

Палеонтологический журнал, 2019, № 4, стр. 87-92
Новый пермский тероцефал (Therocephalia, Theromorpha) из сундырского комплекса Восточной Европы
Ю. А. Сучкова a, *, В. К. Голубев a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
* E-mail: js@paleo.ru
** E-mail: vg@paleo.ru
Поступила в редакцию 04.12.2018
После доработки 11.12.2018
Принята к публикации 28.12.2018
Аннотация
По материалам из типового местонахождения сундырского комплекса тетрапод Сундырь-1 (Республика Марий Эл, Россия; средняя пермь) описан новый тероцефал сем. Lycosuchidae – Gorynychus sundyrensis sp. nov. В сундырском сообществе тетрапод G. sundyrensis занимал нишу крупного падалеяда.
ВВЕДЕНИЕ
Завершающий этап эволюции диноцефаловой фауны тетрапод на территории Восточной Европы представлен сундырским комплексом (Голубев и др., 2015; Golubev, 2015; Сенников, Голубев, 2017). В более древних диноцефаловых комплексах: мезенском, голюшерминском, очёрском, ишеевском экологическую нишу крупного хищника в тетраподных сообществах занимали примитивные горгонопии и диноцефалы (Голубев, 2000; Ивахненко, 2001, 2015; Ivakhnenko, 2003). В сундырском комплексе впервые крупнейшими хищниками стали тероцефалы. В работе Ю.А. Сучковой и В.К. Голубева (2019) на материале из местонахождения Сундырь-1 был описан самый крупный тероцефал сундырской фауны – Julognathus crudelis Sutschkova et Golubev. Наряду с многочисленными остатками Julognathus в этом же местонахождении обнаружены более редкие и чаще уступающие по размерам кости другого примитивного тероцефала. Второй тероцефал отличается от юлогнатуса (1) значительно более короткой преорбитальной частью черепа, (2) меньшим количеством верхних резцов и верхних заклыковых зубов, (3) отсутствием характерного для примитивных тероцефалов вытянутого и отнесенного далеко назад подбородочного выступа и (4) наличием грубой зазубренности на режущих кантах всех зубов (5–10 зазубрин на 5 мм в отличие от 15–20 зазубрин на 5 мм у юлогнатуса). Последние два признака среди примитивных тероцефалов известны только у Gorynychus masyutinae Kammerer et Masyutin, который описан по материалам из местонахождения Котельнич в Кировской области (Kammerer, Masyutin, 2018). С G. masyutinae нового сундырского тероцефала также сближает наличие отчетливо выраженного преорбитального гребня на предлобных костях и характерной ямчатой скульптуры на внешней поверхности челюстных и носовых костей. В то же время, от котельничского тероцефала сундырская форма отличается морфологией зубной системы – строением зубов и их числом. Все вышеперечисленное позволяет рассматривать сундырского тероцефала в качестве нового вида рода Gorynychus – G. sundyrensis sp. nov. Род Gorynychus отнесен нами к семейству Lycosuchidae на основании таких признаков, как короткая и широкая предглазничная часть черепа, наличие менее пяти верхних заклыковых зубов и не более пяти верхних резцов (van den Heever, 1987, 1994; Abdala et al., 2008, 2014).
МАТЕРИАЛ
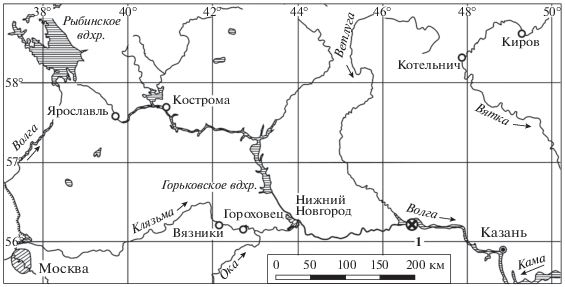
Материал происходит из местонахождения Сундырь-1. Местонахождение расположено на правом берегу р. Волга (южный берег Чебоксарского водохранилища) непосредственно ниже устья р. Сундырь, в 2.5 км северо-западнее д. Токари и в 1.6 км восток-северо-восточнее с. Юлъялы, в Горномарийском р-не Республики Марий Эл (рис. 1). Описание разреза и местонахождения приведено в работах Голубева (Голубев и др., 2015; Голубев, Буланов, 2018). Стратиграфическое положение костеносных слоев – средняя пермь, нижняя часть верхнесеверодвинского подъяруса (=кептенский ярус), нижний подгоризонт путятинского горизонта. Изученная коллекция ископаемых остатков хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (Москва), № 5388.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД THEROCEPHALIA
СЕМЕЙСТВО LYCOSUCHIDAE NOPSCA, 1923
Род Gorynychus Kammerer et Masyutin, 2018
Gorynychus sundyrensis Sutschkova et Golubev, sp. nov.
Табл. VIII , фиг. 1–5 ; табл. IX, фиг. 1–6 (см. вклейку)
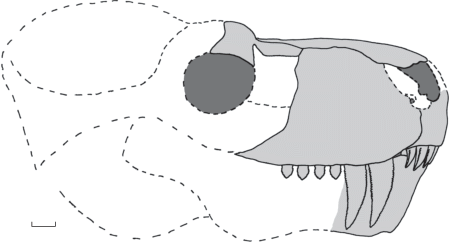
Рис. 2.
Gorynychus sundyrensis sp. nov., череп сбоку, реконструкция по голотипу ПИН, № 5388/327 и экз. ПИН, №№ 5388/55, 109, 213, 340, 364, 436. Длина масштабной линейки 2 см.
Название вида по типовому местонахождению Сундырь-1.
Голотип – ПИН, № 5388/327, правое maxillare; Республика Марий Эл, Горномарийский р-н, местонахождение Сундырь-1; средняя пермь, верхнесеверодвинский подъярус, нижнепутятинский подгоризонт.
Описание (рис. 2). Средние и крупные тероцефалы с короткой и высокой предглазничной частью черепа, массивной нижней челюстью и грубозазубренными челюстными зубами. Реконструируемая длина черепа у взрослых особей составляла не менее 35–40 см. Зубная формула: I 4/3; C 1-2/1; Pc 4/?.
Челюстная кость (табл. VIII , фиг. 1а, б) сравнительно короткая и высокая, с коротким скуловым отростком. Большая часть латеральной поверхности кости покрыта скульптурой из многочисленных тесно расположенных округлых ямок с приподнятыми, часто общими краями. Местами между ямками располагаются небольшие отверстия, от которых по поверхности кости следуют длинные и разветвленные желобки. Над альвеолярным краем поверхность кости гладкая, с мелкими отверстиями и отходящими от них желобками. В задней части на processus jugalis развит неглубокий желоб, выходящий на вентральную сторону и дающий небольшое латеральное расширение альвеолярного края кости с уровня четвертого заклыкового зуба. Это расширение несколько напоминает максиллярный выступ, встречающийся у южноафриканских ликозухид (van den Heever, 1994), но очень слабо выраженный. На более крупном экз. ПИН, № 5388/59 максиллярный выступ развит еще слабее.
Нижняя стенка максиллярного синуса практически сразу от его начала образует заметный наклон в постеро-медиальном направлении. Поверхность контакта с небной костью имеет субтреугольную форму: она высокая в начале, у альвеолы клыка, и резко уменьшается по направлению к processus jugalis. На постеро-медиальной части стенки альвеолы клыка контактная поверхность покрыта гребешками, образующими хорошо выраженный “древовидный” узор. По остальной ее части распространены “лесенкой” более грубые субвертикальные контактные гребни (табл. VIII , фиг. 1б). Медиальная стенка альвеолы клыка в нижней части повреждена, край хоаны здесь не сохранился. Позади клыка вдоль края хоаны располагается небольшой валик, но crista choanalis не развит. Зубной ряд вместе с альвеолой клыка слабо выгнут наружу, клык по вентрально-латеральному краю почти не выделяется. Заклыковых зубов четыре, что отличает данного тероцефала от Gorynychus masyutinae, у которого их три.
Альвеола клыка экз. ПИН, № 5388/327 содержит два корня обломанных зубов. Судя по их размеру, оба клыка были примерно одной величины. У экз. ПИН, № 5388/59 сохранилась только латеральная стенка альвеолы клыка, обломанная спереди, но по пропорциям относительно длины зубного ряда она также могла вмещать два соразмерных зуба. Очевидно, у Gorynychus sundyrensis смена клыков проходила следующим образом: сначала вырастал новый клык, затем, спустя некоторое время выпадал старый. Таким образом, время от времени в верхней челюсти с каждой стороны головы располагалось по два клыка. Подобное явление широко распространено среди южноафриканских ранних тероцефалов (van den Heever, 1980). Например, среди обнаруженных челюстных костей ликозухид почти 40% с двумя клыками (Abdala et al., 2014).
Предчелюстная кость низкая и сравнительно короткая (табл. VIII , фиг. 3 а, б). Задний конец кости за альвеолой последнего резца, в области захождения нижнечелюстного клыка (fossa dentia canini inferioris) сужается и латерально уплощается. Вертикальный отросток тонкий, имеет узко-треугольную форму в сечении. Симфиз покрыт неглубокими гребешками в передней части и более выраженными сзади. Латеральная поверхность покрыта хорошо выраженной скульптурой в виде ямок с отходящими от них желобками (табл. VIII , фиг. 3 б). Область налегания премаксиллярного отростка maxillare имеет мелкопористую структуру, она гладкая, за исключением прохождения в верхней части двух небольших извилистых желобков и одного крупного под ними, входящего в толщу кости над третьим резцом на границе контактной и внешней поверхностей. Под основанием spina nasalis приблизительно посередине высоты кости развито крупное сосудистое отверстие.
На дорсальной поверхности кости также в основании spina nasalis присутствует вытянутая ямка с сосудистыми отверстиями по переднему и заднему краям. Одно из них, переднее, связано каналом с вышеописанным отверстием на латеральной поверхности под основанием spina nasalis, а второе – с foramen incisivum ossis praemaxillaris на вентральной стороне. Большая часть дорсальной поверхности кости занята площадкой налегания септомаксиллы. Площадка округло-ромбической формы, покрыта мелкими контактными гребешками, с гладким бугорком посередине. Сразу за ней на сужении кости в задней части располагается контактная площадка с челюстной костью.
Небная пластина предчелюстной кости короткая и широкая, с сосудистым отверстием (foramen incisivum ossis praemaxillaris), расположенным в районе альвеол первого и второго резцов (табл. VIII , фиг. 3 а). Приблизительно в середине пластины она неглубоко вырезана. Со стороны интерпремаксиллярного симфиза от вырезки и большей частью на дорсальной стороне премаксиллы виден след вхождения сошника. Противоположный край вырезки имеет вид овальной площадки, отгибающейся внутрь от небной пластины praemaxillarе. Сошниковый отросток отсутствует.
Предчелюстная кость вмещала четыре резца. Ни на одном из обнаруженных экземпляров зубы не сохранились. Судя по размеру и глубине альвеол, первый зуб был самый мелкий, второй и четвертый несколько крупнее, третий – самый крупный.
Носовые и предлобные кости сравнительно короткие и широкие. В передней половине nasale несколько расширено. Здесь на дорсальной поверхности кости развита ямчатая скульптура (табл. VIII , фиг. 2), аналогичная скульптуре на внешней поверхности maxillare. Подобная скульптура присутствует на носовых и челюстных костях Gorynychus masyutinae. Дорсальная поверхность в задней части кости бугристая. На вент-ральной стороне за impressio olphactoris хорошо заметен нервно-сосудистый желоб (sulcus ethmoidalis), в котором прослеживаются два сосудистых канала с близко расположенными отверстиями. Скрепление с челюстной костью выглядит довольно слабым: дорсальный край лицевой пластины максиллы входит в носовую кость только в передней части контакта, а проксимальнее налегает на нее свободно.
Предлобная кость с широкой лицевой частью и довольно высоким орбитальным краем (табл. VIII , фиг. 4 ). Перегиб от края орбиты к предглазничной поверхности довольно резкий. Зона контакта со слезной костью глубоко заходит на орбитальный край кости.
Передний край зубной кости резко опускается к подбородочному выступу (табл. VIII , фиг. 5 ). Покатый, отнесенный назад подбородочный выступ и вытянутая симфизная поверхность, характерные для примитивных тероцефалов, не развиты. Эта особенность отличает Gorynychus от всех других ранних тероцефалов, как восточноевропейских, так и южноафриканских. Книзу передний край dentale уплощается, что особенно заметно на контрасте с округлым бугром на латеральной стороне, который у Gorynychus sundyrensis соответствует по положению на переднезадней оси вентральной площадке зубных костей Julognathus (Сучкова, Голубев, 2019). Сама эта площадка не выражена, вентральный край кости в этом месте гладкий. Резцовая область относительно длинная, количество резцов – три. Наклон двух первых резцов вперед отсутствует. Скульптура внешней поверхности представлена мелкими отверстиями с отходящими от них короткими желобками, похожа на скульптуру зубных костей Julognathus. Все присутствующие в коллекции образцы сильно повреждены, поэтому количество заклыковых зубов и строение проксимальной части зубной кости неизвестно.
Все зубы челюстей характеризуются грубой зазубренностью режущих кантов (табл. IX , фиг. 1–6 ): резцы – 6–9 зазубрин на 5 мм, клыки – 5–9 зазубрин на 5 мм, заклыковые зубы – 10 зазубрин на 5 мм. Эта особенность отличает описываемого тероцефала от всех прочих базальных тероцефалов и соразмерных горгонопий. Например, у сундырского Julognathus количество зазубрин на кантах зубов варьирует от 15 до 20 на 5 мм длины канта, а у котельничского Gorynychus masyutinae – от 9 до 18 на 5 мм.
Клыки уплощенные с боков (табл. IX , фиг. 2б, в), в дистальной трети заметно отгибаются назад (табл. IX , фиг. 1, 2а). Заклыковый зуб имеет короткую шейку и треугольную, практически не уплощенную коронку (табл. IX , фиг. 3 ). Зазубрины в начале и конце ряда на каждом канте заклыкового зуба мельче, чем в срединной части.
Сравнение. От G. masyutinae отличается наличием четырех, а не трех верхних заклыковых зубов, и более грубой зазубренностью всех челюстных зубов (5–10 зазубрин на 5 мм длины режущего канта).
Материал. Кроме голотипа, из типового местонахождения челюстные кости (экз. ПИН, №№ 5388/59, 61, 192, 211), зубные кости (экз. ПИН, №№ 5388/213, 337), предчелюстные кости (экз. ПИН, №№ 5388/55, 56, 231), носовые кости (экз. ПИН, №№ 5388/340, 431), предлобные кости (экз. ПИН, №№ 5388/67, 436), резцы (экз. ПИН, №№ 5388/178, 180, 295, 296, 364, 366, 435), клыки (экз. ПИН, №№ 5388/109, 278, 429), заклыковый зуб (экз. ПИН, № 5388/430).
ОБСУЖДЕНИЕ
Остатки Gorynychus sundyrensis встречаются в местонахождении гораздо реже, чем остатки Julognathus crudelis Sutschkova et Golubev. На первый взгляд, G. sundyrensis также уступает J. crudelis по линейным размерам. Тем не менее, среди обнаруженных костей есть и очень крупные (например, фрагменты зубных костей, особенно экз. ПИН, № 5388/213; табл. VIII , фиг. 5 ). Кроме того, анализ размерозависимой изменчивости костей G. sundyrensis показывает существенное усиление их массивности при незначительном увеличении линейного размера. То есть, крупный череп G. sundyrensis, вероятно, будет выглядеть более массивным, чем равный ему по длине череп J. crudelis.
Интересной особенностью G. sundyrensis также является заметная прижизненная стертость поверхностей коронок зубов на внешней и внутренней сторонах (табл. IX , фиг. 1, 2, 5, 6 ). Она отмечается на всех зубах, кроме заклыкового зуба – экз. ПИН, № 5388/430 (табл. IX , фиг. 3 ) и резца – экз. ПИН, № 5388/180 (табл. IX , фиг. 4 ). На голотипе котельничского G. masyutinae (экз. КМП, № 346) зубов с такими повреждениями нет, но они присутствуют на изолированном зубе (экз. КПМ, № 349) (Kammerer, Masyutin, 2018, фиг. 10 ). У G. sundyrensis стертость клыков всегда затрагивает вершинки коронок, но может охватывать и значительную длину режущего канта, а также часть боковой поверхности (экз. ПИН, № 5388/429; табл. IX , фиг. 2). Резцы почти всегда повреждены в апикальной части, а также на передних или задних поверхностях, с сохранением боковых режущих кантов (табл. IX , фиг. 5, 6 ). У современных хищников стирание клыков происходит при их контакте с твердыми объектами, например, преимущественно задней поверхности зуба у собак, которые активно грызут металлическую клетку (так называемый “cage chewer syndrome”), или преимущественно на внешней поверхности клыков у гепардов при питании мягкой пищей из стальной миски (Steenkamp et al., 2018). На палеонтологическом материале описано более частое повреждение зубов у животных, питающихся более твердой пищей, что для хищников связывается с более полной утилизацией тел жертв (van Valkenburgh, 2009). Поскольку уплощенные, с режущими кантами клыки G. sundyrensis имеют типично хищную морфологию, можно предположить, что повреждения зубов были получены во время питания падалью при обгрызании остатков мяса с костей. Вероятно, в сундырском сообществе G. sundyrensis играл роль падалеяда, уступая нишу доминирующего хищника J. crudelis. Однако уже в следующем, котельничском сообществе G. masyutinae при меньших размерах, возможно, занимал положение доминирующего хищника, разделяя его с Viatkogorgon ivakhnenkoi Tatarinov. Подтверждением истинности сделанной палеоэкологической реконструкции может являться тот факт, что среди всех пермских местонахождений тетрапод Восточной Европы (более 500 местонахождений) только в местонахождении Сундырь-1 обнаружены кости с погрызами хищников (Сучкова, 2017; Ульяхин, Сучкова, 2019).
* * *
Работа выполнена при финансовой поддержке РФФИ (проекты №№ 17-04-01937, 17-04-00410, 17-54-10013) и программы фундаментальных научных исследований Президиума РАН № 22 “Эволюция органического мира и планетарных процессов”, а также за счет средств субсидии, выделенной в рамках государственной поддержки Казанского (Приволжского) федерального ун-та в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров.
Список литературы
Голубев В.К. Пермские и триасовые хрониозухии и биостратиграфия верхнетатарских отложений Восточной Европы по тетраподам. М.: Наука, 2000. 174 с. (Тр. Палеонтол. ин-та РАН. Т. 276).
Голубев В.К., Буланов В.В. Амфибии сундырского фаунистического комплекса пермских тетрапод Восточной Европы // Палеонтол. журн. 2018. № 6. С. 50–62.
Голубев В.К., Куркин А.А., Сенников А.Г. О возрасте сундырского фаунистического комплекса пермских тетрапод Восточно-Европейской платформы // Уч. зап. Казанск. ун-та. Сер. естеств. науки. 2015. Т. 157. Кн. 1. С. 49–58.
Ивахненко М.Ф. Тетраподы Восточно-Европейского плакката – позднепалеозойского территориально-природного комплекса. Пермь: Гос. учр. культуры “Пермский областной краеведческий музей”, 2001. 200 с. (Тр. Палеонтол. ин-та РАН. Т. 283).
Ивахненко М.Ф. Закономерности смен таксонов Theromorpha в наземных сообществах перми Восточной Европы // Палеонтол. журн. 2015. № 1. С. 65–74.
Сенников А.Г., Голубев В.К. Последовательность пермских фаун тетрапод Восточной Европы и пермо-триасовый экологический кризис // Палеонтол. журн. 2017. № 6. С. 30–41.
Сучкова Ю.А. Первая находка погрызов на костях пермских тетрапод Восточной Европы // Интегративная палеонтология: перспективы развития для геологических целей. Матер. 63 сесс. Палеонтол. об-ва (Санкт-Петербург, 3–7 апреля 2017 г.). СПб.: Изд-во ООО “Ладога”, 2017. С. 210–211.
Сучкова Ю.А., Голубев В.К. Новый примитивный тероцефал (Therocephalia, Theromorpha) из средней перми Восточной Европы // Палеонтол. журн. 2019. № 3. С. 88–96.
Ульяхин А.В., Сучкова Ю.А. Тафономия и генезис костеносных отложений позднепермского местонахождения Сундырь-1 (Марий Эл) // Палеострат-2019. Годичн. собр. (науч. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН (Москва, 28–30 января 2019 г.). Прогр. и тез. докл. М.: ПИН РАН, 2019. С. 64–65.
Abdala F., Kammerer C.F., Day M.O. et al. Adult morphology of the therocephalian Simorhinella baini from the middle Permian of South Africa and the taxonomy, paleobiogeography, and temporal distribution of the Lycosuchidae // J. Paleontol. 2014. V. 88. P. 1139–1153.
Abdala F., Rubidge B.S., van den Heever J. The oldest therocephalians (Therapsida, Eutheriodontia) and the early diversification of Therapsida // Palaeontol. 2008. V. 51. P. 1011–1024.
Golubev V.K. Dinocephalian stage in the history of the Permian tetrapod fauna of Eastern Europe // Paleontol. J. 2015. V. 49. № 12. P. 1346–1352.
Ivakhnenko M.F. Eotherapsids from the East European placket (Late Permian) // Paleontol. J. 2003. V. 37. Suppl. 4. P. 339–465.
Kammerer C.F., Masyutin V. A new therocephalian (Gorynychus masyutinae gen. et sp. nov.) from the Permian Kotelnich locality, Kirov Region, Russia // PeerJ. 2018. 6:e4933. P. 1–28.
Steenkamp G., Boy S.C., van Staden P.J., Bester M.N. Oral, maxillofacial and dental diseases in captive cheetahs (Acinonyx jubatus) // J. Compar. Pathol. 2018. V.158. P. 77–89. https://doi.org/10.1016/j.jcpa
Van den Heever J.A. On the validity of the therocephalian family Lycosuchidae (Reptilia, Therapsida) // Ann. S. Afr. Museum. 1980. V. 81. P. 111–125.
Van den Heever J.A. The comparative and functional cranial morphology of the early Therocephalia (Amniota: Therapsida). PhD thesis. Univ. Stellenbosch, 1987. 576 p.
Van den Heever J.A. The cranial anatomy of the early Therocephalia (Amniota: Therapsida) // Ann. Univ. Stellenbosch. 1994. V. 1. P. 1–59.
Van Valkenburgh B. Costs of carnivory: tooth fracture in Pleistocene and Recent carnivorans // Biol. J. Linn. Soc. 2009. V. 96. P. 68–81.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал