0* | 5.2 | 2.4 | 5.4* | 2.6* | 6.3 | 3.0 | 4.6 | 2.4 | 5.1 | 2.6 | 6.5 | 3.4 |
| Hyaenodon chunkhtensis | ПИН, № 3110/552 | – | – | 2.6* | – | 5.9* | 2.9* | 6.3 | 3.2 | 6.8 | 3.2 | – | – | – | – | – | – |
| Hyaenodon chunkhtensis | ПИН, № 3935/558 | – | – | – | – | – | – | – | – | 8.2 | 4.8 | 5.2 | 3.7 | 7.1* | – | – | – |
| Hyaenodon cf. chunkhtensis | ПИН, № 3110/618 | – | – | – | – | 7.1* | 3.5* | 9.1* | 3.6* | 9.1* | 3.1* | – | – | – | – | – | – |
| Hyaenodon chunkhtensis | PST, № 29-1 (голотип) | 6.5 | 4.4 | 6.2* | 2.7* | 7.0 | 3.4 | 7.5 | 4.1 | 7.3 | 4.2 | 5.1 | 3.0 | 6.3 | 3.7 | 10.6** | 4.1 |
| Hyaenodon chunkhtensis | PTS, № 29-2 | – | – | – | – | 6.0 | 3.0 | 7.5 | 3.8 | 7.5 | 4.0 | 5.6 | 3.4 | – | – | – | – |
| Hyaenodon chunkhtensis | MPC-M, № 30/51 | – | – | – | – | – | – | – | – | 6.9 | 3.2 | 4.6 | 2.4 | 6.1 | 2.8 | 8.4 | 3.6 |
Размеры в мм. Длина щечного зубного ряда P1‒M3 – 34.8; длина ряда премоляров P1–P4 – 19.4; длина M1–M3 – 15.4. Высота зубов (h): hМ3 (par) – 4.6; hМ3 (pr) – 3.7; hМ2 (par) – 4.1; hМ2 (pr) – 5.0; hМ1 – 3.9; hР4 – 6.6, hP2 – 4.9. Коэффициент относительного размера М1 (Km1 = 100 × × Lm1/Lm1–m3) равен 29.8. Коэффициент относительного размера М3 (Km3 = 100 × Lm3/Lm1–m3) составляет 42.2. Отношение длины ряда премоляров к длине моляров (LP1–P4/LM1–M3) у нового вида равно 1.26. Длина симфиза голотипа – 16.0. Высота горизонтальной ветви нижней челюсти составляет: между Р4 и М1 – 8.3; между М1 и М2 – 8.9; между М2 и М3 – 9.1. Длина параконида М3 равна 2.3, длина протоконида М3 – 4.5; отношение длины протоконида М3 к паракониду (lpr/lpa) равно 1.96. Отношение длины протоконида к длине М3 (lpa/LM3) у Hyaenodon pumilus – 0.692 (для сравнения у H. chunkhtensis оно равно 0. 64; Tsubamoto et al., 2008). Отношение высоты нижней челюсти между Р4 и М1 к длине щечного зубного ряда (KhD = KhM1/LP1–M3) составляет 0.238. Высота Р1 – 4.8 мм; высота Р4 – 7.0 мм; высота М1 – 3.7 мм. Другие измерения приведены в табл. 1 .
Сравнение. Hyaenodon pumilus sp. nov. отличается от всех гиенодонов Старого Света очень мелкими размерами и талонидом М1 с хорошо развитым посткристидом. От P. microdon новый вид отличается cиндосмозным симфизом нижней челюсти; от P. venturae он отличается более мелкими размерами, грацильной и более низкой, почти прямой горизонтальной ветвью нижней челюсти, слабо изогнутой вентрально линией нижнего щечного зубного ряда, отсутствием костного валика вдоль лабиовентрального края челюсти. Новый вид отличается от Hyaenodon chunkhtensis более мелкими размерами и низкой и грацильной горизонтальной ветвью (у H. chunkhtensis наиболее массивная нижняя челюсть среди всех карликовых видов), более компактным рядом премоляров, менее массивными зубами (в особенности Р4), удлиненными талонидами М1 и М2; Р1 сильно удлинен и его альвеола более чем в 1.5 раза длиннее альвеолы Р2 (у H. chunkhtensis альвеолы этих зубов равны по длине, табл. 1 ). У Hyaenodon pumilus задний край альвеолы Р1 расположен вплотную к альвеоле Р2, и параконид Р2 нависал над талонидом Р1; у H. chunkhtensis Р1 и Р2 разделены небольшой диастемой. Тригонид М1 длинный и низкий, в отличие от других гиенодонов.
Протоконид М3 у нового вида сильно удлинен относительно параконида: отношение их длины (lpr/lpa) равно 1.96. У Neohyaenodon cruentus, например, это отношение составляет 1.63. Горизонтальная ветвь нижней челюсти у Hyaenodon pumilus sp. nov. очень низкая и значение индекса KhD (0.238 – отношение высоты нижней челюсти между Р4 и М1 к длине щечного зубного ряда: KhD = KhM1/LP1–M3) является одним из наименьших в группе карликовых гиенодонов.
Новый вид отличается от Protohyaenodon raineyi вдвое меньшими размерами, размерами талонида и формой тригонида М1, более высокой горизонтальной ветвью; от H. filholi формой тригонида М1, мелкими размерами, грацильной нижней челюстью, однокорневым P1, ассиметричным тригонидом M2.
Отношение длины талонида к длине зуба у нового вида (Ltal/LМ1) – 0.210 (у H. chunkhtensis 0.8/5.1 = 0.156). В отличие от H. pumilus sp. nov., у H. filholi тригонид M2 более симметричный: высота и длина параконида почти равны размерам протоконида; пара- и протоконид не так глубоко разделены; экз. ГГM, № X -520/7. У H. pumilus sp. nov. протоконид М3 сильно удлинен (lpr/lpa = 1.96). У H. chunkhtensis число lpr/lpa составляет 1.8 (экз. MPC–M, № 30/51), у H. filholi – 1.7 (экз. MNHN, № Qu 8661), у P. crucians – 1.5 (экз. AMNH, 647), у Neohyaenodon horridus – 1.5, у Neohyaenodon sp. из МНР (экз. ПИН, № 3110/578) – 1.37. Талонид М2 имеет посткристид подобно талонидам M1‒M2 Schizophagus (Hyainailouridae; см. Lange-Badre, 1979, табл. 16 ). Отношение длины талонида М2 к длине зуба у H. pumilus sp. nov. (Ltal/LМ2 = = 1.0/5.1 = 0.196) близко к H. chunkhtensis (1.1/6.3 = 0.174), у нового вида относительная длина талонида немного больше (плезиоморфный признак).
Материал. Голотип.
* * *
Hyaenodon chunkhtensis Dashzeveg, 1985, голотип PST, № 29–1 (рис. 3; табл. X , фиг. 3). По сравнению с большинством других видов мелких гиенодонов, нижняя челюсть массивная и высокая. Такая же массивная челюсть среди этой группы имеется только у Protohyaenodon venturae. Коэффициент относительного размера М1 (KМ1 = = 100 × LМ1/LМ1–М3) равен 23.1 (реконструированное значение LМ3 – 10.6 мм для голотипа на основании соотношения длины параконида и протоконида для экз. MPC-M, № 30/51). Коэффициент относительного размера М3 (KМ3 = = 100 × LМ3/LМ1–М3) на экз. MPC-M, № 30/51 составляет 45.9 мм. Реконструируемая длина ряда моляров LM1–M3 – 21.3 мм, щечного ряда (LР1‒М3) – 48.5 мм. Высота челюсти между Р4 и М1 – 18.0 мм. Коэффициент массивности нижней челюсти KhD (отношение высоты нижней челюсти между Р4 и М1 к длине зубного ряда – KhМ1/LР1–М3) равен 0.371. Таким образом, нижняя челюсть у этого вида даже массивнее, чем у P. venturae (KhМ1/LР1–М3 = 0.339). У остальных пяти видов мелких Hyaenodontinae массивность нижней челюсти намного меньше (наибольшее значение из них у H. eminus – 0.282; минимальное – 0.203 у H. filholi). Задний край симфиза расположен на линии переднего края Р3. Поверхность эмали сильно ругозистая (на лингвальной поверхности Р4–М3 микрорельеф эмали хорошо сохранился).
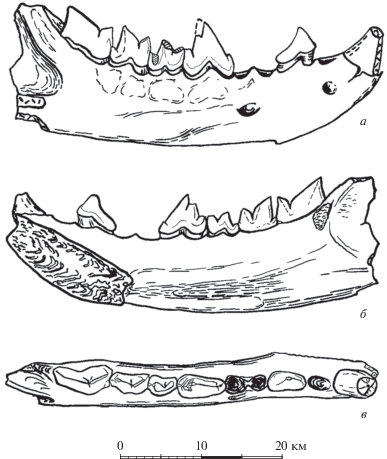
Рис. 3.
Hyaenodon chunkhtensis, голотип PST, № 29–1, левая ветвь нижней челюсти с С1, альвеолой Р1, Р2–М2 и фрагментом М3; Монголия, местонахождение Татал-Гол; олигоцен: а – с латеральной стороны; б – с медиальной стороны. Точками показаны места, реконструированные гипсом.
Лингвальный цингулид очень хорошо выражен на Р2–Р3 и немного слабее на Р4, на М1–М2 он развит под протоконидом и простирается назад до заднего края талонида. Лабиальный цингулид хорошо развит на Р3 и Р4, под протоконидом М2 и у заднего края параконида М3. М1 высокий и короткий. Его тригонид длинный, талонид предельно короткий. Протоконид М2 очень крупный, намного длиннее и выше параконида. Поэтому протостилид примерно в 2 раза длиннее парастилида. Талонид очень короткий, конический. У М3 отношение длин лезвий протоконида и параконида (lpr/lpa) равно 1.85, что устанавливается на основании образца MPC-M, № 30/51 (Tsubamoto et al., 2008). На этом основании нами указана реконструированная длина M3 для голотипа (табл. 1 ). В поперечном профиле горизонтальная ветвь имеет на лингвальной поверхности слабо выраженный продольный валик; максимальная толщина горизонтальной ветви челюсти слегка смещена в приальвеолярную часть. Под передним корнем Р4 наибольшая толщина челюсти находится на середине ее высоты и равна 6.3 мм.
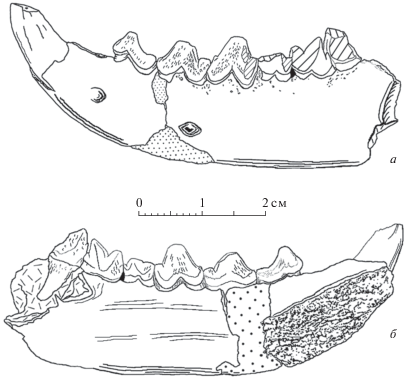
Hyaenodon cf. chunkhtensis, экз. ПИН, № 3110/552; местонахождение Хоер-Дзан; свита Эргилин-Дзо, верхний эоцен (рис. 4, а, б; табл. X , фиг. 2). Р1 однокорневой и, судя по альвеоле, по длине почти в 2 раза меньше, чем Р2; Р2 двухкорневой, в отличие от всех остальных образцов, поперечное сечение переднего корня в 2 раза меньше, чем заднего. Этот экз. выглядит мелкозубым по сравнению с образцами из олигоцена (голотип PST, № 29-1; экз. ПИН, № 3935/538; рис. 3–5). При этом, экз. ПИН, № 3110/552 и голотип имеют практически одинаковую высоту горизонтальной ветви нижней челюсти между Р3 и Р4 (13.3 и 13.1 мм, соответственно). При этом Р4 крупнее Р3: длина Р3 у образца из Хоер-Дзана составляет 84% длины Р3 у голотипа, а длина Р4 – 93% от длины этого зуба у голотипа. Длина ряда Р2–Р3 (Lр2 измерена по альвеоле) составляет у описываемого образца 19.1 мм, у голотипа – 21.6 мм (измерена также по переднему краю альвеолы Р2). Есть отличие в расположении заднего края симфизного шва: у голотипа он расположен у переднего края Р3, а у экз. ПИН, № 3110/552 он смещен вперед и расположен практически на линии оси главного конуса Р2 (табл. X , фиг. 5б). У экз. ПИН, № 3110/552 на Р3 талонидный выступ в 1.5 раза длиннее, чем у голотипа. В строении Р4, как и у голотипа, отмечается развитый параконид впереди от главного конуса. В отличие от голотипа, Р4 выглядит грацильным. Талонид Р4 длиннее, чем у голотипа, и сильнее отведен назад от главного конуса за счет большой ширины хищнической вырезки. Поверхность зубов образца сильно ругозистая, особенно на Р3. Лингвальный цингулид развит слабее лабиального, но присутствует на обоих премолярах. Лабиальный цингулид хорошо развит вдоль всей коронки на Р3 и Р4.
Рис. 4.
Hyaenodontinae из верхнего эоцена Монголии: а, б – Hyaenodon chunkhtensis, экз. ПИН, № 3110/552, правая ветвь нижней челюсти с Р3–Р4 и альвеолами Р1–Р2; Монголия, местонахождение Хоер–Дзан; свита Эргилин-Дзо, верхний эоцен; а – с латеральной стороны; б – с медиальной стороны; в–д – H. cf. chunkhtensis: в, г – экз. ПИН, № 3110/618, фрагмент левой ветви нижней челюсти с альвеолами Р2–Р4; Монголия, местонахождение Хоер-Дзан; свита Эргилин-Дзо, верхний эоцен; в – с латеральной стороны; г – с медиальной стороны; д – экз. SHG, № S/2 (слепок), схема поперечного сечения через правую горизонтальную ветвь нижней челюсти между М1 и М2 (вид с каудальной стороны); пунктиром показан контур М1. Обозначение: 1 – латеровентральный костный валик.
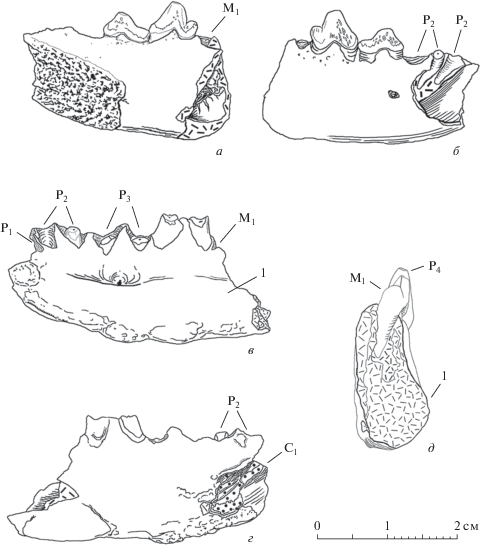
Рис. 5.
Hyaenodon chunkhtensis, экз. ПИН, № 3935/538, правая ветвь нижней челюсти с Р4–М2 и альвеолой Р3; Монголия, местонахождение Татал-Гол; свита Шанд-Гол, верхний олигоцен: а – с латеральной стороны; б – с медиальной стороны; в – сверху. Косой штриховкой показаны фасетки стирания зубов. Обозначения: 1 – антериорный корень М2; 2 – канал постериорного корня М2; 3 – медиальный костный валик; 4 – латеровентральное костное утолщение.
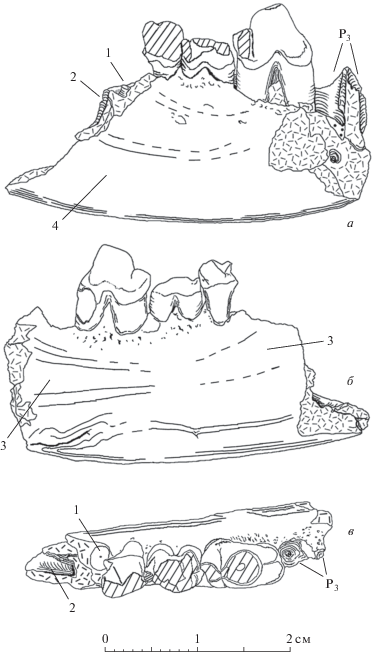
Подбородочное отверстие (for. mentalis) у экз. ПИН, № 3110/552 одно и находится на линии между Р2 и Р3 (у голотипа имеются 2 отверстия). Задний край симфиза расположен на линии оси главного конуса Р3. Толщина челюсти слабо увеличивается в вентральной части со стороны латеральной поверхности. Максимальная толщина челюсти, измеренная под передним корнем Р4, – 5.6 мм. В целом, отмеченные различия в размерах, массивности зубов и челюсти, расположении подбородочного отверстия между этим образцом и голотипом могут быть признаны не выходящими за рамки внутривидовой изменчивости.
Hyaenodon chunkhtensis, экз. ПИН, № 3935/538 (рис. 5; табл. X , фиг. 5); местонахождение Татал-Гол; свита Шанд-Гол (найден над верхним слоем базальтов, верхний олигоцен). В отличие от экз. ПИН, № 3110/552 и голотипа, он заметно крупнее и массивнее: длина ряда Р4–М1 – 13.4 мм, что составляет 109.8% (LP4–M1 у голотипа –12.2 мм). По форме Р4 экз. ПИН, № 3935/538 ближе к экз. ПИН, № 3110/552: главный конус также сильно наклонен назад, выступ талонида такой же удлиненный. В строении Р4 между этими образцами имеется различие: и зуб, и талонид у экз. ПИН, № 3935/538 массивные, подобно голотипу (талонид – самая широкая часть зуба). Отношение ширины Р4 к длине (W/L; табл. 1 ) для голотипа равно 0.575, у экз. ПИН, № 3935/538 – 0.585; у экз. ПИН, № 3110/552 этот показатель составляет 0.470. Из этих цифр видно, что по строению Р4 образец из Татал-Гола ближе к голотипу, чем к образцу из Хоер-Дзана. В строении М1 также отмечается увеличение массивности (отношение ширины зуба к длине – W/L): у экз. ПИН, № 3935/538 – 0.711. Она больше по сравнению с голотипом (W/L=0.588) и экз. ПИН, № 3110/552. На Р4 заметны почти уничтоженные стиранием следы микрорельефа ругозистой эмали. Можно заключить, что массивность зубов у этой формы имеет тенденцию к увеличению от находок из слоев из верхнего эоцена к находкам позднеолигоценового возраста.
Экз. ПИН, № 3935/538 отличается массивностью как челюсти, так и зубов. Высота горизонтальной ветви между Р3 и Р4 у голотипа – 13.1 мм, у экз. ПИН, № 3935/538 – 17.0 мм, что больше почти на треть (129.8%). Увеличение высоты горизонтальной ветви челюсти на 30% не случайно. На медиальной поверхности ветви челюсти, в отличие от голотипа, хорошо выражен костный валик вдоль продольной оси челюсти (рис. 5, б). Такой продольный костный валик на медиальной поверхности ветви челюсти типичен для крупных Hyaenodontinae (Neohyaenodon). У карликовых видов костный валик до сих пор был отмечен только у Protohyaenodon venturae – вдоль латеровентрального края челюсти. Максимальная толщина горизонтальной ветви на линии переднего корня Р4 – 9.1 мм. Имеется пологий валик и вдоль латеровентрального края горизонтальной ветви (рис. 5, а).
Таким образом, образец H. chunkhtensis из верхнего олигоцена (экз. ПИН, № 3935/538) отличается от голотипа: 1) более крупными размерами (примерно на 10%); 2) большей массивностью (на 30%). Увеличение массивности челюсти проявляется не только количественно, но и в появлении костных валиков на медиальной поверхности горизонтальной ветви (укрепляют механическую конструкцию челюстного аппарата). Можно заключить, что эти две тенденции развития были ключевыми в этой линии Hyaenodontinae на протяжении 10 млн лет своего существования.
Hyaenodon cf. chunkhtensis, экз. ПИН, № 3110/618 (рис. 4, в, г; табл. X , фиг. 4) – фрагмент левой горизонтальной ветви нижней челюсти с задней частью альвеолы С1 и альвеолами с фрагментами зубных коронок и корней Р2–Р4. Альвеола корня, следующая за Р4, неглубокая т.е. принадлежала небольшому зубу – М1 (передний корень М1 короткий, в отличие от корней премоляров).
Дорсальная часть челюсти впереди от Р2 не сохранилась. Судя по близости заднего края альвеолы С1, Р1 был небольшим и однокорневым (рис. 4, в). Корень С1 был очень слабо наклонен вперед. По-видимому, коронка Р1 также имела наклон, в отличие от голотипа H. chunkhtensis. Длина Р2 по альвеоле – 7.1 мм, что близко к размерам голотипа, как и большинство других промеров (табл. 1 ). Форма альвеол Р3 идентична с голотипом и соответствует симметричной коронке зуба. Отличия состоят в том, что Р3 и Р4 почти равны по длине и крупнее, чем у голотипа, на 21–28%. Высота горизонтальной ветви между Р3 и Р4 составляет 16.6 мм, между Р4 и М1 – 17.2 мм, что составляет 122.8% от соответствующего размера голотипа (у него последний промер равен 14.0 мм).
На латеральной поверхности горизонтальной ветви, вдоль ее вентрального края, имеется костный валик, начинающийся от переднего края Р2, достигающий М1 и уходящий далее в каудальном направлении (рис. 4). Дорсально костный валик достигает вентрального края ментального отверстия. Костный валик очень мощный и образует полку, ограничивающую с вентральной стороны выход for. mentalis: этот валик с вентральной стороны образует подобие синуса вокруг ментального отверстия. Поверхность костного валика имеет крупно-бугристую текстуру. Костные бугры развиты и на вентральной половине медиальной поверхности горизонтальной ветви (возможно, что такая текстура поверхности – проявление остеофитов, которые сформировались в результате заболевания – напр., остеомиелитом). Поверхность симфизного шва разрушена, но, судя по ряду признаков, его каудальный край должен был располагаться на линии между Р2 и Р3. Подбородочное отверстие одно и расположено на линии оси Р3.
ОБСУЖДЕНИЕ
Hyaenodon pumilus sp. nov. характеризуется мелкими размерами, синдосмозным симфизом, задний край которого достигает средней оси Р3; горизонтальная ветвь нижней челюсти низкая, грацильная, слабо изогнута, P1 однокорневой; тригонид M1 необычно низкий и длинный, а талонид удлинен, по сравнению с М1 у других гиенодонтин (при этом он подобен талониду М1 только у H. chunkhtensis). Посткристид М1 у нового вида хорошо развит. У H. pumilus M2 характеризуется коротким талонидом с выраженным посткристидом. Значение KM1 – около 30; KM3 – около 42, степень удлинения премоляров относительно ряда моляров (LP1–P4/LM1–M3) составляет около 1.25–1.26.
Исследование H. pumilus sp. nov. и H. chunkhtensis подтверждает ранее сделанное нами наблюдение, что у мелких видов гиенодонов длина М1 относительно длины ряда моляров больше, чем у более крупных видов, а относительный размер М3 меньше (Lavrov, Emry, 1998). У H. pumilus sp. nov. КМ1=100 × LM1/LM1–M3 составляет 29.8, что близко к показателю у P. venturae (28–30). У H. chunkhtensis КМ1 (23.1–25.0) – как у H. filholi и H. minor (25.8 и 24.4 соответственно). Таким образом, у H. pumilus sp. nov. M1 редуцирован слабее, по сравнению с H. chunkhtensis. При этом у H. pumilus sp. nov. относительные размеры М3 в ряду моляров небольшие, и КМ3 составляет 42.2 (как у H. filholi – 41.0–41.5). У H. chunkhtensis М3 в ряду моляров заметно увеличен (KM3–45.9) – почти как у крупных видов гиенодонтин (у Neohyaenodon montanus – 46.3; у Hyaenodon dubius – 47.4).
Степень удлинения ряда премоляров (отношение LP1–P4/LM1–M3) у H. chunkhtensis (голотип PST, № 29-1) равно 1.39 (LP1–P4 – 28.4; реконструированные LM3 – 10.2; LM1–M3 – 20.3): это существенно больше, чем у H. pumilus sp. nov. (1.26). H. filholi обладает наиболее удлиненным лицевым отделом по сравнению с другими карликовыми видами, и наиболее низкой нижней челюстью (KhD – 0. 203). Ряд премоляров у него более удлинен: соотношение LP1–P4/LM1–M3 равно 1.45. У P. raineyi индекс KhD составляет 1.35 (экз. TMM, № 40504-236). Следует отметить, что наиболее коротким рядом премоляров (и лицевым отделом) среди Hyaenodontinae обладает некрупный H. minor, особенно H. cf. minor с острова Уайт (Сray, 1973) – у него длина ряда премоляров почти равна длине ряда моляров: значение LP1‒P4/LM1–M3 близко к 1. Можно заключить, что степень удлинения лицевого отдела у H. pumilus sp. nov. была меньше, чем у H. chunkhtensis. Степень его удлинения была промежуточной между H. minor и P. raineyi. У голотипа H. pumilus sp. nov. М1 оказался слабо стертым при полностью прорезавшемся М3. Такая хорошая сохранность М1 не типична для гиенодонов. Из этого можно заключить, что у М1 было позднее прорезание, и он не функционировал в составе смены молочных зубов, как это имело место у других видов Hyaenodontinae (Balst et al., 2011).
У H. pumilus sp. nov. на нижних зубах сохраняется выраженный лингвальный цингулид, как у H. chunkhtensis (экз. ПИН, № 3110/552), а лабиальный развит слабее (как у экз. ПИН, № 3110/552). У H. pumilus sp. nov. лабиальный цингулид есть только на премолярах (Р2 и Р4) и под параконидом М1. У всех исследованных образцов H. chunkhtensis лабиальный цингулид развит слабо на премолярах. У H. filholi лингвальный цингулид развит на премолярах в той же степени, как у азиатских форм. Следует отметить особое положение экз. ПИН, № 3110/552. С одной стороны, мы видим определенное сходство с H. pumilus sp. nov. (развитый лигвальный цингулид премоляров), с другой – форма Р2 у этого экз. идентична Р2 голотипа H. chunkhtensis. У экз. ПИН, № 3110/552 имеется фрагмент двухкорневого Р1, что не характерно для группы карликовых гиенодонов. Только у H. filholi Р1 двухкорневой, что является для карликовых гиенодонов плезиоморфным признаком.
У H. pumilus sp. nov. отношение высоты М1 к его длине (hМ1/LМ1 × 100 = 3.7/4.6) составляет 80.4. Дашзевэг для характеристики габитуса М1 у H. chunkhtensis приводит отношение 85.7 (Dashzeveg, 1985). М1 у H. pumilus sp. nov. низкий по сравнению с М1 у H. chunkhtensis: при близких значениях длины зуба (табл. 1 ) высота нестертого тригонида у H. pumilus sp. nov. равна высоте сильно стертого тригонида H. chunkhtensis (3.8 мм). Низкое значение, приведенное Дашзевэгом, объясняется сильной стертостью М1 у голотипа H. chunkhtensis. Если реконструировать форму нестертого М1 у H. chunkhtensis, то коэффициент габитуса должен быть близок к 100 – т.е., он значительно более компактный (короткий и высокий) по сравнению с H. pumilus sp. nov. Талонид М1 у H. pumilus sp. nov. имеет рудиментарный посткристид. Рудиментарный посткристид талонида имеется и у Protohyaenodon venturae. У высокоспециализированных хищников это является архаическим признаком.
У Protohyaenodon microdon и P. venturae, как и у европейского H. filholi, строение талонида М1 сходно с H. chunkhtensis, он короткий и режущий. Талонид М2 у всех карликовых гиенодонов редуцирован почти так же, как у H. chunkhtensis. Но у последнего вида типичный короткий режущий талонид М2 гиенодонов трансформируется в еще более короткий конический талонид колющего типа. Редукция талонидов М1–М2 является диагностической особенностью H. chunkhtensis. В строении тригонидов обращает на себя внимание значительное удлинение протостилида относительно парастилида на М3 у мелких видов гиенодонов, что выражается в отношении lpr/lpa (степень развития протоконида этого зуба выражается также через индекс lpr/LM3; Tsubamoto et al., 2008). Этот индекс отражает, насколько режущее лезвие протоконида (протостилид) длиннее режущего лезвия параконида (парастилид). У Neohyaenodon (N. horridus, N. cruentus) и крупных видов Protohyaenodon (P. crucians) значение lpr/lpa составляет около 1.5–1.6. У мелких видов гиенодонов протостилид значительно удлинен. Относительная длина протостилида нарастает в ряду Hyaenodon filholi, H. chunkhtensis, H. pumilus sp. nov. от 1.7 до 1.96. Удлинение длины режущих лезвий зубов отражает повышение секториальной специализации моляров и является проявлением гиперкарниворной специализации. H. pumilus sp. nov., отставая от других мелких видов в специализации М1 (форма тригонида, размеры и наличие посткристида на талониде), заметно опережает другие виды в специализации М3. Специализация заключалась в удлинении протостилида М3 и окклюдирующего с ним метастиля М2 – за счет развития этих двух лезвий совершенствовался механизм разрезания главной пары хищнических зубов М2/М3.
Hyaenodon pumilus sp. nov. имеет ряд архаичных признаков: морфология тригонида М1 (близка к форме DP4 большинства гиенодонов), развитый талонид, хорошее развитие цингулидов премоляров и М1. Эти плезиоморфные признаки позволяют предположить, что новый вид морфологически близок к предковой форме мелких гиенодонов, общей для азиатских видов карликовых гиенодонов (H. pumilus sp. nov., H. chunkhtensis, H. eminus).
В отличие от P. microdon, имеющего синостозный симфиз, P. venturae и азиатские формы демонстрируют другой набор апоморфных признаков, указывающих на усиление конструкции челюстного аппарата: костный валик вдоль горизонтальной ветви нижней челюсти, большая высота горизонтальной ветви. Кроме того, у H. chunkhtensis отмечается усиление структуры эмали премоляров, которая внешне выражается в ругозистой поверхности эмали премоляров.
Следует обратить внимание на H. chunkhtensis, экз. MPC–M, № 30/51 из местохождения Их-Дзан (рис. 1); свита Эргилин-Дзо, верхний эоцен (Tsubamoto et al., 2008). Материал представляет фрагмент левой горизонтальной ветви нижней челюсти с Р4–М3; вентральная часть челюсти не сохранилась. Этот образец характеризуется меньшими размерами и менее массивными Р4–М2 по сравнению с образцами из местонахождения Шункт (рис. 1, 4). При этом М2 у экз. MPC-M, № 30/51 более крупный не только относительно размеров М1, но М2 физически крупнее, чем у голотипа (табл. 1 ). Отношение W/L для М2 у экз. MPC-M, № 30/51 составляет 0.460, у голотипа – 0.587. Отношение длины М2 к М1 (LМ2/LМ1) для экз. MPC-M, № 30/51 равно 1.326, у голотипа – 1.235. Таким образом, у экземпляра MPC-М, № 30/51 размеры М1 мельче относительно М2, по сравнению с голотипом, что согласуется с установленными закономерностями для размеров моляров у мелких форм (Lavrov, Emry, 1998). Вентральная часть челюсти у экз. MP-М, 30/51 не сохранилась, и неизвестно, имелся ли костный вентролатеральный бортик на горизонтальной ветви.
Среди образцов мелких гиенодонов из Монголии обращает на себя внимание экз. ПИН, № 3110/618, определенный как H. cf. chunkhtensis. Для него характерны массивный продольный костный валик вдоль латеровентрального края горизонтальной ветви нижней челюсти. Этот валик похож на таковой у P. venturae. Но у американского вида этот костный валик меньше выражен, и не достигает симфизной части челюсти в переднем направлении. Отметим, что для крупных Hyainailouridae характерен продольный медиодорсальный костный валик, расположенный под линией альвеол и являющийся дополнительным усилителем конструкции нижней челюсти. Основную роль по усилению челюсти у P. venturae выполняет общая форма нижней челюсти (горизонтальная ветвь короткая, высокая и изогнутая). У H. cf. chunkhtensis костный валик выражен сильнее, чем у P. venturae, и достигает симфиза; высота горизонтальной ветви больше, чем у остальных мелких видов гиенодонов, но меньше, чем у P. venturae. Поэтому мощный костный валик у экз. ПИН, № 3110/618 должен рассматриваться как диагностический признак видового ранга. Такое же строение нижней челюсти отмечается и у H. cf. сhunkhtensis, экз. SHG, № S/2 (рис. 4, д; Татал-Гол, свита Шанд-Гол, нижний олигоцен; ранее был определен как H. eminus; Morlo, Nagel, 2006). Челюсть имеет мощный выступ по латеровентральному краю, где толщина горизонтальной ветви примерно в 1.5 раза больше, чем в средней части профиля челюсти.
Следует учесть, что от азиатских форм известны только немногочисленные фрагменты горизонтальных ветвей нижней челюсти, т.е., крайне бедный материал. Возможно, экз. ПИН, № 3110/618 и SHG, № S/2 принадлежат другому виду карликовых азиатских гиенодонов с робустной челюстью, морфологически более близкому к американскому P. venturae, чем к H. chunkhtensis. Своими низкими и прямыми нижними челюстями остальные карликовые виды сильно отличаются от P. venturae, у которого челюсть высокая, а изгиб горизонтальной ветви и зубного ряда ярко выражен.
Гиперкарниворная пищевая специализация Protohyaenodon venturae и Hyaenodon chunkhtensis выражена в механическом усилении челюстного аппарата. Мы может констатировать для этих видов специализацию к макрофагии. По-видимому, мелкие гиенодоны, подобно ряду современных мелких Carnivora (Mustelidae: Martes zibellina L. и другие виды), были в состоянии справляться с добычей, равной себе по размерам. У P. microdon, несмотря на его наименьшие в подсемействе размеры, эта специализация (механическое усиление нижней челюсти) выражена в формировании синостозного симфиза. Гиперкарниворная специализация среди мелких гиенодонов, как и среди крупных форм, является одной из генеральных линий направления эволюции. По сравнению с мелкими Carnivora, гиперкарниворная специализация у мелких Hyaenodontinae имеет иное морфологическое выражение. У мелких гиенодонов такая специализация в строении зубов выражена намного меньше. Эта специализация выражена в усилении конструкции челюстного аппарата. У мелких Carnivora (Mustelidae) гиперкарниворная специализация выражена, прежде всего, в сильном увеличении относительного размера М1 по сравнению с остальными зубами (например, у Martinogale – см. Dunkle, 1938). Увеличение размеров хищнических зубов у креодонтов (и у мелких гиенодонтин, в частности) было затруднено положением главного хищнического зуба в конце зубного ряда. Такое положение делает невозможным увеличить длину за счет приближения его заднего края к оси вращения челюстного сустава, так как сокращение рычага приложения силы сделает работу челюстной мускулатуры не эффективной (Greaves, 1983). Увеличению размера хищнического зуба в антериорном направлении препятствуют предстоящие зубы, и увеличение М3 связано с редукцией М1. В строении зубов у мелких форм гиперкарниворная специализация выражается в развитии ругозистой эмали, что является внешним отражением усиления ее внутренней структуры для высоких нагрузок – разгрызания костей (премоляры P. venturae, H. filholi, премоляры и моляры Р2–Р4 H. chunkhtensis).
Признаки гиперкарниворной спрециализации среди мелких гиенодонов отмечаются у шести из семи видов. Обнаруживается параллелизм в направлении эволюции мелких форм хищников у Creodonta и Carnivora – направление на гиперкарниворную специализацию для значительной части форм. Разные стратегии достижения такой хищнической специализации у Creodonta и Carnivora обусловлены принципиальными различиями в анатомии черепа и челюстного аппарата у этих отрядов. Тип гиперкарниворной специализации мелких гиенодонов является ярким примером адаптивного компромисса (Расницын, 1987), реализуемого креодонтами. Для ряда карликовых форм характерно сохранение плезиоморфных признаков: рудиментарных бассейнов талонидов, цингулидов, параконидов премоляров и, возможно, позднее прорезание М1.
Анализ материала по карликовым гиенодонам Центральной Азии показывает, что в позднем палеогене существовало 4 типа этих мелких хищников. Первый тип (самый мелкий) представлен H. pumilus sp. nov., второй и третий типы из позднего эоцена и олигоцена объединены под видовым именем H. chunkhtensis. Четвертый тип – более крупный карликовый вид с грацильной низкой челюстью – H. eminus. Он характеризуется рудиментарным однокорневым Р1 и низкой грацильной нижней челюстью (подобно Protohyaenodon raineyi). Отмеченные отличия в морфологии нижней челюсти между H. chunkhtensis и H. cf. chunkhtensis (экз. ПИН, № 3110/618 и SHG, № S/2) могут означать, что, возможно, здесь существовал другой вид мелких гиенодонов с более массивным челюстным аппаратом, чем у H. chunkhtensis. Для позднеэоценовых экземпляров отмечены меньшие размеры, меньшая массивность зубов и челюсти (экз. ПИН, № 3110/552), чем у олигоценовых форм. Эволюционными трендами мелких гиенодонов является гиперкарниворная специализация и формирование однокорневого Р1. Поэтому форму из позднего эоцена (экз. ПИН, № 3110/552) с двухкорневым Р1 можно рассматривать как предковую для олигоценовых H. chunkhtensis, у которых Р1 однокорневой.
Фрагментарность экз. ПИН, № № 3110/552, 3935/538, 3110/618, MPC-M, № 30/51 и SHG, № S/2 не позволяет надежно произвести таксономическое разделение этих находок. В настоящее время целесообразнее все находки морфологического типа карликовых гиенодонов Азии с массивным челюстным аппаратом относить к виду H. chunkhtensis. Находки карликовых гиенодонов в Азии более фрагментарны и редки, чем в Северной Америке и Европе. Знания об этой азиатской группе очень малы. Новые находки должны разрешить проблемы морфологии и таксономического разнообразия мелких креодонтов Центральной Азии.
Список литературы
Расницын А.П. Темпы эволюции и эволюционная теория (гипотеза адаптивного компромисса) // Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 46–64.
Balst K., Morlo M., Nagel D., Heizmann E. Differences in the tooth eruption sequence in Hyaenodon (Creodonta: Mammalia) and implications for the systematics of the genus // J. Vertebr. Paleontol. 2011. V. 31. № 1. P. 181–192.
Berkey C.P., Granger W. Later sediments of the desert basins of Central Mongolia // Amer. Mus. Novit. 1923. № 77. P. 1–77.
Cray P.E. Marsupialia, Insectivora, Primates, Creodonta and Carnivora from Headon Beds (Upper Eocene) of Southern England // Bull. Brit. Mus. (Natur. Hist.) Geol. 1973. V. 23. № 1. 102 p.
Dashzeveg D. On two Oligocene Hyaenodontidae from Erghilyin-dzo (Mongolian People`s Republic) // Acta Palaeontol. Pol. 1964. V. 9. № 2. P. 263–276.
Dashzeveg D. Nouveaux Hyaenodontinae’s (Creodonta, Mammalia) du Paléogene de Mongolie // Ann. Paléontol. 1985. V. 71. № 4. P. 223–256.
Daxner-Höck G., Badamgarav D., Erbaeva M. Oligocene stratigraphy based on a sediment-basalt association in Central Mongolia (Taatsiin gol and Taatsin Tsagaan Nuur area, Valley of Lakes): review of a Mongolian-Austrian project // Vertebr. PalAsiat. 2010. V. 48. № 4. P. 348–366.
Dunkle D.H. A lower jaw of Martinogale alveodens // Univ. Kansas. Sci. Bull. 1938. V. 25. № 8. P. 181–185.
Fritzell E.K., Haroldson K.J. Urocyon cineaeoargenteus // Mammal. spec. 1982. № 189. P. 1–8.
Greaves W. S. A functional analysis of carnassial biting // Biol. J. Linn. Soc. 1983. V. 20. № 4. P. 267–274.
Gustafson E.P. Carnivorous mammals of the late Eocene and early Oligocene of Trans-Pecos, Texas // Texas Mem. Mus. Bull. 1986. V. 33. P. 1–66.
Lange-Badre B. Les Creodontes (Mammalia) d’Europe occidentale de l’Eocéne supèrieur a l’Oligocéne supèrieur // Mém. Mus. Natn. Hist. Natur. 1979. Sér. C. V. 42. 249 p.
Lavrov A.V., Emry R.J. Hyaenodon venturae (Hyaenodontidae, Creodonta, Mammalia) from Early Chadronian (latest Eocene) of Wyoming // J. Paleontol. 1998. V. 72. № 4. P. 752–757.
Matthew W., Granger W. New mammals from the Irdin Manha Eocene of Mongolia // Amer. Mus. Novit. 1925. V. 198. P. 1–10.
Mellett J.S. Paleobiology of North American Hyaenodon (Mammalia, Creodonta) // Contr. Vertebr. Evol. 1977. V. 1. 134 p.
Morlo M., Nagel D. New remains of Hyaenodontidae (Creodonta, Mammalia) from the Oligocene of Central Mongolia // Ann. Paléontol. 2006. V. 92. P. 305–321.
Polly P.D. The skeleton of Gazinocyon vulpeculus gen. et comb. nov. and the cladistic relationships of Hyaenodontidae (Eutheria, Mammalia) // J. Vertebr. Paleontol. 1996. V. 16. № 1. P. 303–319.
Tsubamoto T., Watabe M., Tsogtbaatar K. Hyaenodon chunkhtensis and the hyaenodontid fauna from the Upper Eocene Ergilin Dzo formation of Mongolia // J. Vertebr. Paleontol. 2008. V. 28. № 2. P. 559–564.
Wang X., Qui Z., Wang B. Hyaenodonts and carnivorans from the early Oligocene to early Miocene of Xianshuihe Formation, Lanzhou basin, Gansu Province, China // Paleontol. Electron. 2005. V. 8:6A. P. 1–14.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал