

Палеонтологический журнал, 2019, № 4, стр. 31-42
Классификация позднепалеозойского семейства Medlicottiidae Karpinsky (Ammonoidea)
Т. Б. Леонова *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: tleon@paleo.ru
Поступила в редакцию 20.11.2018
После доработки 26.11.2018
Принята к публикации 06.02.2019
Аннотация
В статье кратко рассматривается изменение взглядов на систему и филогенетические связи в семействе Medlicottiidae Karpinsky. Критически обсуждается последняя версия, предложенная в ревизованном издании “Treatise …” (Furnish et al., 2009). На основании ревизии старых и изучения новых данных приводится обоснование принимаемой автором классификации, уточняются диагнозы родов Paramedlicottia, Parasicanites, Miklukhoceras и границы временного распространения Neouddenites и Medlicottia. Ревизуются модели развития отдельных филогенетических ветвей медликоттиид в подсемействах Medlicottiinae, Sicanitinae, Episageceratinae.
ВВЕДЕНИЕ.
Семейство Medlicottiidae Karpinsky, 1889, номинативное для подотряда Medlicottiina Yu. Zakharov (Захаров, 1983), является наиболее своеобразной и сложной в морфологическом плане группой в составе палеозойского отряда аммоноидей Prolecanitida.
Классификация и эволюция Medlicottiidae была подробно рассмотрена В.Е. Руженцевым 70 лет назад (Руженцев, 1949). Основой исследования послужили уникальные по полноте и сохранности коллекции медликоттиид, собранные на Южном Урале, которые позволили детально проследить филогенез семейства на протяжении четырех геологических веков и обосновать морфологические основы его классификации. Ни в одном другом регионе мира представители этого семейства не были найдены в такой длительной временной последовательности и в таком разнообразии. В монографии было проанализировано 13 родов медликоттиид. Кроме этого, нескольким малоизученным родам был придан статус семейств. Это Episageceratidae Ruzhencev (три рода, три вида), Sundaitidae Ruzhencev (один род, один вид) и Shikhanitidae Ruzhencev (один род, один вид). Родовой состав семейства показан на таблице 1. Подсемейственные категории в этой работе выделены не были.
Таблица 1.
Классификационные схемы медликоттиид
| Руженцев, 1949 | “Tretiase…” Miller et al., 1957 | Руженцев, 1960 | “Основы …” Руженцев, 1962 | Leonova, 2002 | “Tretiase…” Furnish et al., 2009 |
|---|---|---|---|---|---|
| Семейство Medlicottiidae | |||||
| Prouddenites Uddenites Daixites Artioceras Synartinskia Sicanites Propinacoceras Akmilleria Aktubinskia Artinskia Medlicottia Eumedlicottia Neogeoceras | п/сем. Uddenitinae Prouddenites Uddenites Uddenoceras п/сем. Melicottiinae Medlicottia Artinskia Propinacoceras Episageceras Sicanites |
п/сем Uddenitinae Prouddenites Uddenites Daixites Uddenoceras п/сем Sicanitinae Artioceras Synartinskia Sicanites Propinacoceras Akmilleria п/сем Medlicottiinae Artinskia Medlicottia Eumedlicottia Aktubinskia Neogeoceras |
п/сем Uddenitinae Prouddenites Uddenites Daixites Uddenoceras Neouddenites п/сем Sicanitinae Artioceras Propinacoceras Synartinskia Sicanites Akmilleria п/сем Medlicottiinae Artinskia Aktubinskia Medlicottia Eumedlicottia Neogeoceras |
п/сем Uddenitinae Prouddenites Daixites Uddenites Uddenoceras Neouddenites п/сем Medlicottiinae Artinskia Paramedlicottia Medlicottia Eumedlicottia Akmilleria Neogeoceras Syrdenites п/сем Propina-coceratinae Propinacoceras Bamyaniceras Difuntites п/сем Sicanitinae Vanartinskia Synartinskia Prosicanites Sicanites Parasicanites Aktubinskia п/сем Artioceratidae Artioceras Artioceratoides п/сем Miklukhoceratinae Miklukhoceras |
п/сем Uddenitinae Uddenites Daixites Neouddenites Prouddenites, Uddenoceras п/сем Medlicottiinae Medlicottia Artinskia Eumedlicottia Neogeoceras Syrdenites п/сем Propinacoceratinae Propinacoceras Akmilleria Bamyaniceras Darvasiceras Difuntites Kunlunoceras Miklukhoceras п/сем Sicanitinae Sicanites Artioceras Artioceratoides Synartinskia п/сем. Episageceratinae Episageceras Latisageceras Nodosageceras |
| “Особые роды” Episageceras, Sundaites | Сем. Episageceratidae Episageceras Latisageceras Nodosageceras | Сем. Episageceratidae
Episageceras
Latisageceras
Nodosageceras Protosageceras |
Сем. Episageceratidae Episageceras Latisageceras Nodosageceras | ||
В американской сводке “Tretiase…” (Miller et al., 1957) в составе семейства было выделено два подсемейства: Uddenitinae Miller et Furnish, 1940 в составе трех родов и Medlicottiinae Karpinsky, 1889 в составе пяти родов (табл. 1), остальные роды, описанные в работе Руженцева (1949) были сведены в синонимику или вообще не упомянуты.
Позднее Руженцев (1960) подразделил семейство Medlicottiidae на три подсемейства: Uddenitinae (четыре рода), Sicanitinae Noetling, 1904 (пять родов) и Medlicottiinae (пять родов) (табл. 1). Практически та же классификация медликоттиид приведена в “Основах …” (Руженцев и др., 1962), она отличается лишь появлением вновь установленного рода Neouddenites Ruzhencev, 1961 в подсемействе Uddenitinae (всего пять родов).
В сводке по пермским аммоноидеям автора этой статьи (Leonova, 2002) была кардинально переработана система медликоттиид. Основой для такой ревизии послужили исследования богатейших коллекций из ассельско-сакмарского и, особенно, артинского и кунгурского ярусов Памира и Дарваза (Руженцев, 1978; Леонова, 1984, 1985; Леонова, Дмитриев, 1989; Левен и др., 1992). Семейство принято в составе 25 родов и подразделено на шесть подсемейств (табл. 1), каждое из которых характеризовалось своеобразным путем развития (Леонова, Дмитриев, 1989). Подсемейство Uddenitinae рассмотрено в составе пяти родов, так же как и у Руженцева и др. (1962). Номинативное подсемейство Medlicottiinae в составе восьми родов было дополнено родом Syrdenites Nassichuk, Furnish et Glenister, 1965 и вновь выделенным Paramedlicottia Leonova, 1992; также в него был помещен род Akmilleria Ruzhencev, 1940. В подсемейство Sicanitinae включено шесть родов: кроме ранее рассматриваемых в его составе Sicanites и Synartinskia, были введены Prosicanites Toumanskaya, 1949; Vanartinskia Ruzhencev, 1978; Parasicanites Leonova, 1985 и Aktubinskia Ruzhencev, 1947. Восстановлено подсемейство Propinacoceratinae Plummer et Scott, 1937, которое предыдущими авторами рассматривалось как синоним Sicanitinae. Его состав определен следующим образом: Propinacoceras Gemmellaro, 1887; Bamyaniceras Termier et Termier, 1970 и Difuntites Glenister et Furnish, 1988. Также было выделено два новых малочисленных подсемейства, резко отличающихся от всех других медликоттиид. Это Artioceratinae Leonova с двумя родами Artioceras Ruzhencev, 1947 и Artioceratoides Leonova, 1985, а также Miklukhoceratinae Leonova с одним родом Miklukhoceras Pavlov, 1967 (плохо изученный китайский род Kunlunoceras Wang, 1983 принимался как синоним Miklukhoceras).
В ревизованном издании “Tretiase…” (Furnish et al., 2009) семейство Medlicottiidae рассматривается в составе пяти подсемейств: Uddenitinae, Medlicottiinae, Propinacoceratinae, Sicanitinae и Episageceratinae. Единственным принципиальным новшеством предложенной системы было введение Episageceratinae в качестве подсемейства в состав Medlicottiidae (табл. 1). В отличие от рассматриваемой выше работы (Leonova, 2002), авторы не признают два малочисленных подсемейства Artioceratinae и Miklukhoceratinae, а представляющие их роды помещают в состав других подсемейств. В подсемейство Propinacoceratinae вводятся Miklukhoceras и Kunlunoceras (при этом последний род признается валидным, несмотря на очень плохую сохранность единственного, найденного на сегодняшний день, экземпляра и нечеткое его описание). В Sicanitinae переносятся Artioceras и Artioceratoides. Роды Prosicanites, Vanartinskia и Aktubinskia сводятся в синонимику рода Sicanites, тем самым практически уничтожая его диагноз. Род Parasicanites рассматривается в качестве синонима Synartinskia, несмотря на четкие морфологические различия этих двух таксонов (Леонова, 1985). По-видимому, такая трактовка связана с тем, что авторы ревизии не имели возможности лично ознакомиться с коллекционным материалом по этим редко встречающимся формам. Чтобы избежать этого недопонимания, в предлагаемой статье приводятся более полные сравнительные материалы для родов Parasicanites и Synartinskia; Paramedlicottia, Medlicottia и Artinskia.
Медликоттииды, в отличие от других пермских палеозойских аммоноидей, занимают очень небольшую, четко ограниченную морфологическую область 4А (по классификации: Barskov et al., 2008) в “рауповском” морфопространстве, т.е., все они имеют платиконовую форму раковины с узким умбиликом и характерную лопастную линию со сложно-рассеченным наружным седлом и многочисленными, убывающими в размерах лопастями на боковой стороне. В предлагаемой статье мы не будем останавливаться на вопросах происхождения этих лопастей, являются ли они боковыми или умбиликальными (Леонова, 2012). В данном случае важно отметить основные диагностические признаки, которые можно использовать для наиболее объективной классификации таксонов, входящих в эту группу. Такие признаки относятся к особенностям строения лопастной линии и внешней морфологии раковины. (1) Строение наружного седла с системой адвентивных лопастей. Практика показывает, что этот признак очень строго выдерживается в пределах таксона родовой категории. (2) Положение основания первой боковой лопасти по отношению ко второй и третьей (более глубокое положение первой боковой лопасти по отношению к основанию второй свидетельствует о более древнем возрасте вида, также может характеризовать отдельные филогенетические ветви). (3) Общее число боковых лопастей и число двураздельных боковых лопастей (как правило, этот признак хорошо коррелирует с эволюционным положением изучаемой формы в филогенетическом ряду). (4) Относительная ширина умбилика, степень инволютности раковины (характер и скорость изменения этих параметров в онтогенезе являются диагностическими признаками для каждого рода). (5) Ширина вентральной стороны. (6) Наличие и характер скульптурных элементов (килей или бугорков), расположенных, главным образом, на вентральной стороне и вентро-латеральном перегибе.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В последние десятилетия не было получено новых данных о составе и морфогенезе подсемейства Uddenitinae (Uddenites, Daixites, Prouddenites, Uddenoceras, Neouddenites), но появились сведения о более широком распространении рода Neouddenites. Представители этого рода были обнаружены в казанских отложениях Волго-Урала. В 2000 г. в карьере Кремешки казанскими геологами была найдена ювенильная форма, которая определялась как ?Neouddenites (Леонова и др., 2002). Вторая находка более крупного экземпляра немного лучшей сохранности, сделанная несколькими годами позже из того же местонахождения, подтвердила присутствие Neouddenites в роудском комплексе аммоноидей этого региона (Barskov et al., 2014). Таким образом, время существования Neouddenites распространилось за рамки кунгура и включило в себя роуд. Морфология этого рода определенно указывает на его родство с Uddenites и Uddenoceras, но существование огромного временного хиатуса в три века между предковой формой (гжель) и “потомком” Neouddenites (кунгур-роуд) по-прежнему не находит объяснения. Остается только ожидать, что будут найдены какие-то родственные формы в этом временном интервале.
Основной филогенетический ряд в подсемействе Medlicottiinae, в состав которого входят роды с наиболее сложными лопастными линиями, принимается практически во всех классификациях: Artinskia → Medlicottia → Eumedlicottia → → Syrdenites. Основным звеном развития является усложнение наружного седла лопастной линии, а также увеличение числа и усложнение лопастей, расположенных на боковой стороне раковины.
Самая древняя Artinskia Karpinsky, 1926 – A. irinae Ruzh. – известна из верхнего карбона (оренбургский ярус В.Е. Руженцева) Южного Урала (Руженцев, 1950). Автор связывал ее происхождение с родом Uddenoceras. Это предположение представляется вполне логичным. О связи указанных родов свидетельствует сильно уплощенная форма раковины Uddenoceras, а также скульптура одного из видов – U. harlani (Plummer et Scott), на вентральной стороне которого наблюдаются выпуклые округлые бугорки, характерные и для Artinskia, и строение лопастной линии с высоко поднятой наружной частью первой латеральной лопасти (рис. 1). Единичные представители рода Artinskia были распространены в раннепермских бассейнах разных биогеографических областей. С территории Американского континента достоверно известны находки A. lilianae Miller et Yuongquist (формации Bursum, Pueblo и Wolfcamp Техаса, соответствующие ассельскому и сакмарскому ярусам). В Тетической области всего два вида Artinskia найдены в сакмарских и кунгурских отложениях – A. separata Leonova (Памир, хориджская свита) (Левен и др., 1992) и A. timorensis Haniel (Тимор, слои Bitauni) (Haniel, 1915). Наиболее полно этот род представлен на Урале, где он был впервые установлен (Karpinsky, 1926) с типовым видом A. artiensis (Grunewaldt, 1860). Все остальные виды уральских Artinskia были выделены и изучены Руженцевым (1950, 1951, 1956), который построил детальный филогенетический ряд: A. irinae Ruzhencev (поздняя гжель) → A. kazakhstanica Ruzhencev (основание ассели) → A. nalivkini Ruzhencev (ассель-сакмара) → A. multituberculata Ruzhencev (сакмара) → → A. artiensis (арти). Следует отметить, что очертания вершины наружного седла в лопастной линии не претерпели значительных изменений в течение всего времени существования рода. В процессе эволюции увеличивалось число лопастей на боковой стороне, уменьшалась глубина первой боковой лопасти, увеличивались размеры зубцов в основании лопастей, но в вершине наружного седла сохранялись всего две пары адвентивных лопастей.
Рис. 1.
Uddenoceras harlani (Plummer et Scott, 1937): a, б – раковина (×1), в – лопастная линия (×3) (по: Plummer, Scott, 1937, с.70, табл. 3 , фиг.15, 16, 18 ) и Artinskia irinae Ruzhencev, 1938: г, д – раковина (×1), е – лопастная линия (×2) (по: Руженцев, 1950, табл. 3 , фиг. 3 b, c, рис. 27 а).

Приобретение большего числа адвентивных лопастей знаменует переход к новому роду Medlicottia Waagen, 1880. Medlicottia значительно превосходит Artinskia по сложности строения наружного седла: число парных адвентивных лопастей в его вершине увеличивается до трех-шести, в зависимости от геологического возраста вида. До настоящего времени считалось, что Medlicottia появилась в конце сакмарского века. В течение сакмарского и артинского веков оба рода развивались параллельно. В артинском ярусе Урала многочисленные Artinskia artiensis найдены в одних и тех разрезах, что и M. orbignyana (Verneuil, 1845) (рис. 2).
Филогенетический ряд уральских Medlicottia был выстроен Руженцевым (1960), который считал, что род возник в конце сакмарского века: M. semota Ruzhencev (поздняя сакмара) → M. vetusta Ruzhencev (поздняя сакмара) → M. intermedia Ruzhencev (раннее арти) → M. orbignyana (позднее арти) → M. postorbignyana Bogoslovskaya (кунгур-роуд). Последний вид М.Ф. Богословская (1997) описала по материалам с Полярного Урала (Пай-Хой), а затем он был обнаружен в роудских (казанских) отложениях Волго-Урала (Леонова и др., 2002; Leonova, 2007; Barskov et al., 2014) и Арктической Канады (Леонова, 2006; Leonova, 2007). В этом ряду морфогенез шел по пути постоянного усложнения лопастной линии в зоне вершины наружного седла (увеличение числа адвентивных лопастей), сужения оборотов и вентральной стороны, а также исчезновения косо-направленных ребрышек и бугорков на наружной части боковых и вентральной стороны. На узкой вентральной стороне образовалось два острых киля.
В последние годы (2015–2017 гг.) А.В. Мазаевым и М.С. Бойко было найдено несколько экземпляров Medlicottia в более древних отложениях раннепермского рифа Шах-Тау, которые датируются позднеассельским-раннесакмарским временем. Это почти на целый век опускает нижнюю границу распространения рода. Вид M. subdorbignyi из шихана Тра-Тау, расположенного недалеко от Шах-Тау, был описан Н.П. Герасимовым по единственному образцу – обломку раковины с сохранившимся небольшим фрагментом лопастной линии (Герасимов, 1937, с. 18, табл. 1, фиг. 7). Руженцев (1951), не располагая дополнительным материалом, отнес вид Герасимова к роду Artinskia.
Medlicottia subdorbignyi Gerassimov – самая древняя из найденных на сегодняшний день медликоттий – отличается относительно широкой вентральной стороной с двумя рядами ярко выраженных ребрышек-бугорков, косо расположенных по периферии боковых сторон (рис. 3, a). Этот признак приближает ее к предковому роду Artinskia. Лопастная линия с тремя парами адвентивных лопастей на наружном седле и одной непарной, двузубчатой (рис. 3, б). При высоте оборота 38 мм на боковой стороне расположено 12 лопастей, семь из них с двузубчатым основанием. В целом она очень близка лопастным линиям сакмарских M. semota и M. vetusta при соответствующих размерах раковины. Герасимов (1937) при описании этого вида указывал, что лопастная линия нового вида при высоте оборота 50 мм отвечает таковой M. orbignyana при высоте оборота в 15 мм. В действительности это утверждение справедливо лишь для наружного седла, на боковой стороне взрослой раковины M. subdorbignyi расположено 12 лопастей, а у юной M. orbignyana – только девять. Новый материал еще более надежно подтверждает связи между Artinskia и Medlicottia как предка и потомка.
Рис. 3.
Medlicottia subdorbignyi Gerassimov: a – раковина (×1), б – фрагмент лопастной линии (×3). Обр. 5615/2; шихан Шах-Тау; верхнеассельский–нижнесакмарский подъярус.

Существует еще два региона, где было найдено несколько видов Medlicottia. Это Памир и запад США. Памирские медликоттии представлены четырьмя видами (М. basarensis Toumanskaya, M. tenuis Leonova, M. busterensis Toumanskaya, M. jakovlevi Toumanskaya), все они собраны примерно с одного стратиграфического уровня, из болорских отложений, и демонстрируют морфологическое разнообразие раковины и лопастной линии, отличаясь шириной раковины, числом лопастей на боковой стороне и тонкими особенностями скульптуры. В отношении валидности двух последних видов существуют некоторые сомнения, поскольку они были неполно описаны по единичным образцам плохой сохранности. В наших коллекциях, представленных гораздо более богатым материалом, представителей этих видов установить не удалось.
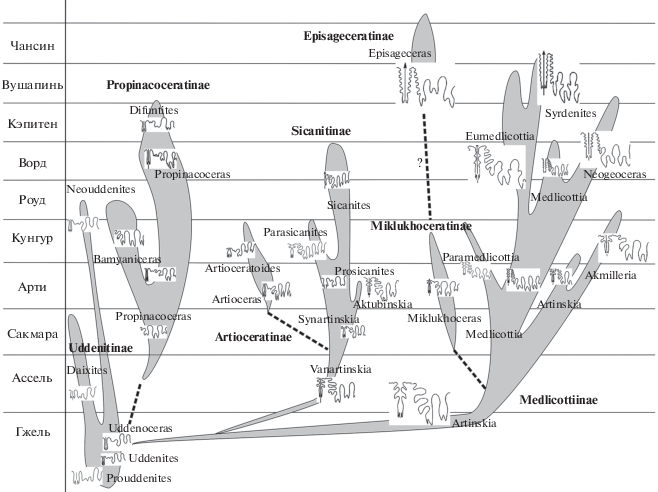
Все американские медликоттии происходят из перми Техаса и представлены следующими видами: M. copei White, 1891 (нижний арти – формация Clyde) → M. arroensis Plummer et Scott, 1937 (арти – формация Arroyo) → M. chozaensis Plummer et Scott, 1937 и M. milleri Plummer et Scott, 1937 (кунгур – формация Choza); M. costellifera Miller et Furnish, 1940 (кунгур – формации Leonard и Bone Spring) → M. kingorum Miller et Furnish, 1940 (ворд – формация Word). Их морфологические особенности вполне согласуются с той стратиграфической последовательностью, в которой были сделаны находки. Изменения очертаний их лопастных линий в процессе морфогенеза показаны на рис. 4.
Рис. 4.
Филогенетический ряд американских медликоттий: a – M. copei White, 1891 (нижний арти, формация Clyde), Д = 35 мм; б – M. arroensis Plummer et Scott, 1937 (арти, формация Arroyo), Д = 33 мм; в – M. chozaensis Plummer et Scott, 1937, Д = 41 мм; г – M. milleri Plummer et Scott, 1937 (кунгур, формация Choza), Д = 35 мм (по: Plummer, Scott, 1937, табл. 5 , фиг. 3, 8 , табл. 6 , фиг. 3, 6); д – M. costellifera Miller et Furnish, 1940 (кунгур, формации Leonard и Bone Spring), Д = 72 мм; е – M. kingorum Miller et Furnish, 1940 (ворд, формация Word), Д = 36 мм (по: Miller, Furnish, 1940, с. 61, рис. 15b, c ).
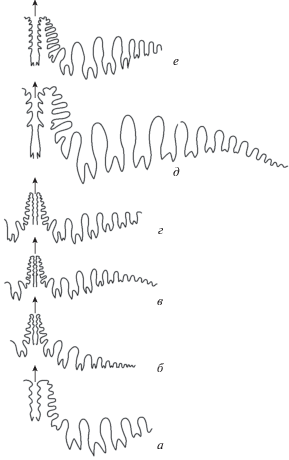
В отношении двух-трех боковых ответвлений от основного ствола существуют разногласия. В ревизованном издании “Treatise …” (Furnish et al., 2009), кроме указанных выше четырех родов основного ряда, в состав подсемейства введен только род Neogeoceras. Род Paramedlicottia сведен в синонимику рода Medlicottia. По-видимому, непризнание рода связано с тем, что материал по нему очень ограничен, сохранность его не слишком хорошая, и описание дано только на русском языке (Левен и др., 1992). Поэтому здесь приведены дополнительные пояснения, чем отличаются роды Paramedlicottia и Medlicottia. Они четко различны по двум группам признаков, по форме раковины и по очертаниям лопастной линии. Medlicottia имеет совершенно инволютную раковину, начиная с третьего оборота, с узким диаметром умбилика (Ду/Д = 0.05–0.07; Руженцев, 1956). В отличие от нее, взрослая Paramedlicottia (6–7 оборотов) обладает умеренно инволютной раковиной со значительно более широким умбиликом (соотношение Ду/Д = 0.19–0.20) (рис. 5, г–е). Лопастная линия памирского рода характеризуется более простым расчленением наружного седла (рис. 5, ж), которое несет всего две-три пары адвентивных лопастей. Но при этом общее число лопастей на боковой стороне (до умбиликального шва) больше, чем у одновозрастной M. orbignyana – 12 против девяти при сходных размерах раковины (рис. 5, ж, и). При этом у обоих родов двузубчатых лопастей на боковой стороне равное число – по шесть. Paramedlicottia напоминает Artinskia небольшим числом адвентивных лопастей в вершине наружного седла и умеренно инволютной раковиной, характерной для юных артинский (рис. 5, б), но отличается отсутствием выпуклых бугорков на вентральной стороне (рис. 5, a), у Paramedlicottia на ней расположены два гладких киля. Лопастная линия у Artinskia при сопоставимых размерах имеет более простое строение: в наружном седле имеется только одна пара адвентивных лопастей, а на боковой стороне расположено всего четыре двузубчатых лопасти, причем зубцы значительно слабее развиты. В отличие от Paramedlicottia, боковые лопасти Artinskia более узкие и глубокие, слабо пережатые в средней части (рис. 5, в). Описанные особенности позволяют достаточно уверенно предполагать филогенетическую связь между Artinskia → Paramedlicottia.
Рис. 5.
Представители родов Artinskia, Paramedlicottia и Medlicottia: a–в – A. artiensis Grünwaldt, раковины молодого (×1) и юного (×1.5) экземпляров и лопастная линия (×5) (по Руженцев, 1956, табл. 3 , фиг. 3, 4, рис. 20 m ); г–ж – P. sauksayensis Leonova: г и е – раковины (× 1.5), д – (×1), ж – лопастная линия (×5) (по Левен и др., 1992, табл. 30 , фиг. 8–10 , рис. 29 в); з, и – M. orbignyana Verneuil: з – раковина (×1) (по Miller et Furnish, 1940, табл. 10 , фиг. 1, 2), и – лопастная линия (×5) (по Руженцев, 1956, табл. 5 , фиг.2, рис. 27p ).
Подсемейство Sicanitinae Noetling, 1904 – одно из наиболее часто упоминаемых в работах по медликоттиидам. Как было показано выше, состав его у разных авторов сильно варьирует (табл. 1). Как и в отношении всех других медликоттиид, основной массив данных по этой группе собран на Урале и Памире. Номинативный род Sicanites Gemmellaro был описан из типового местонахождения на о. Сицилия; кроме этого известны находки из Афганистана, Омана и Памира. Неоправданное расширение набора признаков этого рода и отнесение к нему целого ряда таксонов родового ранга (Vanartiskia Ruzhencev, Aktubinskia Ruzhencev, Prosicanites Toumanskaya и даже Medlicottia Waagen – вида M. costellifera) противоречит диагнозу как самого рода Sicanites, так и всех сводимых в синонимию родов. При установлении рода Дж. Джеммелларо указал его диагностические признаки и сравнил с другими медликоттиидами (Gemmellaro, 1887, с. 62 –64). В частности, автор отметил “дисковидную раковину … с широким пупком и с узкой и округлой вентральной стороной, которая на … последнем обороте украшена поперечными бугорками, которые располагаются по вентро-латеральному перегибу и образуют по средней линии продольную канавку. На оборотах имеются глубокие пережимы, напоминающие контур зонтика (ombellicale)… Сифонная лопасть узкая, глубокая … Наружное седло имеет раздвоенную вершину и одно (курсив мой – Т.Л.) адвентивное седло на внешней стороне”. При сравнении с Medlicottia Джеммелларо (Gemmellaro, 1887, с. 63) указывает, что новому роду “не хватает характерного строения седла с адвентивными лопастями, как у Medlicottia, а также он имеет обороты с пережимами, которых у медликоттий нет”.
Возможно, из-за нечеткого представления о характерных чертах рода Sicanites в ревизованном издании “Treatise …” (Furnish et al., 2009) в этот таксон включено несколько совершенно разных родов, объединенных лишь одним признаком – глубоким положением первой боковой лопасти. Нужно отметить, что этот признак наблюдается во многих группах медликоттиид и часто зависит от эволюционного возраста таксона. Первые представители Artinskia, а также Medlicottia на ранних онтогенетических стадиях обладали этим признаком. По непонятной причине в справочнике не приведено лопастной линии типового вида S. shopeni, а даны линии самых разных родов, никоим образом не соответствующих диагнозу рода Sicanites ни по форме раковины, ни по скульптуре, ни по очертанию лопастной линии.
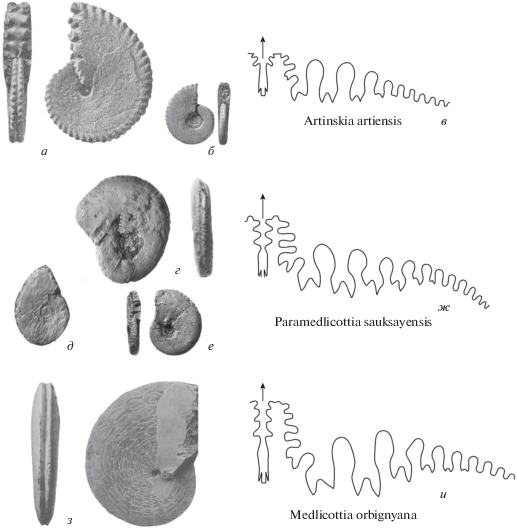
Несмотря на то, что описание рода Parasicanites публиковалось дважды (Леонова, 1985; Леонова, Дмитриев, 1989), по-видимому, основные отличия его от рода Synartinskia остались неясными. На рис. 6 показаны некоторые онтогенетические стадии лопастных линий обоих родов при близких размерах раковины. Нетрудно заметить, что наблюдается много различий в двух, казалось бы, очень похожих линиях. И что еще более примечательно, их онтогенетическое развитие проходит по-разному: при В = 6–7 мм линии очень близки (рис. 6, в, и), но уже при В = 10 мм у Synartinskia появляется хорошо сформированная адвентивная лопасть на наружной стороне наружного седла (рис. 6, к), а у Parasicanites в этой части нет никаких образований (рис. 6, г). При этом общее число лопастей на боковой стороне Parasicanites равно 12, а у Synartinskia 10. При В = 17–20 мм у обоих родов имеется хорошо сформированная наружная адвентивная лопасть, но при этом у Parasicanites (В = 17 мм) хорошо развита и внутренняя адвентивная лопасть, парная с наружной (рис. 6, д), а у Synartinskia (В = 20.5 мм) эта лопасть остается плохо сформированной (рис. 6, л). Кроме этого, на боковой стороне Parasicanites расположено 14 лопастей, а Synartinskia – 11. При В = 25 мм у Parasicanites непарная крупная внутренняя адвентивная лопасть становится многозубчатой, парная внутренняя лопасть – глубокая и широкая, самое высокое – третье боковое седло (рис. 6, е), а у Synartinskia непарная лопасть остается двузубчатой, парная внутренняя адвентивная лопасть плохо сформирована, а самым высоким является второе боковое седло (рис. 6, м).
Рис. 6.
Parasicanites meridionalis Leonova: a – раковина (×1), б – скульптура в области вентро-латерального перегиба (×1.5), в–е – лопастная линия (в – при В = 7.3 мм (×4), г – при В = 10.4 мм (×4), д - при В = 17 мм (×3), е – при В = 25 мм (×3) (по Леонова, 1985, рис. 5); и Synartinskia principalis Ruzhencev: ж – раковина (×1), з –скульптура в области вентро-латерального перегиба (×0.75) (по Руженцев, 1951, табл. 3 , фиг. 9 , табл. 4 , фиг. 2), и–м – лопастная линия (и – при В = 6 мм (×4), к – при В = 9.8 мм (×4) (по Леонова, Воронов, 1989, рис. 2, m, n), л – при В = 20.5 мм (×3), м – при В = = 25 мм (×2.5) (по Руженцев, 1951, рис. 17 d, f )).
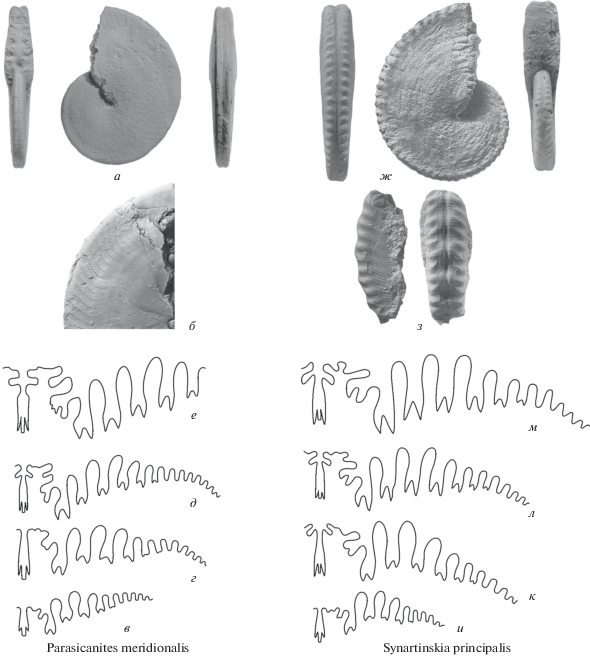
Рис. 7.
Представители родов Miklukhoceras (a–к) и Episageceras (л–p): a, б – M. pressulum Leonova (a – раковина (×1), б – лопастная линия (×2)); в–е – M. pamiricum Pavlov (лопастные линии: в – (×2), е – ×3; г, д – раковины (×1) (по Леонова, 1984, табл. 4 , рис. 5); ж, з – M. artumbilicatum Leonova (ж – (×1), з – лопастная линия (×4)); и, к – M. praevium Leonova (и – раковина (×1), к – лопастная линия (×6)) (по Левен и др., 1992, табл. 29 , рис. 21 ); л–p – E. noetlingi Haniel, раковины – л (по Haniel, 1915, табл. 10 ), н, п (×1); лопастные линии: о (×4), p (×6) (по Leonova, 2002, fig. 17 ), м (по Furnish et al., 2009, fig. 137n ).

Наиболее ярко отличия проявляются в скульптуре и форме раковины. В то время как вентральная сторона Synartinskia украшена крупными сильно выпуклыми бугорками, переходящими на вентро-латеральный перегиб (рис. 6, ж, з), вентральная сторона Parasicanites несет два киля, поверхность которых почти гладкая (рис. 6, a, б).
Подсемейства Miklukhoceratinae и Episageceratinae
Включение Episageceratinae в качестве подсемейства в состав медликоттиид (Furnish et al., 2009) не вызывает особых возражений. На фоне огромного морфологического разнообразия лопастных линий Medlicottiidae эписагецератины вполне могут рассматриваться в составе этого таксона. Больше сомнений ранее вызывала форма раковины. До находок представителей рода Miklukhoceras (рис. 7, a, г, д, ж, и) казалось совершенно необъяснимым появление среди медликоттиид вздутой эволютной раковины с грубой поперечной скульптурой, которая наблюдается на ранних онтогенетических стадиях Episageceras (рис. 7, н, п). Находки в сакмарско-кунгурских отложениях Тетической области нескольких представителей рода Miklukhoceras позволяют предположить зарождение направления с таким особым типом развития раковины внутри семейства медликоттииид на стадии расцвета этой группы, ее максимального таксономического разнообразия. Как показывают морфологический и количественный анализ эволюционных изменений медликоттиид (Богословская и др., 1996), сакмарско-вордский период для этой группы был наиболее продуктивным. Самый древний Miklukhoceras был обнаружен в хориджской свите Дарваза, коррелируемой с сакмарским ярусом (Левен и др., 1992). Еще несколько форм были найдены в артинском и кунгурском ярусах Памира (Леонова, 1984; Леонова, Дмитриев, 1989; Левен и др., 1992). Кроме этого, представители Miklukhoceras позднеартинско-кунгурского возраста указываются из известняка Ratburi Таиланда (Glenister et al., 1990) и формации Longyin Южного Китая (Zhou, 2017). Относительно валидности рода Kunlunoceras (Wang, 1983) из Кунь-Луня возникают большие сомнения, поскольку описанный образец имеет плохую сохранность, и изображение его в опубликованной работе не дает достаточно информации. Наиболее вероятно, “Kunlunoceras” является юным представителем Miklukhoceras, возможно, новым видом. Утверждать что-то более определенное можно будет лишь после получения более качественных данных.
В настоящее время можно уверенно утверждать, что в сакмарском веке в небольшой группе медликоттиид появилась новая тенденция, которая выразилась в достаточно долгом сохранении в онтогенезе вздутой эволютной раковины (первые четыре-пять оборотов до диаметра раковины 15 мм). Для других медликоттиид такая форма была характерна только для самых ранних стадий, не далее первых двух оборотов. Второй особенностью этой группы является возникновение грубой скульптуры в виде резких ребрышек на боковых и вентральной сторонах, что совсем не типично для всего отряда Prolecanitida.
Большую проблему для установления каких-либо связей между Miklukhoceratinae и Episageceratinae представляет строение их лопастных линий. У Miklukhoceras она довольно простая с одной наружной адвентивной лопастью, что сближает ее с лопастной линией рода Akmilleria. Вторая боковая лопасть самая крупная (рис. 7, б, в, е, к). У Episageceras, наоборот, уже на достаточно ранних стадиях вторая боковая лопасть приобретает характер вспомогательной, становится очень мелкой (рис. 7, о, p). У взрослых форм наружное седло имеет очень сложное строение с восемью парами адвентивных лопастей (рис. 7, м), что несколько напоминает наружное седло рода Syrdenites, который, в свою очередь, отличается очень мелкой первой боковой лопастью.
Определяется ли эта общность признаков внешней морфологии раковины параллелизмами, связанными с общностью происхождения отдаленно родственных групп? Собранный к настоящему времени материал является совершенно недостаточным для решения филогенетических вопросов, но заставляет задуматься о них.
По тем же причинам пока нельзя однозначно определить в системе подотряда Medlicottiina положение монотаксонных семейств Sundaitidae (Руженцев, 1949), Shikhanitidae (Руженцев, 1949) и Darvasiceratidae (Леонова, 1990).
Список литературы
Богословская М.Ф. Аммоноидеи из пермских отложений Пай-Хоя и острова Вайгач // Палеонтол. журн. 1997. № 6. С. 23–28.
Богословская М.Ф., Леонова Т.Б., Наймарк Е.Б. К вопросу о таксономической иерархии пролеканитов (Ammonoidea) // Палеонтол. журн. 1996. № 3. С. 9–14.
Герасимов Н.П. Уральский отдел пермской системы // Уч. зап. Казан. гос. ун-та. 1937. Т. 97. Кн. 3–4. Геология. Вып. 8–9. С. 3–68.
Захаров Ю.Д. Рост и развитие аммоноидей и некоторые проблемы экологии и эволюции // Систематика и экология головоногих моллюсков / Ред. Старобогатов Я.И., Несис К.Н. Л.: ЗИН АН СССР, 1983. С. 26–31.
Левен Э.Я., Леонова Т.Б., Дмитриев В.Ю. Пермь Дарваз-Заалайской зоны Памира: фузулиниды, аммоноидеи, стратиграфия. М.: Наука, 1992. 203 с. (Тр. ПИН РАН. Т.253).
Леонова Т.Б. Роды Bamyaniceras и Miklukhoceras (Ammonoidea) на Памире // Палеонтол. журн. 1984. № 2. С. 40–50.
Леонова Т.Б. Некоторые медликоттииды (Ammonoidea) Памира // Ископаемые головоногие моллюски: Основные направления изучения. М.: Наука, 1985. С. 70–82.
Леонова Т.Б. Darvasiceratidae – новое семейство пермских пролеканитид // Палеонтол. журн. 1990. № 3. С. 104–108.
Леонова Т.Б. Онтофилогенетические исследования палеозойских аммоноидей // Изв. РАН. Сер. биол. 2012. № 2. С. 237–248.
Леонова Т.Б., Воронов А.В. Онтогенетическое развитие лопастной линии раннепермского рода аммоноидей Synartinskia // Палеонтол. журн. 1989. № 1. С. 113–116.
Леонова Т.Б., Дмитриев В.Ю. Раннепермские аммоноидеи Юго-Восточного Памира // Тр. ПИН АН СССР. 1989. Т. 235. 198 с.
Леонова Т.Б., Есаулова Н.К., Шиловский О.П. Первая находка казанских аммоноидей в Волго-Уральском регионе // Докл. АН. 2002. Т. 383. № 4. С. 509–511.
Руженцев В.Е. Систематика и эволюция семейств Pronoritidae Frech и Medlicottiidae Karpinsky // Тр. ПИН АН СССР. 1949. Т. 19. 206 с.
Руженцев В.Е. Верхнекаменноугольные аммониты Урала // Тр. ПИН АН СССР. 1950. Т. 29. 220 с.
Руженцев В.Е. Нижнепермские аммониты Южного Урала. 1. Аммониты сакмарского яруса // Тр. ПИН АН СССР. 1951. Т. 33. 188 с.
Руженцев В.Е. Нижнепермские аммониты Урала: II. Аммониты артинского яруса // Тр. ПИН АН СССР. 1956. Т. 60. 271 с.
Руженцев В.Е. Принципы систематики, система и филогения палеозойских аммоноидей // Тр. ПИН АН СССР. 1960. Т. 33. 331 с.
Руженцев В.Е. Ассельские аммоноидеи на Памире // Палеонтол. журн. 1978. № 1. С. 36–52.
Руженцев В.Е., Богословский Б.И., Либрович Л.С. Надотряд Ammonoidea // Основы палеонтологии. Моллюски–головоногие / Ред. Руженцев В.Е. М.: Изд-во АН СССР, 1962. С. 243–428.
Barskov I.S., Boiko M.S., Konovalova V.A. et al. Cephalopods in the marine ecosystems of the Paleozoic // Paleontol. J. 2008. V. 42. № 11. P. 1167–1284.
Barskov I.S., Leonova T.B., Shilovsky O.P. Middle Permian cephalopods of the Volga-Ural region // Paleontol. J. 2014. V. 48. № 13. P. 1331–1414.
Furnish W.M., Glenister B.F., Kullmann J., Zhou Z. Treatise on Invertebrate Paleontology. Pt. L. Mollusca 4. Revised, V. 2: Carboniferous and Permian Ammonoidea (Goniatitida and Prolecanitida) / Ed. Seldon P.A. Lawrence: Geol. Soc. Amer. and Univ. Kansas Press, 2009. 258 p.
Gemmellaro G.G. La fauna dei calcari con Fusulina della valle del fiume Sosio nella provincia di Palermo // Giorn. Sci. Natur. Econ. 1887. V. 19. 106 p.
Glenister B.F., Furnish W.M., Zhou Z., Malai P. Ammonoid cephalopods from the Lower Permian of Thailand // J. Paleontol. 1990. V. 64. № 3. P. 479–480.
Haniel C.A. Die Cephalopoden der Dyas von Timor // Paläontol. von Timor. 1915. Lfg. 3. Abh. 6. P. 1–153.
Karpinsky A. On a new species of Ammonoid of the family Medlicottiidae; on the relationship between genera of this family, and on the ontogeny and phylogeny of Prolecanitidae // Ann. Soc. Paléontol. Russ. 1926. V. 4. P. 1–19.
Leonova T.B. Permian ammonoids: classification and phylogeny // Paleontol. J. 2002. V. 36. Suppl. 1. P. 1–114.
Leonova T.B. Correlation of the Kazanian of the Volga-Urals with the Roadian of the global Permian scale // Paleoworld. 2007. V. 16. P. 246–253.
Miller A.K., Furnish W.M., Schindewolf O.H. Paleozoic Ammonoidea // Treatise on Invertebrate Paleontology. Pt. L. Mollusca 4. Cephalopoda. Ammonoidea. Lawrence: Univ. Kansas Press, 1957. P. 11–81.
Plummer F.B., Scott G. Upper Paleozoic ammonites in Texas // Geology of Texas. Bull. Texas Univ. 1937. V. 3. № 3701. 516 p.
Wang M. Ammonoidea // Paleontological Atlas of Northwest China, Xinjiang volume. Pt II. Beijing: Geol. Publ. House, 1983. P. 510–533.
Zhou Z. Permian basinal ammonoid sequence in Nanpanjiang area of South China – possible overlap between basinal Guadalupian and platform-based Lopingian // J. Paleontol. 2017. V. 91. Mem. 74. P. 1–95.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал