

Палеонтологический журнал, 2019, № 4, стр. 56-63
Представители надсемейства Richthofenioidea (Brachiopoda, Productida) из перми Дарваза, Таджикистан
В. С. Журавлев a, А. В. Пахневич a, *
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: alvpb@mail.ru
Поступила в редакцию 11.12.2017
После доработки 01.02.2018
Принята к публикации 02.07.2018
Аннотация
Изучена новая коллекция пермских брахиопод надсемейства Richthofenioidea Waagen, 1885 (отряд Productida) из отложений гундаринской свиты кубергандинского яруса Дарваза, Таджикистан, представленная Gundaria insolita Angiolini et al., 2016, Hercosestria sp., Coscinarina sp. и Proteguliferina sp. Выявлены особенности сохранности раковин. По кольцам роста на раковине определен индивидуальный возраст животных. Изучена внешняя морфология и внутреннее строение раковин, в том числе, с привлечением метода рентгеновской томографии.
ВВЕДЕНИЕ
Надсемейство Richthofenioidea Waagen, 1885 – группа высоко специализированных позднепалеозойских продуктид. Внешне ее представители напоминают одиночные кораллы ругозы. Животные вели неподвижный образ жизни, прирастая макушкой брюшной створки к субстрату. Рихтгофениоидеи относятся к экологическому типу прирастающих брахиопод (по: Иванова, 1949). Они известны с позднего карбона до конца перми, наибольшее биоразнообразие надсемейства приходится на пермский период. В настоящее время описано 24 рода надсемейства. Представители Richthofenioidea редко встречаются в ископаемом состоянии. Из пермских отложений Стеклянных гор Техаса известно наибольшее число родов надсемейства Richthofenioidea (Cooper, Grant, 1969, 1975). Здесь встречается восемь родов: Hercosia Cooper et Grant, 1969, Hercosestria Cooper et Grant, 1969, Cyclacantharia Cooper et Grant, 1969, Taphrosestria Cooper et Grant, 1975, Sestropoma Cooper et Grant, 1969, Collumatus Cooper et Grant, 1969, Richthofenia Kayser, 1881, Acritosia Cooper et Grant, 1969. Они образовывали скопления, прирастая друг к другу, поселяясь как на раковинах своего вида, так и других видов рихтгофениоидей и прочих брахиопод. Часто они обитали в ассоциации с кораллами и другими фильтраторами, в том числе, на рифах.
Одно из богатейших местонахождений рихтгофениоидей на территории бывшего СССР – отложения гундаринской свиты кубергандинского яруса нижней – начала средней перми Дарваза (Таджикистан). Но они еще остаются слабо изученными. Первый вид надсемейства из пермских отложений Памира описан в 1973 г. (Грунт, Дмитриев, 1973), а непосредственно из пермских отложений Дарвазского хребта (является частью Памира) только недавно (Angiolini et al., 2016).
Изучая пермских брахиопод Дарваза, мы пришли к выводу, что разнообразие рихтгофениоидей больше, чем это было известно на 2016 г. Цель настоящей работы исследовать особенность сохранности раковин брахиопод надсемейства Richthofenioidea, их морфологические и экологические особенности и таксономическое разнообразие.
ИСТОРИЯ ИЗУЧЕНИЯ НАДСЕМЕЙСТВА RICHTHOFENIOIDEA ПЕРМИ ПАМИРА И ДАРВАЗА
В настоящее время изучены только отдельные группы каменноугольной и пермской фаун Дарваза (Горюнова, 1975; Левен и др., 1992). Брахиоподы описаны лишь частично (Angiolini et al., 2016), а изучение продуктид надсемейства Richthofenioidea только начинается. Первый (см. Чернышев, 1914) представитель рода Richthofenia Kayser, 1881, отнесенный к северокавказскому виду R. caucasica Licharew, 1928, был обнаружен на Памире (Грунт, Дмитриев, 1973, табл. V , фиг. 5–7 ). Он был отмечен в отложениях джульфинского яруса, тахтабулакской свиты, среднетахтабулакской подсвиты Юго-Восточного Памира.
Первое исследование представителя надсемейства Richthofenioidea Юго-Западного Дарваза, определенного как Richthofenia sp., с помощью рентгеновского микротомографа Skyscan 1172, было проведено недавно (Pakhnevich, 2010). В результате исследования в стенке брюшной створки были идентифицированы псевдопоры, отдельные элементы микроструктуры.
В трудах Седьмого брахиоподового конгресса были опубликованы тезисы (Angiolini et al., 2015), в которых упоминалось присутствие в пермских отложениях (гундаринская свита) рихтгофеноидей, близких к роду Richthofenia. Один из экземпляров таксона, который предполагался в качестве нового, был изображен. Позже был представлен доклад, посвященный биоразнообразию надсемейства Richthofenioidea в отложениях гундаринской свиты перми Юго-Западного Дарваза, основные выводы которого опубликованы в тезисах (Журавлев, Пахневич, 2016). Показано, что рихтгофениоидеи принадлежат к трем семействам, одно из которых точно определить не удалось, а два остальных – это Richthofeniidae и Cyclacanthariidae. В том же году выходит статья (Angiolini et al., 2016), где описывается новый род и вид Gundaria insolita Angiolini et al., 2016, отнесенный к семейству Cyclacanthariidae, подсемейству Cyclacanthariinae. На представительном материале проиллюстрировано внутреннее строение створок раковины, внешняя морфология. В статье подробно описаны геология, стратиграфия, литология гундаринской и сафетдаронской свит Дарваза, палеоэкологические условия обитания сообществ брахиопод, комплексы важных стратиграфических групп – конодонтов и фораминифер. Проведена корреляция тетической региональной стратиграфической шкалы перми Дарваза и Международной стратиграфической шкалы перми. В работе описаны брахиоподы отрядов Productida, Rhynchonellida, Athyridida, Spiriferida, Spiriferinida, Terebratulida.
МАТЕРИАЛ И МЕТОДИКА
Находки рихтгофеноидей – редкость, поэтому каждая новая представляет собой большую ценность. В коллекции, собранной В.Ю. Дмитриевым и Т.А. Грунт в период 60–80-х гг. на Дарвазе из отложений гундаринской свиты кубергандинского яруса перми верховьев р. Зида-Дара, левого борта реки, было обнаружено 14 экз. различной сохранности (восемь раковин хорошей сохранности и шесть фрагментов брюшных створок), идентифицированных как представители надсемейства Richthofenioidea. Экземпляры представлены брюшными створками или их фрагментами, фрагментами двух спинных створок. Раковины были получены в результате растворения образцов карбонатной породы с фауной в уксусной кислоте.
Рентгеновская микротомография производилась в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН) на микротомографе Skyscan 1172. Параметры сканирования: шаг среза 10–34.1 мкм, фильтр Al (мм), шаг вращения 0.7°, U = = 100 kV, I = 100 mA, вращения – 180°. Для получения виртуальных срезов использовались программы NRecon и DataViewer.
Также в исследовании применялся цифровой бинокулярный микроскоп Leica M165C, производилась съемка на сканирующем электронном микроскопе (СЭМ) TESCAN/VEGA в режиме низкого вакуума. Замеры раковин производились с помощью штангенциркуля с точностью 0.1 мм. Материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН в колл. № 4104.
Авторы выражают сердечную благодарность за помощь в работе на СЭМ Р.А. Ракитову (ПИН РАН), за консультации В.Ю. Дмитриеву (ПИН РАН).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
Сохранность. Раковины рихтгофений частично или полностью окремнены, напоминая таковые из известного техасского пермского местонахождения в Стеклянных горах. В некоторых случаях внутренняя полость раковины была заполнена мелкокристаллическими массами кварца. У переднего края брюшных створок рост кварца происходил по трещинам внутри породы. В результате после растворения карбонатной породы оставалась сетчатое кристаллическое образование, которое можно было спутать с косцинидиумом рихтгофений (сросшиеся, разветвленные, уплощенные внутренние иглы на вершине (у переднего края) брюшной створки, образующие защитную сетку). Особенно это затруднило выявление рихтгофеноидей среди кораллов-ругоз, которые внешне на них похожи. У некоторых мелких одиночных кораллов, вытравленных из породы, на вершине кубков также оставалось сетчатое кристаллическое образование. В результате изучения внутреннего строения всех экземпляров, которые были предварительно отнесены к рихтгофеноидеям, с помощью рентгеновской микротомографии среди них были выявлены четырехлучевые кораллы. Затем, при просмотре материала с помощью бинокуляра, было обнаружено важное отличие, по которому рихтгофеноидей можно легко отличить от ругоз: кубки кораллов имели сглаженную поверхность, а раковины рихтгофеноидей имели слои нарастания, напоминающие чешуи или черепицу.
Из-за неполного окремнения раковины были растворены некоторые элементы строения раковин. Например, слабо окремненными были спинные створки раковин. Их почти полное отсутствие сильно затрудняло определение рихтгофеноидей и возможность отнесения к какому-либо роду.
Индивидуальный возраст. Материал уникален тем, что в нем присутствуют ювенильные экземпляры, на что указывает их индивидуальный возраст. Он определялся по кольцам роста на брюшных створках продуктид (табл. VII , фиг. 1; см. вклейку). Если морщинистость – это складки на поверхности створки, то годовые кольца – четкие, хорошо различимые, регулярные структуры. Метод (см. Алимов, 1981; Золотарев, 1989) успешно использовался для определения возраста пресноводных и морских двустворчатых моллюсков и современных брахиопод (Зезина, 1976; Пахневич, 2012). Учитывались только хорошо различимые кольца, которые наблюдались по всей поверхности брюшной створки. Для целых брюшных створок были измерены размеры и возраст. Удалось измерить десять экземпляров из 14. Высота изучаемых экземпляров варьирует от 5 до 19 мм, а возраст – от 1 до 9 лет. Из них 1–3-летние экземпляры являются ювенильными. Единственная однолетняя особь имела длину (высоту) раковины 5 мм. Универсальность этого метода была показана при сравнении индивидуального возраста ископаемых и современных брахиопод (Пахневич, 2012). Предельный возраст современных брахиопод не превышает 15 лет. Он был установлен у дальневосточных брахиопод Laqueus californianus (Koch, 1843). Для ископаемых брахиопод максимальный установленный возраст не превышает 29 лет (Пахневич, 2012). Он был выявлен на примере раннепермской продуктиды Dictyoclostus sp. Чаще возраст ископаемых палеозойских, мезозойских и кайнозойских брахиопод тринадцати отрядов был не более 14 лет. Несмотря на некоторые отличия в предельном возрасте, значительной разницы в этих параметрах не выявлено. Максимальный индивидуальный возраст рихтгофеноидей совпадает с таковым у современных брахиопод. Новые данные важны, поскольку рихтгофеноидеи представляют особую экологическую группу прирастающих продуктид.
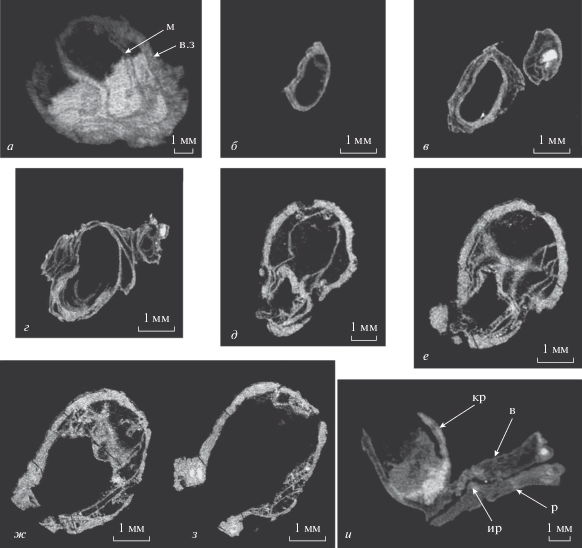
Рис. 1.
Виртуальные микротомографические срезы раковин представителей надсемейства Richthofenioidea: а–г – Gundaria insolita Angiolini et al., 2016: а – экз. ПИН, № 4104/1300, две раковины, видно положение раковины молодой брахиоподы внутри взрослой; б–г – экз. ПИН, № 4104/1303, последовательные срезы брюшной створки через заднюю часть створки; д–з – Proteguliferina sp., экз. ПИН, № 4104/1304, поперечное сечение брюшной створки; и – экз. ПИН, № 4104/1305, сросшиеся четырехлучевой коралл, обызвествленная водоросль и раковина Gundaria insolita. Обозначения: кр – четырехлучевой коралл; р – фрагмент брюшной створки G. insolita; в – водоросль; ир – игла G. insolita; в.з – взрослый экземпляр; м – молодой экземпляр.
Морфология и таксономический состав. В состав надсемейства Richthofenioidea входят четыре семейства: Richthofeniidae, Hercosiidae, Cyclacanthariidae (с тремя подсемействами), Gemmellaroiidae, и 24 рода (Wardlaw et al., 2000; Angiolini et al., 2016).
По форме среди брюшных створок выделяются высокие конические кубковидные, до 19 мм высотой, и короткие чашевидные (рис. 1, 2). Целые спинные створки не обнаружены, за исключением двух фрагментов. Вероятно, створки были слабо окремнены, и растворились вместе с вмещающей породой. На нескольких раковинах и их фрагментах обнаружены элементы внутреннего и внешнего строения, которые отсутствуют у Gundaria insolita. То есть, разнообразие таксонов больше, чем предполагалось ранее (Angiolini et al., 2016).
Рис. 2.
Внешний вид раковин продуктид надсемейства Richthofenioidea: а, б, г ‑ Gundaria insolita Angiolini et al., 2016: а – экз. ПИН, № 4104/1308, раковина с хорошо выделяющимися опорными иглами; б – экз. ПИН, № 4104/1309, общий вид раковины; г – экз. ПИН, № 4104/13011, часть раковины; в – Hercosestria sp., экз. ПИН, № 4104/13010, внешний вид раковины; д – Proteguliferina sp., экз. ПИН, № 4104/1304, вид с переднего края брюшной створки. Обозначение: и – игла.

Большая часть раковин и фрагментов брюшных створок принадлежат виду G. insolita. На это указывает ряд элементов строения раковины: раковины высокие конические; на брюшной створке имеются прикрепительные, иногда ризоидоподобные, иглы. Именно у данного вида два типа игл: прямые или в разной степени согнутые иглы, иногда морщинистые у основания, и спирально скрученные, которые располагались у места прикрепления раковины к субстрату (табл. VII , фиг. 7 ). Спиральные иглы в процессе роста могли вкручиваться в субстрат. Они расположены у основания раковины (макушки брюшной створки). Простые иглы располагаются у переднего края. Внутри брюшных створок у переднего края располагались внутренние иглы. На экз. № 4104/1300 наблюдалась и внутренняя игла взрослой брахиоподы, прижатая к внутренней стенке брюшной створки, вследствие поселения молодой особи внутри раковины взрослой (табл. VII , фиг. 2г). На внутренней поверхности брюшных створок обнаружена шагреневая поверхность (рис. 3, д). Септа в брюшной створке отсутствует, имеется арея. Пузырчатая ткань обрастает брюшную створку в виде широких, почти гладких черепитчатых слоев. На фрагменте спинной створки выявлен замочный отросток, характерный для G. insolita (см. Angiolini et al., 2016, рис. 18 , 19Aj) (рис. 3, а, б). Спинная створка вытянута в ширину и имеет выемку на переднем крае.
Рис. 3.
Особенности строения раковин продуктид надсемейства Richthofenioidea: а, б – Gundaria insolita Angiolini et al., 2016, экз. ПИН, № 4104/1300, замочный отросток, изображенный при разном увеличении; в, г – фрагмент раковины Coscinarina sp., экз. ПИН, № 4104/1306, внешний вид (в) и увеличенный косцинидиум (г); д – Gundaria insolita Angiolini et al., 2016, экз. ПИН, № 4104/1307, шагрень на внутренней поверхности брюшной створки. Обозначение: з. о. – замочный отросток.
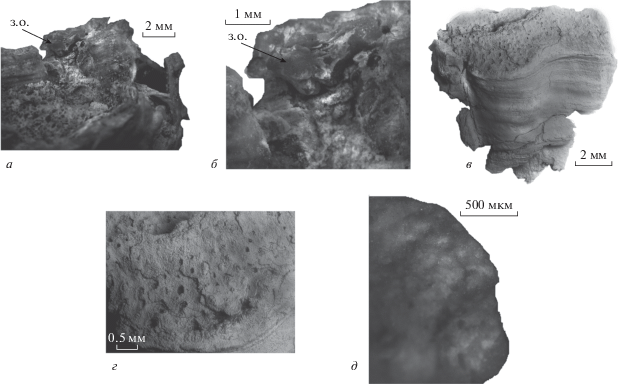
Выявлены особенности строения ювенильных экземпляров и процессов формирования раковин. Хорошо видно, что опорные иглы около макушки брюшной створки многочисленные (табл. VII , фиг. 3–5 ); некоторые огибают ее. Отмечены особенности зарастания макушки раковинным веществом пузырчатой ткани на начальных стадиях роста ювенильных экземпляров (табл. VII , фиг. 6 ). Пузырчатая ткань в виде налегающих друг на друга слоев образует черепитчатую наружную скульптуру.
Экземпляр с низкой конической чашевидной раковиной (рис. 1; 2, д) имел сжатую с боков спинную створку, в то время как раковины G. insolita выше, а их спинные створки вытянуты в ширину. Септа в брюшной створке не обнаружена. Прикрепительные иглы не выявлены. Отпечаток спинной створки данного экземпляра напоминает спинную створку представителей подсемейства Teguliferininae. Форма раковины также близка к представителям данного подсемейства. Более всего данный экземпляр сходен с родом Proteguliferina Licharew in Sarytcheva et al., 1960 (верхний карбон–нижняя пермь; Русская платформа, Донецкий бассейн, а также, возможно, Северная Америка). Поэтому данный экземпляр определен как Proteguliferina sp. Для подтверждения данного определения необходим дополнительный материал.
Для экз. № 4104/1306 вытянутой конической формы (рис. 3, в, г) характерно наличие косцинидиума. Он образован плотно сросшимися иглами. Подобное образование отсутствует у G. insolita и в подсемействе Teguliferininae семейства Cyclacanthariidae, почти у всех представителей семейства Richthofeniidae и в семействе Gemmellaroiidae. Относительно косцинидиума, септы в брюшной створке и других морфологических структур рихтгофеноидей, следует отметить несовершенство существующих диагнозов семейств (Wardlaw et al., 2000), так как эти признаки встречаются иногда у отдельных представителей семейств, но для большинства родов тех же семейств не характерны. Или, наоборот, отдельные представители родов не имеют структур, которые характерны для всех остальных представителей семейства. Поскольку раковина изученного экземпляра полностью не сохранилась, точно определить ее видовую принадлежность трудно, так же как и описать новый вид. Но строение косцинидиума очень характерное: отверстия защитной сетки расположены косыми рядами. Подобный косцинидиум не характерен для семейства Hercosiidae, у представителей которого иглы срастаются не столь плотно. То же можно отметить и для представителей семейства Cyclacanthariidae, у которых или толщина сросшихся игл меньше, чем ширина пространства между ними, или косцинидиум образован редкими толстыми иглами. Очень похож только косцинидиум рода Coscinarina Muir-Wood et Cooper, 1960 (средняя пермь Европы) из семейства Richthofeniidae. Это редкий представитель семейства, у которого данная структура присутствует. Таким образом, экз. № 4104/1306 может быть отнесен к Coscinarina sp.
Строением фрагмента спинной створки от G. insolita отличается еще один экземпляр (табл. VII , фиг. 8 ). Он имеет высокую коническую раковину, но с меньшим числом прикрепительных игл, морщинистой пузырчатой тканью и, самое главное, не совпадает по строению срединной септы спинной створки. Она разделена желобком на две части, из-за чего выглядит утолщенной. У представителей семейства Cyclacanthariidae, к которому отнесена G. insolita, септа тоньше и без желобка. По форме спинной створки, выявленной с помощью рентгеновской микротомографии, изученный экземпляр сходен с G. insolita. В брюшной створке септа не обнаружена. По строению септы спинной створки данный экземпляр можно отнести к семейству Hercosiidae. На внутренней поверхности спинной створки, как и у представителей этого семейства, имелись крупные сосочки шагрени. По отсутствию септы в брюшной створке данный экземпляр наиболее близок к роду Hercosestria Cooper et Grant, 1969 (нижняя пермь Западного Техаса), но у последнего пузырчатая ткань не такая морщинистая. Возможно, данный экземпляр представляет новый род и вид Hercosiidae, но из-за недостатка материала он пока отнесен к Hercosestria sp. Подобная срединная септа была изображена ранее (Грунт, Дмитриев, 1973, табл. V , фиг. 6 ), но она указывалась (вероятно, ошибочно) как арея брюшной створки. Не исключено, что Richthofenia caucasica из отложений джульфинского яруса Памира также принадлежит к роду Hercosestria.
Помимо Gundaria insolita, в отложениях гундаринской свиты встречается несколько видов рихтгофеноидей, которые относятся к Hercosestria sp. (Hercosiidae), Coscinarina sp. (Richthofeniidae) и Proteguliferina sp. (Cyclacanthariidae, Teguliferininae). Эти роды принадлежат к фауне продуктид конца первой трети (цисуралий) – начала второй трети (гваделупий) перми. В фауне есть и европейские виды, и североамериканские. Это дает основание говорить о более широком распространении родов, которые ранее, почти без исключения, отмечались или на территории Северной Америки, или Европы. К концу первой трети перми происходит смешивание североамериканских и европейских фаун рихтгофеноидей. И только род Gundaria, вероятно, является эндемичным для Дарваза.
Внутреннее строение раковин по данным рентгеновской микротомографии. С помощью рентгеновской микротомографии изучено 14 экз. брахиопод. За счет контраста между минералами кальцит–кварц хорошо заметны окремненные участки раковин и карбонатная порода (рис. 1, а), которая сохранилась в промежутках пузырчатой ткани. С помощью рентгеновской микротомографии удалось наблюдать первичную форму раковины G. insolita до обрастания пузырчатой тканью (рис. 1, б–г). Стенки раковины выглядят плотнее и толще, чем слои пузырчатой ткани. В поперечном сечении раковина G. insolita имела овальную фасолевидную форму (рис. 1, а–г). Она повторяет форму спинной створки, которая вытянута в ширину, и у этого вида на переднем крае имеет выемку. Однако форму замочного отростка таким образом определить не удалось. Важный диагностический признак, септа в брюшной створке, не обнаружен.
Для экземпляра Proteguliferina sp. с чашевидной раковиной, ранее отнесенного (Журавлев, Пахневич, 2016) к подсемейству Teguliferininae, также удалось наблюдать поперечное сечение брюшной створки. Оно овальное (рис. 1, д–з), но, в отличие от G. insolita, спинная створка вытянута в длину, а не в ширину. Замочный отросток был массивный, но его детальное строение осталось неизвестным.
В двух раковинах G. insolita (экз. №№ 4104/1300 и 4104/1301) наблюдались молодые экземпляры того же вида, которые поселялись внутри кубковидной брюшной створки. Вероятно, субстратом для них служила спинная створка взрослых особей, но из-за растворения сказать об особенностях положения молодых рихтгофеноидей на ее поверхности невозможно. Хорошо просматривается, что молодые брахиоподы занимают почти все пространство внутри “кубка” над поверхностью спинной створки взрослого животного (рис. 1, а). Иглы ювенильных животных упираются в стенку раковины взрослого животного (табл. VII , фиг. 2а–2в). А внутренние иглы взрослых особей прижаты к поверхности брюшных створок (табл. VII , фиг. 2г), что является доказательством прижизненного поселения ювенильных рихтгофеноидей. Обнаружена игла молодого животного, которая внедряется в раковинное вещество взрослой особи (Журавлев, Пахневич, 2018, рис. 2, г–з).
Характеристика сообщества. Рихтгофеноидеи обитали в условиях микробиально-водорослевого рифа. Помимо многочисленных обызвествленных талломов водорослей, вместе с этими брахиоподами встречаются и другие представители типа, в том числе, другие продуктиды, спирифериды, атиридиды (подробнее о фауне брахиопод см.: Angiolini et al., 2016). Рихтгофеноидеи ‑ самые крупные из брахиопод этих рифовых отложений, остальные брахиоподы имеют очень мелкие раковины. Эта особенность свойственна для брахиопод якорного этологического типа, обитающих на рифах и селящихся внутри рифовых каверн и полостей (Pakhnevich, 2009). Выявлены и некоторые прижизненные ассоциации рихтгофеноидей с другими организмами. Они были компонентами сообщества фильтраторов, в котором преобладали четырехлучевые кораллы, а также обитали мшанки. На одном из экземпляров хорошо видно, как раковина G. insolita срастается боковой стенкой с ругозой и водорослью. При этом иглы, встречая преграду, изгибаются вдоль раковины брахиоподы (рис. 1, и). Помимо перечисленных представителей фауны, отмечены также брюхоногие и единичные двустворчатые моллюски.
ЗАКЛЮЧЕНИЕ
Таким образом, в отложениях гундаринской свиты Дарваза разнообразие прирастающих продуктид надсемейства Richthofenioidea больше, чем оценивалось ранее. Выявлено их четыре представителя: Gundaria insolita, Hercosestria sp., Coscinarina sp. и Proteguliferina sp. Сохранность не позволяет более точно произвести определение или описать новые виды. Рихтгофеноидеи обитали в условиях микробиально-водорослевого рифа в ассоциации с другими фильтраторами. По особенностям индивидуального развития и темпов роста эти прирастающие продуктиды слабо отличались от современных якорных ринхонеллид и теребратулид, а также от других ископаемых брахиопод, относящихся к свободнолежащему, якорному, зарывающемуся, прирастающему этологическим типам.
Список литературы
Алимов А.Ф. Функциональная экология пресноводных двустворчатых моллюсков. Л.: Наука, 1981. 248 с.
Горюнова Р.В. Пермские мшанки Памира. М.: Наука, 1975. 127 с. (Тр. Палеонтол. ин-та АН СССР. Т. 148).
Грунт Т.А., Дмитриев В.Ю. Пермские брахиоподы Памира. М.: Наука, 1973. 211 с. (Тр. Палеонтол. ин-та АН СССР. Т. 136).
Журавлев В.С., Пахневич А.В. Представители надсемейства Richthofenioidea (Brachiopoda: Productida) из гундаринской свиты (пермь) Дарваза // ПАЛЕОСТРАТ-2016. Годичн. собр. (науч. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН. Москва, 26–28 января 2016 г. Тез. докл. М.: ПИН РАН, 2016. С. 32–33.
Журавлев В.С., Пахневич А.В. К вопросу о брахиопод–брахиоподовых взаимоотношениях // Палеонтол. журн. 2018. № 1. С. 28–35.
Зезина О.Н. Опыт определения темпа роста и продукции брахиопод Pelagodiscus atlanticus (King) из батиали и абиссали океана // Тр. Ин-та океанол. АН СССР. 1976. Т. 99. С. 85–90.
Золотарев В.Н. Склерохронология морских двустворчатых моллюсков. Киев: Наук. думка, 1989. 112 с.
Иванова Е.А. Условия существования, образ жизни и условия развития некоторых брахиопод среднего и верхнего карбона Подмосковной котловины. М.: Изд‑во АН СССР, 1949. 152 с. (Тр. Палеонтол. ин-та АН СССР. Т. 21).
Левен Э.Я., Леонова Т.Б., Дмитриев В.Ю. Пермь Дарваз-Заалайской зоны Памира: фузулиниды, аммоноидеи, стратиграфия. М.: Наука, 1992. 204 с. (Тр. Палеонтол. ин-та РАН. Т. 253).
Пахневич А.В. Изменчивость и популяционная экология брахиопод. Изменчивость раковин и размерно-возрастной состав популяций высокоширотных брахиопод Северного полушария. Saarbrücken: Lambert Acad. Publ., 2012. 289 с.
Чернышев Ф.Н. Фауна верхнепалеозойских отложений Дарваза // Тр. Геол. ком. Нов. сер. 1914. Вып. 104. 66 с.
Angiolini L., Campagna M., Borlenghi L. et al. Brachiopods from the Cisuralian – Guadalupian of Darvaz, Tajikistan and implications for Permian stratigraphic correlations // Palaeoworld. 2016. V. 25. Iss. 4. P. 539–568.
Angiolini L., Campagna M., Grunt T. et al. A silicified brachiopod fauna from the Cisuralian-Guadalupian Gundara Formation of Darvaz, Pamir, Tajikistan // 7th Intern. Brachiopod Congr.: The Brachiopod World (Newsletter of the Subcommission on Permian Stratigr. 2015. № 61. Suppl. 1. P. 1–2).
Cooper G.A., Grant R.E. New Permian brachiopods from West Texas // Smithson. Contrib. Paleobiol. 1969. № 1. P. 1–20.
Cooper G.A., Grant R.E. Permian brachiopods of west Texas III. Part 1 & 2 // Smithson. Contrib. Paleobiol. 1975. № 19. P. 795–1298 (part 1, text); p. 1299–1921, pl. 192–502 (part 2, plates).
Pakhnevich A.V. Reasons of micromorphism in modern and fossil brachiopods // Paleontol. J. 2009. V. 43. № 11. P. 1458–1468.
Pakhnevich A.V. Study of fossil and Recent brachiopods, using a Skyscan 1172 X-Ray microtomograph // Paleontol. J. 2010. V. 44. № 9. P. 1217–1230.
Wardlaw B., Grant R.E., Brunton C.H.C. Richthofenioidea // Treatise on Invertebrate Paleontology. Part H. Brachiopoda. Revised. V. 3. Lawrence: Univ. Kansas Press, 2000. P. 610–619.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал