

Палеонтологический журнал, 2019, № 4, стр. 64-72
Реконструкция генезиса фаунистических комплексов насекомых (Insecta, Ectognatha) верховых болот Белорусского Поозерья
Г. Г. Сушко *
Витебский государственный университет им. П.М. Машерова
Витебск, Беларусь
* E-mail: gennadisu@tut.by
Поступила в редакцию 22.01.2018
После доработки 12.03.2018
Принята к публикации 31.01.2019
Аннотация
Предложен вероятный сценарий формирования энтомокомплексов верховых болот Белорусcкого Поозерья. Отмечено, что начало их образования, возможно, приходится на ранний голоцен, когда сформировались группировки хортобионтных видов, трофически связанных с пушицей, а также сфагниколов и видов, ассоциированных с сосной. Следующим этапом был атлантический период, когда на болота, по всей видимости, переместились остатки перигляциальной фауны и флоры, в том числе фитофаги вересковых кустарничков. Формирование ядра энтомокомплексов с набором характерных стенобионтных видов, вероятно, закончилось в суббореальный период, так как к этому времени присутствовал основной спектр экологических ниш для рецентного комплекса насекомых.
ВВЕДЕНИЕ
Верховые болота Центральной Европы являются островными экосистемами со специфическими экологическими условиями, не характерными для умеренной зоны Северного полушария как таковой. Бедный комплекс растительности с преобладанием холодолюбивых видов сфагновых мхов, а также верескоцветных и осоковых, размах колебаний суточных температур, низкая минерализация и высокая кислотность почвы (торфа) придают им отчетливые черты тундры (Spitzer, Danks, 2006). Территория Республики Беларусь, находящаяся на рубеже двух крупнейших природных областей (бореальной и неморальной зон), характеризуется значительной площадью торфяных болот, находящихся на границе их сплошного распространения. В Беларуси выделяются четыре хорошо геоморфологически выраженные зональные полосы: Белорусское Поозерье, возвышенности и равнины Центральной Беларуси, равнины и низины Предполесья и Белорусское Полесье (Гельтман, 1982; Матвеев и др., 1988). Наибольшие площади верховых болот страны сконцентрированы в Белорусском Поозерье – регионе, подвергавшемся последнему оледенению. На долю этих экосистем приходится 314.5 тыс. га (Зеленкевич и др., 2016).
До настоящего времени попытки реконструкции динамики рецентных энтомокомплексов верховых болот практически не предпринимались. Среди имеющихся публикаций можно указать две работы. В первой В.Ю. Маавара (1955) в очень краткой форме изложил общие воззрения на возможные пути формирования энтомофауны болот Эстонии, во второй В. Рабелер (Rabeler, 1931) обосновал этапы формирования энтомокомплекса болота (Göldenitzer Hochmoor) на северо-западе Германии.
Если обратиться к исследованиям динамики фауны насекомых на протяжении голоцена в Беларуси, то также нужно отметить их ограниченное число: это, в частности, материалы, изложенные в работах В.И. Назарова (1984, 1988), О.Р. Александровича (1995), С.В. Буги (2002) и О.И. Бородина (2014). Этим материалы, касающиеся динамики энтомокомплексов и фауногенеза насекомых Беларуси в голоцене, ограничиваются. Следует упомянуть также и палеозоологические исследования особенностей формирования авифауны Беларуси, полученные М.Е. Никифоровым (2008).
К настоящему времени имеется немалое число работ, в которых рассматриваются различные аспекты процесса формирования рецентной флоры и растительности верховых болот Беларуси (Пидопличко, 1961; Махнач и др., 1981; Гельтман, 1982; Кухарчик, 1993; Еловичева, 2001; Еловичева и др., 2008). Важная особенность болотных экосистем – формирование торфяников, содержащих в разной степени разложившиеся растительные остатки различного возраста, которые отражают динамику фитоценозов, что позволяет реконструировать их флористический состав. В свою очередь, флористические реконструкции являются основой для предположений о динамике комплексов насекомых–фитофагов, а также зоофагов, обитавших в сфагновом покрове на протяжении голоцена. С другой стороны, накоплен обширный материал по составу насекомых из ориктоценозов плейстоцена современной территории Беларуси (Назаров, 1981, 1984, 1985, 1988), которые указывают на то, что процесс формирования экологических преференций насекомых закончился уже к среднему плейстоцену (Назаров 1981), а найденные в голоценовых отложениях долины Западной Двины ассоциации насекомых включали в себя обычных для этого региона представителей энтомофауны (Назаров, 1985). Следовательно, закономерности современного распространения насекомых во многом связаны с их недавней историей и могут служить основой реконструкций голоценовых перестроек энтомокомплексов. В голоцене энтомофауна Беларуси приобрела современный облик. На это указывает отсутствие криофильных тундровых и азиатских степных и лесных видов насекомых, характерных для микулинского межледниковья (Назаров, 1981). Кроме того, морфологическая стабильность насекомых в течение плейстоцена продемонстрирована достаточно надежно, как и их экологическая устойчивость, и обычно достаточна для реконструкций условий прошлого (Назаров, 1985). Следует отметить, что насекомые весьма чувствительны к изменениям окружающей среды и способны реагировать на них исключительно быстрыми миграциями. При этом энтомофауна реагирует значительно быстрее, чем флора (Назаров, 1988; Coope, 1994).
В настоящей работе предпринята попытка реконструировать генезис энтомокомплексов верховых болот Белорусского Поозерья путем анализа литературных источников, посвященных истории формирования ландшафтов Беларуси, структуре торфяной залежи, а также географическому распространению, трофическим и топическим преференциям специализированных болотных насекомых, выявленных в результате собственных многолетних исследований.
МАТЕРИАЛ И МЕТОДИКА
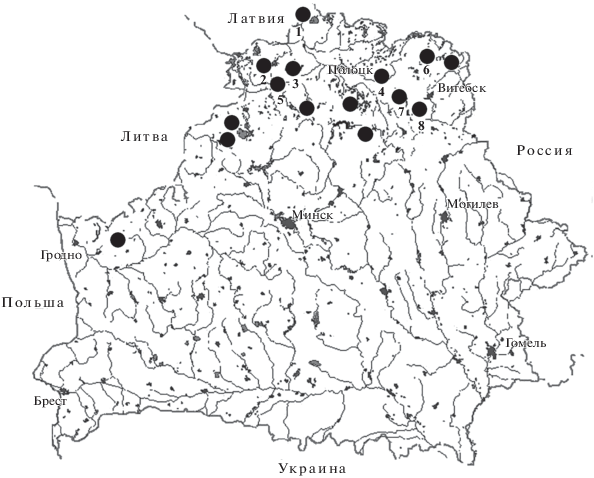
Сбор насекомых проводился автором с 1998 по 2017 гг. с использованием стандартных энтомологических методик (Фасулати, 1971) на 15 верховых болотах в 10 административных районах Витебской, Гродненской и Минской областей (рис. 1), площадь составляла от десяти гектаров до нескольких десятков тысяч гектаров. Подавляющее большинство из них являются охраняемыми территориями и характеризуются невысокой степенью антропогенного воздействия. Суммарно обработано более 80 тыс. экземпляров насекомых, относящихся к 1348 видам. Коллекционные материалы хранятся в зоологическом музее ВГУ им. П.М. Машерова (г. Витебск).
Рис. 1.
Места проведения исследований. Цифрами обозначены болота, для которых определен относительный возраст (по: Беленький, Курзо, 1988).
Исследования проводились в основных типах местообитаний с покрытыми сосной, а также открытыми кустарничково-пушицево-сфагновыми, пушицево-сфагновыми ассоциациями, грядово-мочажинными и грядово-озерковыми комплексами растительности. Характерными элементами их флоры являются такие виды высших сосудистых растений, как Eriophorum vaginatum L., Scheuchzeria palustris L., Rhynchospora alba L., Drosera anglica Huds., D. obovata Mert. et W.D.J. Koch, D. rotundifolia L., Calluna vulgaris (L.) Hull, Ledum palustre L., Andromeda polyfolia L., Oxycoccus palustris Pers., O. microcarpus Turcz. ex Rupr., Chamaedaphne calyculata (L.) Moench, Vaccinium uliginosum L., Empetrum nigrum L., болотные формы сосны (Pinus sylvestris f. uliginosa, P. sylvestris f. litwinowii, P. sylvestris f. willkommii), Betula pubescens Ehrh., B. nana L. Среди мхов доминируют Sphagnum magellanicum Brid., S. angustifolium (C. Jens. ex Russ.) C. Jens, S. fuscum (Schimp.) Klinggr., S. rubellum Wils., S. cuspidatum Ehrh. ex Hoffm. Часто встречается Polytrichum strictum Brid. (Гельтман, 1982; Зеленкевич и др., 2016) (табл. 1).
Таблица 1.
Характеристика консорций высших сосудистых растений рецентной флоры верховых болот Белорусского Поозерья
| Вид растения | Стациальная верность | Биотопическая группа | Географичесий элемент флоры | Основные насекомые-фитофаги |
|---|---|---|---|---|
| Andromeda polifolia (L.) | 5 | Бол | Гол, Бор | Aulacorthum flavum, Athrips pruinosella, Macaria carbonaria, Teia antiquiodes |
| Betula nana L. | 5 | Бол | Гол, Арк-Бор | Ortholepis vacciniella, Coenophila subrosea |
| Calluna vulgaris (L.) Hull. | 3 | Лс-Бол | Гол, Бор-Нем | Altica longicollis, Lochmaea suturalis, Ulopa reticulata |
| Chamaedaphne calyculata (L.) Moench. | 5 | Бол | Гол, Бор | Stephanitis oberti |
| Empetrum nigrum L. | 4 | Лс-Бол | ЕвСиб, Бор | Fimbriaphis latifrons, Altica longicollis, Altenia perspersella, |
| Eriophorum vaginatum L. | 4 | Бол | Гол, Бор | Kelisia vittipennis, Sorhoanus xanthoneurus, Ommatidiotus dissimilis, Plateumaris discolor, Aphthona erichsoni Celaena haworthi, Oeneis jutta |
| Ledum palustre (L.) | 5 | Бол | Гол, Арк-Бор | Cixius similis, Lyonetia ledi, Argyroploce lediana, Eupithecia gelidata, Cacopsylla ledi, Neoamphorophora ledi, Stephanitis oberti, Arichanna melanaria |
| Oxycoccus palustris L. | 4 | Бол | Гол, Бор | Ophiola russeola, Stephanitis oberti, Aspilates gilvaria, Arichanna melanaria, Anarta myrtilli, Carsia sororiata, Vacciniina optilete, Clossiana frigga, Boloria aquilonaris |
| Pinus sylvestris L. | 3 | Лс-Бол | ЕврАз, Бор-Нем | Schizolachnus pineti, Brachonyx pineti, Hylobius abietis |
| Rhynchospora alba (L.) Vahl. | 4 | Бол | Гол, Бор-Нем | Stroggylocephalus livens, Coenonympha tullia |
| Rubus chamaemorus L. | 5 | Бол | Гол, Арк-Бор | Aspilates gilvaria, Clossiana frigga |
| Scheuchzeria palustris L. | 4 | Бол | ЕврАз, Бор-Нем | Acronycta menуanthidis |
| Vaccinium uliginosum L. | 4 | Бол | Гол, Бор | Aulacorthum flavum, Cixius similis, Aspilates gilvaria, Arichanna melanaria, Macaria brunneata, Carsia sororiata, Сolias palaeno |
Примечания. Степень “верности” растений верховым болотам: 3 – встречаются в различных местообитаниях, но нормально развиваются на верховых болотах, 4 – предпочитают верховые болота, 5 – почти исключительно на верховых болотах; эколого-фитоценотическая валентность: Бол – болото, Лс – лес; географический элемент флоры: Гол – голарктический, ЕврАз – евразийский, ЕвСиб – евро-сибирский (долготный элемент), Арк-Бор – аркто-бореальный, Бор – бореальный, Бор-Нем – бореально-неморальный (широтный элемент) (Зеленкевич и др., 2015).
Для характеристики ареалов насекомых использована описательная номенклатура К.Б. Городкова (1984), основанная на физико-географической топонимии и описывающая все составляющие ареала: широтную, долготную и высотную. Сведения о географическом распространении насекомых позаимствованы из работ Б.Ф. Белышева (1963), С. Милевчика (Mielewczyk, 1969), Александровича (1991), Г. Никеля (Nickel, 2003), П. Ивинскиса (Ivinskis, 2004) и интернет-ресурса Coleoptera Poloniae (www.coleoptera.ksib.pl). Данные о трофической и биотопической приуроченности видов получены в результате собственных наблюдений автора и взяты из литературных источников (Мержеевская и др., 1976; Koch, 1984; Nickel, 2003; Ivinskis, 2004; Лопатин, Нестерова, 2005) и интернет-ресурсов (www.kielo.luomus.fi; www.coleoptera.ksib.pl; www.brc.ac.uk.). В качестве наиболее перспективных для анализа видов рассматривались специализированные обитатели верховых болот (Spitzer, Danks, 2006), такие как тирфобионты и тирфофилы. Первые являются стенобионтами данных экосистем, вторые встречаются и в некоторых других местообитаниях со сходными экологическими условиями.
Терминология и хронология приводятся согласно схеме Блитта–Сернандера (Новенко, 2016).
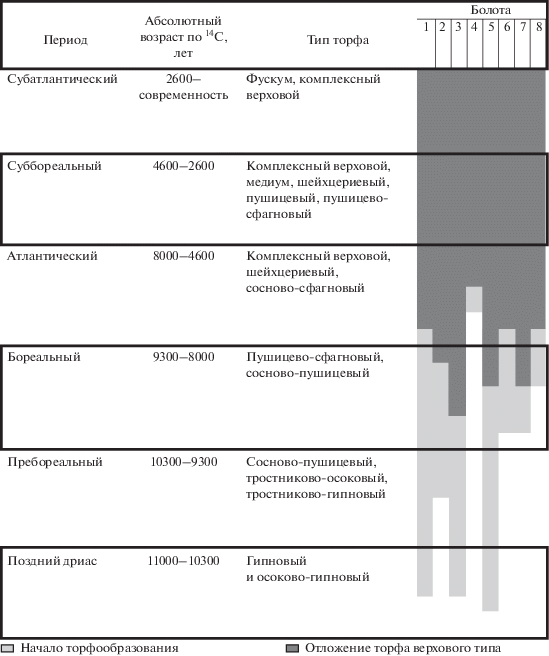
Накопление различных торфов в различающихся климатических условиях позволяет реконструировать флористический состав болотных фитоценозов (Пидопличко, 1961; Еловичева и др., 2008), что является основой для предположений о формировании комплексов насекомых–фитофагов в разные периоды формирования болотных экосистем. Анализ материалов из литературных источников показал, что стратиграфия торфяных залежей верховых болот севера Беларуси довольно однообразна – олиготрофная стадия начинается с отложения торфов с обильным содержанием пушицы и пней сосны, выше находятся слои с повышенным содержанием вересковых кустарничков, самый верхний слой сложен фускум–торфом. При образовании болот в условиях избыточного увлажнения первым олиготрофным слоем торфа может быть шейхцериево-сфагновый (рис. 2) (Пидопличко, 1961; Беленький, Курзо, 1988; Зеленкевич и др., 2016).
Рис. 2.
Этапы формирования отдельных верховых болот Белорусского Поозерья (локализация болот приведена на рис. 1).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Процесс формирования рецентной фауны Беларуси начинается со времени отступления припятского ледника, в днепровскую стадию своего развития покрывавшего всю территорию современной Республики Беларусь. Последующее поозерское оледенение охватило только северную часть страны (Белорусское Поозерье). Ледник продвинулся примерно до линии Вильнюс–Орша–Смоленск. Его отступление началось около 14–13 тыс. лет назад, и через 3–4 тыс. лет он исчез уже и из Скандинавии (Гельтман, 1982; Александрович, 1995). Поэтому фауна Белорусского Поозерья моложе фаун других регионов страны. На это указывает и отсутствие реликтов неогенового периода. В то же время, в ней нет эндемичных видов, что дает основание считать ее миграционной, сложившейся вследствие миграционных потоков из нескольких центров (Александрович, Лопатин, 1994; Лопатин, Нестерова, 2005).
Начало формирования энтомокомплексов верховых болот Белорусского Поозерья приходится, вероятно, на ранний голоцен (пребореал – 10.3–9.3 тыс. лет назад), когда произошло потепление климата и увеличилось количество осадков. Как результат, в северной и центральной части территории современной Беларуси на отдельных, наиболее изолированных болотах начали откладываться самые древние к настоящему времени слои верхового торфа. Бореал (9.3–8 тыс. лет назад) охарактеризовался более интенсивным развитием сфагнового покрова и переходом многих болот в олиготрофную стадию развития (Беленький, Курзо, 1988). На начальных этапах откладывались слои сосново-пушицевого, пушицевого и пушицево-сфагнового торфа (Пидопличко, 1961; Беленький, Курзо, 1988; Еловичева и др., 2008) (рис. 2). Как правило, в наше время наибольшая мощность этих торфов приурочена к окрайкам, несколько уменьшаясь к середине болот (Пидопличко, 1961). Следовательно, на ранних стадиях формирования болот доминировали пушицевые фитоценозы. Фитоценотический облик на пионерных стадиях сукцессии напоминал современные пушицево-сфагновые (во многих местах окаймляющие узкой полосой болотные массивы). Кроме того, в результате распространения семян сосны формировались и сосново-пушицево-сфагновые фитоценозы. Заселение болот насекомыми начиналось, вероятно, с хортобионтных видов, трофически связанных с пушицами. В рецентных энтомокомлексах в числе таких видов монофаги осоковых Kelisia vittipennis (Sahlberg, 1868), Sorhoanus xanthoneurus (Fieber, 1869) (Auchenorryncha) и олигофаги Plateumaris discolor (Panzer, 1795), Aphthona erichsoni (Zetterstedt, 1838) (Coleoptera), Celaena haworthi (Curtis, 1829) (Lepidoptera) (табл. 1).
Энтомокомплекс болот дополнялся и лесными видами, связанными с сосной, среди которых Schizolachnus pineti (Fabricius, 1781) (Sternorrhyncha), Brachonyx pineti (Paykull, 1792), Hylobius abietis (L., 1758) (Coleoptera) и др. С появлением и развитием сфагнового покрова получили возможность заселять болото виды-сфагниколы: Agonum ericeti Panzer, 1809 (Coleoptera), Formica forshlundi Lohmander, 1949, F. uralensis (Hymenoptera) и др. В настоящее время в условиях региона эти насекомые, кроме фитофагов сосны, обитают только или преимущественно на верховых болотах. Кроме того, комплекс эпигейных насекомых должен был пополниться и влаголюбивыми жесткокрылыми [Pterostichus diligens (Sturm, 1824), Atheta arctica Thomson, 1856, Gymnusa brevicornis (Paykull, 1800) и др.], которые в наше время входят в состав преимущественно евро-сибирских, а также трансевразиатских и бореальных зоогеографических комплексов.
В атлантический период среднего голоцена (8–4.6 тыс. лет назад), особенно во второй его половине, климат стал еще более теплым и влажным (Назаров, 1988), что способствовало массовому развитию болотообразующих процессов. В этот период еще больше болот северной и центральной частей современной Беларуси достигли олиготрофной стадии развития. Верховые болота распространились почти по всей территории страны (Беленький, Курзо, 1988).
Изменение климата стало испытанием для субарктических и бореальных элементов флоры, являвшихся остатками перигляциальной растительности. По имеющимся к настоящему времени представлениям, на территории современной Беларуси перигляциальные ландшафты были представлены тундрой с небольшими участками травянистых ассоциаций, а основу ископаемой фауны насекомых того времени составляли представители трех основных экологических групп: обитатели тундры, обитатели прибрежных биотопов и насекомые, связанные с ассоциациями растений семейств злаковых и крестоцветных (Назаров, 1981). В связи с этим криофилы Betula nana, Rubus chamaemorus, Chamaedaphna calyculata были оттеснены на верховые болота, где изолированные популяции данных видов (Гельтман, 1982) и, по всей видимости, многие холодолюбивые насекомые сохранились до наших дней.
В составе торфяной залежи на сформированных ранее сосново-пушицевом (или сосново-сфагновом) и пушицево-сфагновом торфах лежит слой комплексно-верхового, включающего остатки кустарничков (Пидопличко, 1961; Еловичева и др., 2008; Зеленкевич и др., 2016) (рис. 2). Следует отметить, что комплексно-верховой торф имеет генетические связи с безлесными или слабооблесенными растительными группировками, грядово-мочажинными комплексами, обводненными участками с преобладанием мочажин и небольшим количеством гряд (Пидопличко, 1961). В настоящее время подобные фитоценозы приурочены, как правило, к склонам сильно выпуклых болот.
Можно предположить, что в условиях атлантического климатического оптимума на верховых болотах нашли убежища такие чешуекрылые насекомые, ныне имеющие аркто-бореальное распространение, как Сolias palaeno (L., 1761), Clossiana frigga (Thunberg, 1791), Oeneis jutta (Hübner, 1806), Boloria aquilonaris Stichel, 1908, Vacciniina optilete (Knoch, 1781) (табл. 1). В настоящее время они являются как характерными обитателями верховых болот умеренной зоны, так и постоянными компонентами ряда других экосистем северных широт Европы (Mikkola, Spitzer, 1983; Spitzer, Danks, 2006).
Помимо подходящих микроклиматических условий, верховые болота предоставили перигляциальным насекомым и кормовую базу. В это время создались условия для вхождения в состав энтомокомплексов болотных массивов видов, трофически связанных с кустарничками порядка верескоцветных (Ericales). Это, в частности, монофаги багульника Cacopsylla ledi (Flor, 1861) (Sternorrhyncha), Lyonetia ledi Wocke, 1859, Argyroploce lediana (L., 1758), Eupithecia gelidata Möschler, 1860 (Lepidoptera), олигофаги вересковых кустарничков Stephanitis oberti Kolenati, 1857 (Heteroptera), Macaria brunneata (Thunberg, 1784), Carsia sororiata (Hubner, 1813) (Lepidoptera) и др., в числе которых большинство составляют бореальные (по широтной составляющей ареала) виды. Возможно, многие полифаги [Diacrisia sannio (L., 1758), Callophris rubi (L., 1758), Plebeius argus (L., 1758) (Lepidoptera), Cixius similis (Auchenorryncha), Lygus pratensis (Heteroptera)], в спектре кормовых растений которых в настоящее время присутствуют верескоцветные, нашли свои ниши и на верховых болотах.
В суббореале среднего голоцена (4.6–2.6 тыс. лет назад) климат был переменно-влажным, возникали летние засухи (Конойко, 1974; Гельтман, 1982; Беленький, Курзо, 1988). В этот период откладывались различные виды торфа, от сосново-пушицевых до медиум-торфа и комплексного верхового (Пидопличко, 1961).
Ботанический состав торфа показывает, что на болотах произрастало большинство представителей рецентной локальной флоры, и присутствовал основной спектр фитоценозов, существующих в настоящее время: пушицево-сфагновых, сосново-кустарничково-сфагновых и, особенно, грядово-мочажинных комплексов (рис. 2). Очевидно, что к этому времени на наиболее древних верховых болотах закончилось образование ядра энтомокомплекса с набором характерных видов (в том числе, стенобионтов). Скорее всего, в это время завершилось и формирование рецентной фауны Восточно-Европейской равнины в целом (Александрович, 1995). В суббореале повышается обводнeнность торфяных массивов с образованием озерных окон, а также слияние изолированных болот в массивы (Матвеев, 1990). Это должно было способствовать более широкому расселению здесь водных и амфибионтных насекомых, в том числе являющихся в настоящее время специализированными обитателями верховых болот [Somatochlora arctica (Zetterstedt, 1840), Aeschna subarctica (Walker, 1908), Leucorrhinia dubia (Van der Linden, 1825), L. rubicunda (L., 1758) (Odonata), Cymatia bonsdorffii (C. Sahlberg, 1819) (Heteroptera), Enochrus affinis (Thunberg, 1794) (Coleoptera) и др.].
Субатлантический период позднего голоцена (2.6 тыс. лет назад – современность) ознаменовался похолоданием и переменной влажностью климата, что создало еще более благоприятные условия для произрастания на верховых болотах сфагновых мхов, которые заняли здесь господствующее положение (Гельтман, 1982; Беленький, Курзо, 1988). Заболоченность территории приблизилась к современному уровню. Болота верхового и переходного типа были распространены в наибольшей степени по периферии Центральноберезинской равнины и Полоцкой низины (Матвеев, 1990).
В это время происходили и активная экспансия болот на прилегающие леса в равнинных ландшафтах, и рост торфяной залежи. Их выпуклая часть находилась в омбротрофной стадии развития (питание атмосферными водами и полная изоляция от подстилающих почв и их грунтовых вод). При этом мощность торфа значительно увеличилась от краев к центру (Пидопличко, 1961; Беленький, Курзо, 1988). Основным типом торфа являлся фускум–торф. Он генетически связан с безлесными ассоциациями с преобладанием ксерофильного мха Sphagnum fuscum (Пидопличко, 1961). В настоящее время такие ассоциации в основном приурочены к выпуклой вершине болота. Из сосудистых растений здесь преобладают Eriophorum vaginatum и Calluna vulgaris. Как известно, крупные болотные массивы в настоящее время находятся в состоянии безлесного субклимакса, вызванного дефицитом семян сосны в центре массива (Пидопличко, 1961; Богдановская-Гиенэф, 1969; Разумовский, 1981). Приуроченные к ним комплексы насекомых отличаются небольшим видовым богатством и обилием отдельных тирфобионтных и тирфофильных видов (Agonum ericeti, Formica forshlundi, Vacciniina optilete и др.), а также других насекомых, связанных с вереском [Lochmaea suturalis (Thomson, 1866), Dicheirotrichus cognatus Gyllenhal, 1827] и пушицей [Neophilaenus lineatus (L., 1758)] (Peus, 1928; Spitzer, Danks, 2006).
Изменения в энтомокомплексах верховых болот Белорусского Поозерья в минувшее столетие в первую очередь связаны с осушением и увеличением площади сельхозугодий, добычей торфа, участившимся рекреационным воздействием, а также изменением регионального климата. Примером действия последнего фактора, вероятно, может служить обнаружение здесь южного вида жуков-бронзовок Oxythyrea funesta (Poda, 1761), который на протяжении последних нескольких лет регулярно регистрировался в пушицево-сфагновых ассоциациях на большинстве исследованных болот.
Отмечено также присутствие видов, являющихся характерными вредителями сельскохозяйственных культур, таких как крестоцветные блошки [Phyllotreta atra (Fabricius, 1775) и P. nemorum (L., 1758)], синяя льняная блошка (Aphthona euphorbiae Schrank, 1781) (Лопатин, Нестерова, 2005) или капустница [Pieris brassicae (L., 1758)]. Трофические связи этих и ряда других видов на верховых болотах пока не изучены. Можно предположить, что их присутствие в данных экосистемах, причем иногда в достаточно большом количестве, связано с зональной сменой стаций, либо некоторые из этих видов приспособились к питанию на болотных растениях. Эти предположения в некоторой степени подтверждаются находками данных видов на верховых болотах в других регионах Европы (Peus, 1928; Spungis, 2008).
ВЫВОДЫ
Растительные сообщества верховых болот, формирующие специфический микроклимат и предоставляющие кормовую базу фитофагам, определяли направление сукцессионных перестроек энтомокомплексов на протяжении голоцена. Начало образования комплексов насекомых наиболее древних болотных массивов приходится, вероятно, на ранний голоцен, когда отдельные болота перешли в олиготрофную стадию развития. В это время, по всей видимости, сформировались группировки хортобионтных насекомых, трофически связанных с пушицей, а также сфагниколов и видов, ассоциированных с сосной.
Следующим этапом формирования комплексов насекомых данных экосистем был атлантический период, когда на болота переместились остатки перигляциальной фауны и флоры, в том числе фитофаги кустарничков порядка верескоцветных. Это позволяет считать данный этап наиболее важным в генезисе энтомокомплексов верховых болот Беларуси.
В суббореале, вероятно, закончилось формирование ядра энтомокомплексов с набором характерных стенобионтных видов, так, к этому времени присутствовал основной спектр растительных сообществ, существующих в настоящее время, и, как следствие, возникли основные экологические ниши для рецентного комплекса насекомых. Кроме того, предоставилось возможным более широкое расселение водных и амфибионтных насекомых, обусловленное повышением обводненности верховых болот за счет озерных окон и развития грядово-мочажинных комплексов.
В субатлантический период, с продолжением экспансии болот на прилегающие территории и вступлением центральной их части в стадию субклимакса, расширился характерный энтомокомплекс верещатников, представленный небольшим числом видов, большей частью связанных с вереском. На современном этапе происходит пополнение комплекса теплолюбивыми формами, а также мигрирующими из агроценозов вредителями сельскохозяйственных культур.
Таким образом, предложен вероятный сценарий формирования энтомокомплексов верховых болот Белорусcкого Поозерья, который может являться теоретической основой для дальнейших реконструкций с использованием палеонтологических методов.
* * *
Мы выражаем глубочайшую признательность за ценные консультации и помощь в определении отдельных видов насекомых проф. С.В. Буге (Минск), проф. О.Р. Александровичу (г. Слупск, Польша), проф. А.Н. Галкину (Витебск), проф. Е.П. Нарчук (С.-Петербург), А.Г. Мосейко (С.‑Петербург), О.И. Бородину (Минск), С.К. Рындевичу (Барановичи, Беларусь), А.В. Кулаку (Минск), В.В. Блинову (Минск), А.О. Лукашуку (п. Домжерицы, Беларусь), Е.А. Держинскому (Витебск). Благодарим Д.Г. Груммо (Минск) и О.В. Созинову (Гродно) за предоставленные геоботанические описания и материалы по стратиграфии торфяных залежей верховых болот. Отдельная благодарность за ценные советы и конструктивные замечания по улучшению рукописи А.Г. Пономаренко (ПИН РАН, Москва), а также двум анонимным рецензентам.
Список литературы
Александрович О.Р. Жуки жужелицы (Coleoptera, Carabidae) фауны Белоруссии // Фауна и экология жесткокрылых Беларуси. Минск: Навука i технiка, 1991. С. 37–78.
Александрович О.Р., Лопатин И.К. Структура колеоптерофауны Беларуси и история ее формирования // Проблемы изучения, сохранения и использования биологического разнообразия животного мира. Тез. докл. VII зоол. конф. Минск: Ин-т зоологии АН Беларуси, 1994. С. 103–104.
Александрович О.Р. Реконструкция путей формирования фауны жужелиц (Coleoptera, Carabidae) на западе Русской равнины // Фауна и систематика. Тр. Зоол. музея Бел. ун-та. 1995. Вып. 1. С. 52–68.
Беленький С.Г., Курзо Б.В. Возраст торфяных месторождений верхового типа Беларуси и происхождение пограничных горизонтов // Вести АН БССР. Сер. биол. наук. 1988. № 2. С. 27–31.
Белышев Б.Ф. Определитель стрекоз Сибири по имагинальным и личиночным фазам. М.-Л.: Изд-во АН СССР, 1963. 112 с.
Богдановская-Гиенэф И.Б. Закономерности формирования сфагновых болот верхового типа на примере Полистово–Ловатского массива. Л.: Наука, 1969. 186 с.
Бородин О.И. Позднеплейстоценовые и голоценовые перестройки фауны цикадовых Беларуси // Тр. Бел. ун-та. 2014. Вып. 9(2). С. 263–271.
Буга С.В. Структура и экологические основы формирования фауны дендрофильных тлей Беларуси. Автореф. дис. … докт. биол. наук. Минск: НАНБ, Ин-т зоол. 2002. 41 с.
Гельтман В.С. Географический и типологический анализ лесной растительности Белоруссии. Минск: Наука и техника, 1982. 326 с.
Городков К.Б. Ареалы насекомых европейской части СССР. Л.: Наука, 1984. 60 с.
Еловичева Я.К. Эволюция природной среды антропогена Беларуси. Минск: Белсэнс, 2001. 292 с.
Еловичева Я.К., Кольмакова Е.Г., Крюк А.С. Эволюция растительного покрова ландшафтного заказника “Ельня” // Вестн. Бел. ун-та. Сер. 2, Химия. Биология. География. 2008. № 1. С. 75–79.
Зеленкевич Н.А., Груммо Д.Г., Созинов О.В., Галанина О.В. Флора и растительность верховых болот Беларуси. Минск: Строй Медиа Проект, 2016. 244 с.
Конойко М.А. Типология верховых болот и особенности их распространения на территории Белоруссии // Типы болот СССР и принципы их классификации. Л.: Наука, 1974. С. 94–100.
Кухарчик Т.И. Верховые болота Беларуси. Минск: Навука i тэхнiка, 1993. 136 с.
Лопатин И.К., Нестерова О.Л. Насекомые Беларуси: листоеды (Coleoptera, Chrysomelidae). Минск: Технопринт, 2005. 318 с.
Маавара В.Ю. Энтомофауна верховых болот Эстонской ССР и ее изменение под влиянием хозяйственной деятельности человека. Автореф. дис. … канд. биол. наук. Тарту: ИЗ АН Эстонии, 1955. 19 с.
Матвеев А.В., Гурский Б.Н., Левицкая Р.И. Рельеф Белоруссии. Минск: Университетское, 1988. 320 с.
Матвеев А.В. История формирования рельефа Белоруссии. Минск: Навука i технiка, 1990. 144 с.
Махнач Н.А., Еловичева Я.К., Бурлак А.Ф., Рылова Т.Б. Флора и растительность Белоруссии в палеогеновое, неогеновое и антропогеновое время. Минск: Наука и техника, 1981. 106 с.
Мержеевская О.И., Литвинова А.Н., Молчанова Р.В. Чешуекрылые (Lepidoptera) Белоруссии (каталог). Минск: Наука и техника, 1976. 132 с.
Назаров В.И. Изменения энтомофауны северо-востока Белоруссии в позднем кайнозое // Геологические исследования кайнозоя Белоруссии. Минск: Наука и техника, 1981. С. 84–91.
Назаров В.И. Реконструкция ландшафтов Белоруссии по палеоэнтомологическим данным (антропоген) // Тр. Палеонтол. ин-та АН СССР. 1984. Т. 205. 96 с.
Назаров В.И. Межледниковая энтомофауна среднего и позднего плейстоцена Белоруссии (Некоторые аспекты палеогеографии и климатостратиграфии) // Геология и гидрогеология кайнозоя Белоруссии. Минск: Наука и техника, 1985. С. 61–66.
Назаров В.И. Климатостратиграфия позднего плейстоцена Белоруссии // Литология, геохимия и стратиграфия континентальных кайнозойских отложений Белоруссии. Минск: Наука и техника, 1988. С. 245–254.
Никифоров М.Е. Формирование и структура орнитофауны Беларуси. Минск: Белорусская наука, 2008. 297 с.
Новенко Е.Ю. Растительность и климат Центральной и Восточной Европы в позднем плейстоцене и голоцене. Автореф. дисс. … докт. геогр. наук. М.: МГУ, 2016. 50 с.
Пидопличко А.П. Торфяные месторождения Белоруссии. Минск: Навука i Тэхнiка, 1961. 192 с.
Разумовский С.М. Закономерности динамики биоценозов. М.: Наука, 1981. 232 с.
Фасулати К.К. Полевое изучение наземных беспозвоночных. М.: Высшая школа, 1971. 424 с.
Coleoptera Poloniae [Electronic resource]. Information System about Beetles of Poland. http://www.coleoptera.ksib.pl (дата обращения: 24.03.2017).
Coope G.R. The response of insect faunas to glacial–interglacial climatic fluctuations // Phil. Trans. R. Soc. London. B. 1994. V. 344. P. 19–26.
Database of Insects and their Food Plants [Electronic resource]. http://www.brc.ac.uk. (дата обращения: 20.01.2017).
Ivinskis P. Lepidoptera of Lithuania: Annotated catalogue. Vilnius: Vilniaus univ., 2004. 380 p.
Koch M. Wir bestimmen Schmetterlinge. Leipzig: Radebeul, 1984. 792 p.
Luonnontieteellinen keskusmuseo [Electronic resource]. Helsingin yliopisto, http://www.kielo.luomus.fi (дата обращения: 20.05.2016).
Mikkola K., Spitzer K. Lepidoptera associated with peatlands in central and northern Europe: a synthesis // Nota Lepid. 1983. V. 6. № 4. P. 216–229.
Mielewczyk S. Larwy ważek (Odonata) niektórych torfowisk sfagnowych Polski // Polskie Pismo entomol. 1969. V. 39. № 1. P. 2–77.
Nickel H. The Leafhoppers and Planthoppers of Germany (Hemiptera Auchenorrhyncha): Patterns and strategies in a highly diverse group of phytophagous insects. Sofia, M.: Pensoft Publ., 2003. 460 p.
Peus F. Beitrage zur Kenntnis der Tierwelt nordwestdeutscher Hochmoore. Eine ökologische Studie. Insecten, Spinnentiere, Wirbeltiere // Z. Morphol. Oekol. Tiere. 1928. № 12. P. 533–683.
Rabeler W. Die Fauna des Göldenitzer Hochmoores in Mecklenburg // Z. viss. Biol. (A). 1931. № 21. P. 173–315.
Spitzer K., Danks H.V. Insect biodiversity of boreal peat bogs // Annual Rev. Entomol. 2006. № 51. P. 137–161.
Spungis V. Fauna and ecology of terrestrial invertebrates in raised bog in Latvia. Riga: Latvias Entomol. Biedriba, 2008. 80 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал