

Палеонтологический журнал, 2019, № 5, стр. 3-13
Новые таксоны и новая среднефаменская экозона радиолярий Natgorella hirsuta – Spinoalius melekessensis Волго-Уральского бассейна
М. С. Афанасьева *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: afanasieva@paleo.ru
Поступила в редакцию 07.08.2018
После доработки 30.08.2018
Принята к публикации 25.02.2019
Аннотация
Описаны новые таксоны радиолярий из нижнего и среднего фамена Полярного Урала и Волго-Уральского бассейна: три новых рода Natgorella gen. nov., Spinoalius gen. nov., Diversiacus gen. nov., и три новых вида N. elegans sp. nov., N. hirsuta sp. nov. и S. melekessensis sp. nov. Новые таксоны относятся к двум отрядам, Entactiniata и Cancelliata, из двух классов сферических радиолярий Sphaerellaria и Spumellaria. Выявлено конвергентное сходство морфотипов внешних скелетов у Spinoalius gen. nov. и Diversiacus gen. nov. Установлена новая среднефаменская экозона радиолярий Natgorella hirsuta – Spinoalius melekessensis.
ИСТОРИЯ ИЗУЧЕНИЯ
В середине XX в. в отложениях нижнего и среднего фамена Волго-Уральского бассейна Е.В. Быковой (1955) были детально исследованы так называемые “примитивные” мелкие фораминиферы (рис. 1, а–р). При этом Быкова (1955, с. 110) впервые отметила определенное сходство некоторых Archaesphaera и Parathurammina с радиоляриями: “по строению раковин они ближе стоят к древним радиоляриям, чем к современным или мезозойско-кайнозойским планктонным фораминиферам”. Единственное отличие этих организмов от радиолярий, по мнению Быковой, заключается только в том, что их скелет известковый, тогда как у радиолярий он кремневый.
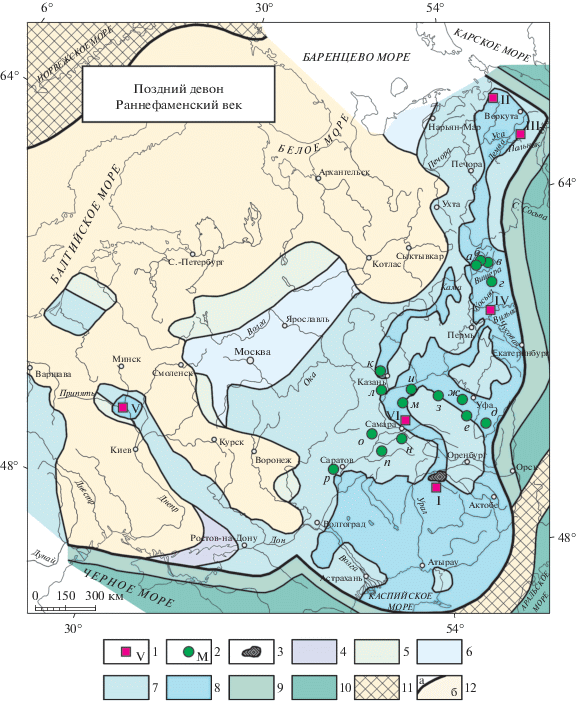
Рис. 1.
Распространение радиолярий фамена; обозначения: 1 – местонахождение экозон радиолярий: I – Прикаспийский бассейн, II – Полярный Урал, Лемвинская зона, III – Тимано-Печорский бассейн, Сарембой-Леккеягинский вал, IV – Западный склон Среднего Урала, V – Припятский прогиб, VI – Волго-Уральский бассейн (I, III – Афанасьева, 2000; II–IV – Afanasieva, Amon, 2011; Афанасьева, Амон, 2012; V – Кручек, Назаров, 1977; VI –настоящая работа); 2 – “уральский” комплекс (Быкова, 1955): а–г – Северный Урал: а – р. Петруниха, б – р. Сторожевая, в – р. Вишера, г – р. Язьва; д – Южный Урал, р. Сиказа; е–з – Западный Башкортостан: е – с. Елатминки (=Новоконстантиновка), ж – с. Каргалы, з – г. Туймазы; и–л – Татарстан: и – пос. Шугурово, к – пос. Камское Устье, л – с. Верхний Услон; м–о – Самарская обл.: м – с. Красновка, н – с. Березовка, о – г. Сызрань; п, р – Саратовская обл.: п – г. Пугачев, р – с. Тепловка; 3 – палеориф Карачаганак; 4–10 – палеобиогеографическая схема фамена Русской платформы и Большого Урала (Афанасьева, 2000; Afanasieva et al., 2011; Афанасьева, Амон, 2012): 4 – море пониженной солености; 5–10 – море нормальной солености: 5 – прибрежная область, 6 – мелкий внутренний шельф, 7 – внутренний шельф, 8 – внешний шельф, 9 – континентальный склон, 10 – дно палеоокеана; 11 – активные складчатые пояса; 12а – граница Русской платформы, 12б – суша.
Вместе с тем, известно, что в процессе литификации вмещающих отложений может происходить перекристаллизация скелетов радиолярий, в результате чего наблюдается или замещение первичного опала скелетов иными минералами (кальцитом, апатитом, флюоритом, гранатом, пиритом и золотом), или остаются только сферы, которые часто принимают за остатки примитивных мелких фораминифер или водоросли. Все сказанное выше позволяет рассматривать отдельных представителей некоторых видов из родов Archaesphaera и Parathurammina среди радиолярий (Braun, Amon, 1991; Афанасьева, 2000; Вишневская, Седаева, 2002; Седаева, Вишневская, 2002, 2008; Afanasieva, Amon, 2011; Афанасьева, Амон, 2012). Однако это мнение разделяют не все палеонтологи.
Почти через четверть века несомненные раннефаменские радиолярии с кремневым скелетом были найдены в елецком горизонте Припятского прогиба (рис. 1, V; 2, V; 3, V) и изучены Б.Б. Назаровым с помощью электронного микроскопа (Кручек, Назаров, 1977; Назаров, 1988). Позднее раннефаменские радиолярии были исследованы (Афанасьева, 2000) на Сарембой-Леккеягинском валу в Тимано-Печорском бассейне и на севере Прикаспия (рис. 1, I, III; 2, I, III; 3, I, III), и обнаружены на Полярном Урале и в бассейне р. Вильва на западном склоне Среднего Урала (рис. 1, II, IV; 2, II; 3, II, IV) (Afanasieva, Amon, 2011; Афанасьева, Амон, 2012).
На территории Волго-Уральского бассейна истинные радиолярии фамена до сих пор не были известны. В настоящее время Л.И. Кононовой при выделении конодонтов из отложений лебедянского горизонта среднего фамена, вскрытых скв. Мелекесс Р-1, обнаружены целые скелеты радиолярий прекрасной сохранности (рис. 1, VI; 2, VI; 3, VI).
Предлагаемая статья посвящена изучению впервые обнаруженных настоящих радиолярий среднего фамена Волго-Уральского бассейна и описанию новой среднефаменской экозоны радиолярий Natgorella hirsuta – Spinoalius melekessensis.
НОВАЯ ЭКОЗОНА РАДИОЛЯРИЙ NATGORELLA HIRSUTA – SPINOALIUS MELEKESSENSIS
Новая ассоциация среднефаменских радиолярий установлена в Мелекесской впадине Волго-Уральского бассейна. Радиолярии обнаружены в средней подсвите малочеремшанской свиты (инт. 1940–1980 м) на двух уровнях скв. Мелекесс Р-1 (обр. 7336, инт. 1961.9–1963.9 м; обр. 7398, инт. 1973.9–1974.7 м)11 (рис. 1, VI; 2, VI; 3, VI). Образцы дезинтегрированы в 10% растворе уксусной кислоты по стандартной методике выделения конодонтов и радиолярий. Совместно с радиоляриями в обр. 7398 Кононова обнаружила конодонты Palmatolepis glabra lepta Ziegler et Huddle, 1969. Этот подвид распространен от зоны Late crepida до зоны Late trachytera. В обр. 7336 конодонты не отмечены.
Возраст интервала с радиоляриями уточнен на основании более разнообразного комплекса конодонтов, выявленного в нижней части средней подсвиты (инт. 1973.9–1976.5 м): Palmatolepis glabra lepta Ziegler et Hudd., P. glabra pectinata Ziegler, P. gracilis gracilis Branson et Mehl, P. utahensis Ziegler et Sandberg, P. perlobata helmsi Ziegler, P. perlobata sigmoidea Ziegler, P. perlobata schindewolfi Müller, P. inflexa Müller, P. rugosa grossi Ziegler и P. marginifera marginifera Helms. Многие из этих видов и подвидов имеют широкий диапазон распространения в фаменском ярусе, но P. utahensis встречается только в зоне Late marginifera, а P. inflexa распространен от зоны Late rhomoidea до зоны Early marginifera. Данный комплекс конодонтов присутствует почти на 2 м глубже, чем обр. 7398 с радиоляриями, и характерен для зон Early marginifera и Late marginifera, соответствующих низам лебедянского горизонта среднего фамена (Родионова и др., 1995).
Таким образом, новая экозона Natgorella hirsuta – Spinoalius melekessensis характеризует часть средней подсвиты малочеремшанской свиты, отнесенной Кононовой по конодонтам к лебедянскому горизонту, которому соответствуют зоны конодонтов Late marginifera и trachytera (рис. 2, VI; 3, VI) (Фортунатова и др., 2016, 2018).
Рис. 2.
Sphaerellaria фаменского яруса верхнего девона: I–VI – экозоны радиолярий фамена: I – Tetrentactinia barysphaera – Holoeciscus auceps, Прикаспийский бассейн, палеориф Карачаганак; II – Tetrentactinia barysphaera – Caspiaza spinifera, Полярный Урал, Лемвинская зона; III – Tetrentactinia barysphaera – Retientactinosphaera magnifica, Тимано-Печорский бассейн, Сарембой-Леккеягинский вал; IV – Haplentactinia alekseevi – Haplentactinia vilvaensis, Западный склон Среднего Урала, р. Вильва; V – Tetrentactinia barysphaera – Ceratoikiscum famennium, Припятский прогиб; VI – Natgorella hirsuta – Spinoalius melekessensis, Волго-Уральский бассейн, Мелекесская впадина (I, III – Афанасьева, 2000; II–IV – Afanasieva, Amon, 2011; Афанасьева, Амон, 2012; V – Кручек, Назаров, 1977; VI – Афанасьева, настоящая работа).
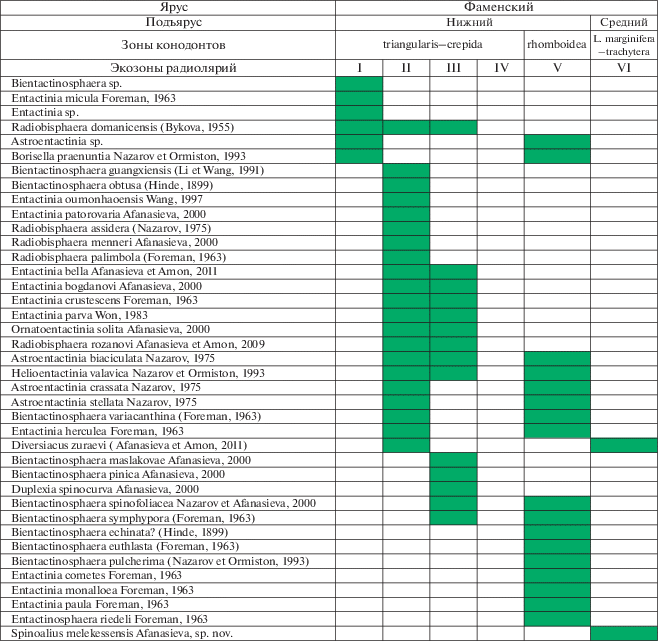
Таксономическое разнообразие новой экозоны характеризуется присутствием шести видов радиолярий (рис. 2; 3): четырех видов из трех родов решетчатых радиолярий класса Spumellaria (рис. 4, а, б; табл. I , см. вклейку) и двух видов из двух родов пористых радиолярий класса Sphaerellaria (рис. 4, в; табл. II , см. вклейку).
Рис. 3.
Spumellaria фаменского яруса верхнего девона: I–VI – экозоны радиолярий фамена (см. рис. 2).
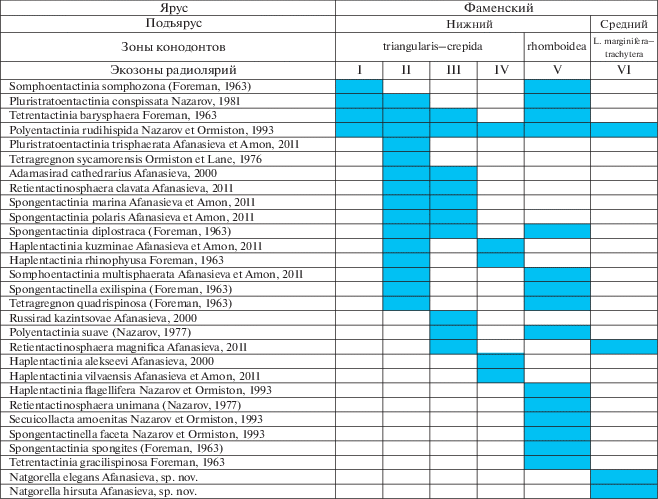
Рис. 4.
Радиолярии Мелекесской впадины: а – Retientactinosphaera magnifica Afanasieva, 2011, экз. ПИН, № 5369/187-L1015 (штрих = 131 мкм); б – Polyentactinia rudihispida Nazarov et Ormiston, 1993, экз. ПИН, № 5369/0132016-019 (штрих = 71 мкм); в – Diversiacus zuraevi (Afanasieva et Amon, 2011), экз. ПИН, № 5369/329-L1024 (штрих = 92 мкм); верхний девон, среднефаменский подъярус, лебедянский горизонт, конодонтовые зоны Late marginifera и trachytera; скв. Мелекесс Р-1: а, б – обр. 7336 (инт. 1961.9–1963.9 м); в – обр. 7398 (инт. 1973.9 –1974.7 м).
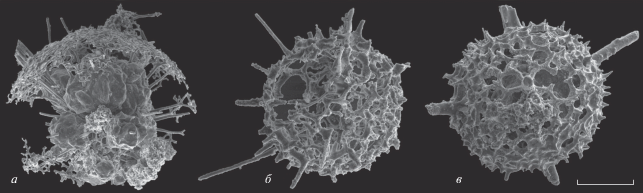
Типичными видами нового комплекса являются Natgorella elegans sp. nov. и N. hirsuta sp. nov. (табл. I ), а также Spinoalius melekessensis sp. nov. (табл. II , фиг. 1–4). Виды Retientactinosphaera magnifica Afanasieva, 2011, Polyentactinia rudihispida Nazarov et Ormiston, 1993 и Diversiacus zuraevi (Afanasieva et Amon, 2011) представлены единичными экземплярами (рис. 4).
P. rudihispida встречен во всех анализируемых местонахождениях радиолярий нижнего и среднего фамена (рис. 1, I–VI). Но впервые этот вид появился в Тимано-Печорском бассейне во франском веке (рис. 3, I–VI) (Афанасьева, 2000). Виды D. zuraevi (рис. 2, II, VI) и R. magnifica (рис. 3, III, VI) характерны только для отложений фамена (Афанасьева, 2011; Afanasieva, Amon, 2011). Вместе с тем, эти виды очень важны, поскольку являются общими между новой ассоциацией радиолярий Волго-Уральского бассейна (среднефаменский подъярус, лебедянский горизонт) и более древними нижнефаменскими экозонами22 (Afanasieva, Amon, 2011; Афанасьева, Амон, 2012) (рис. 2, 3):
Tetrentactinia barysphaera – Holoeciscus auceps, Прикаспийский бассейн Казахстана, палеориф Карачаганак, нижний фамен (Афанасьева, 1987, 2000);
Tetrentactinia barysphaera – Caspiaza spinifera, Полярный Урал, Лемвинская зона, р. Пальник-Ю, зоны конодонтов triangularis и crepida (Афанасьева, 2000; Afanasieva, Amon, 2011; Афанасьева, Амон, 2012);
Tetrentactinia barysphaera – Retientactinosphaera magnifica, Тимано-Печорский бассейн, Сарембой-Леккеягинский вал, скв. Западно-Леккеягинская-65 (инт. 2460–2467 м), зоны конодонтов triangularis и crepida (Афанасьева, 2000; Afanasieva, Amon, 2011; Афанасьева, Амон, 2012);
Haplentactinia alekseevi – Haplentactinia vilvaensis, западный склон Среднего Урала, р. Вильва, зоны конодонтов triangularis и crepida (Afanasieva, Amon, 2011; Афанасьева, Амон, 2012);
Tetrentactinia barysphaera – Ceratoikiscum famennium, Припятский прогиб Белоруссии, елецкий горизонт; совместно с радиоляриями встречены конодонты: Palmatolepis glabra Ulrich et Bassler, P. minuta Branson et Mehl., P. aff. quadrantinodoslobata Sannemann, Prioniodina smithi Stauffer, Scutula venusta Sannemann (Кручек, Назаров, 1977; Назаров, 1988), характеризующие зону romboidea;
Natgorella hirsuta – Spinoalius melekessensis, Волго-Уральский бассейн, Мелекесская впадина, лебедянский горизонт, зоны конодонтов Late marginifera и trachytera.
Ассоциация радиолярий новой экозоны удивительно похожа на комплекс экозоны Haplentactinia alekseevi – Haplentactinia vilvaensis западного склона Среднего Урала (Afanasieva, Amon, 2011; Афанасьева, Амон, 2012). Сходство выражается в общности морфотипов радиолярий, представленных в обоих случаях преимущественно ажурными решетчатыми скелетами, и в присутствии общего вида Polyentactinia rudihispida, но таксономический состав экозон разный.
Таким образом, мы рассматриваем новую среднефаменскую ассоциацию радиолярий в качестве экозоны Natgorella hirsuta – Spinoalius melekessensis, отвечающей лебедянскому горизонту среднего фамена в объеме конодонтовых зон Late marginifera и trachytera. Новая экозона радиолярий имеет район распространения и валидности на территории Волго-Уральского бассейна. Обоснование новой экозоны радиолярий имеет важное значение для стратиграфии девона региона по ряду причин. Так, это первое местонахождение истинных радиолярий среднего фамена, для которых существует контроль возраста по конодонтам. Кроме того, в ходе работ расширен диапазон стратиграфического распространения радиолярий от зоны конодонтов triangularis до зоны trachytera (рис. 2, 3). Также увеличено таксономическое и морфологическое разнообразие радиолярий в нижне–среднефаменских экозонах, которые отличаются друг от друга по таксономическому составу, но вместе с тем характеризуются преобладанием ажурных представителей из класса Spumellaria и имеют общие виды Retientactinosphaera magnifica и Polyentactinia rudihispida.
КОНВЕРГЕНЦИЯ
В биологии понятие конвергенция было введено немецким зоологом Э.О. Шмидтом для анализа схождения или параллелизма отдельных морфологических признаков или суммы признаков для таксономических категорий любого ранга (Майр, 1974). Под конвергенцией понимается “процесс независимого приобретения сходства членами двух или нескольких филетических линий в связи со сходством образа жизни, но вне связи с их родством” (Палеонтология…, 1995, с. 165).
Все конвергентные признаки являются вторичными образованиями, возникшими в процессе развития самостоятельно и независимо в порядке приспособления к условиям обитания. При этом конвергентное сходство морфотипов разных таксонов не бывает глубоким и затрагивает только внешние морфологические признаки. Вот почему в случаях конвергенции даже внутри одной и той же систематической группы конвергентные признаки никогда не являются точным повторением признаков других организмов. Таким образом, совпадение внешней морфологии сравниваемых таксонов обычно частичное и относительное, а не абсолютно идентичное.
Исследование скелетов новой ассоциации радиолярий выявило конвергентное морфологическое сходство морфотипов внешнего облика скелетов новых родов Spinoalius gen. nov. и Diversiacus gen. nov. (рис. 5). Конвергентное сходство этих родов основано на наборе общих признаков: (1) внешняя пористая сфера, (2) шесть основных игл, из которых пять игл являются стержневидными, а одна – граненая (четырехгранная или трехгранная) (табл. II ), (3) обрамление основания стержневидных игл шестью порами лепестковидной формы (табл. II , фиг. 1а, 1б; 7).
Рис. 5.
Конвергентное сходство морфотипов внешних скелетов радиолярий: а – Spinoalius, б – Diversiacus.
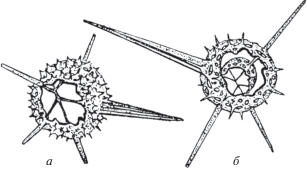
Различия касаются внутреннего строения скелетов, что является признаками подсемейств (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006): род Spinoalius характеризуется наличием только одной внешней сферы (подсемейство Entactiniinae) (рис. 5, а; табл. II , фиг. 2а), а род Diversiacus отличается двумя сферами – внешней и внутренней (подсемейство Bientactinosphaerinae) (рис. 5, б; табл. II , фиг. 5, 6, 8 ).
Рис. 6.
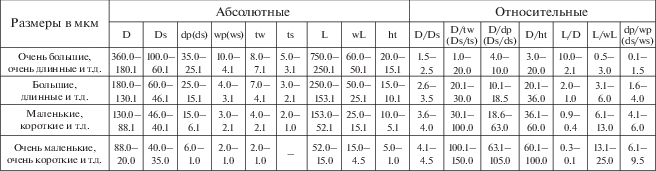
Основные абсолютные и относительные морфометрические параметры скелетов сферических радиолярий палеозоя (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006): D – диаметр внешней сферы, Ds – диаметр первичной сферы, dp – диаметр пор внешней оболочки, ds – диаметр пор первичной сферы, wp – ширина межпоровой перегородки внешней оболочки, ws – ширина межпоровой перегородки пор первичной сферы, tw – толщина стенки внешней оболочки, ts – толщина стенки первичной сферы, L – длина основных игл, wL – ширина основания основных игл, ht – высота шипиков.
Таким образом, в двух подсемействах, Entactiniinae и Bientactinosphaerinae, параллельно во времени и независимо друг от друга сформировались похожие внешние скелеты, что может свидетельствовать о сходных условиях существования и одинаково направленном естественном отборе. Эта разница в строении скелетов позволяет установить два новых рода: Spinoalius gen. nov. и Diversiacus gen. nov.
Коллекции радиолярий фаменского яруса верхнего девона №№ 5312, 5369 хранятся в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН). Систематика радиолярий приведена по работам автора (Афанасьева, Амон, 2003, 2006; Afanasieva et al., 2005). Измерения элементов скелетов радиолярий и морфометрический анализ абсолютных и относительных значений параметров раковин радиолярий палеозоя проводился по стандартной методике (рис. 6) (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006).
Автор приносит свою искреннюю благодарность Л.И. Кононовой за предоставленный материал по радиоляриям и за консультации относительно возраста вмещающих пород; А.С. Алексееву и В.С. Вишневской за плодотворное обсуждение статьи и конструктивные рекомендации; А.Ф. Банникову и Н.М. Кузьминой за ценные советы и замечания при подготовке статьи к печати. Работа выполнена при поддержке Программы фундаментальных исследований президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов”.
ОПИСАНИЕ ТАКСОНОВ
ТИП RADIOLARIA
НАДКЛАСС POLYCYSTINA
КЛАСС SPUMELLARIA
ОТРЯД CANCELLIATA
СЕМЕЙСТВО HAPLENTACTINIIDAE NAZAROV IN NAZAROV ET POPOV, 1980
ПОДСЕМЕЙСТВО PSEUDOROTASPHAERINAE NOBLE, 1994
Род Natgorella Afanasieva, gen. nov.
Название рода в честь палеонтолога и стратиграфа Натальи Валерьевны Горевой; ж. р.
Типовой вид – Natgorella hirsuta sp. nov.
Диагноз. Раковина сферическая с двумя решетчатыми оболочками, образованными переплетением округлых прутьев. Продолжением лучей внутренней 6-лучевой спикулы являются шесть основных стержневидных игл с округлым поперечным сечением. На поверхности прутьев внешней сферы развиты шипики двух типов: конические и тонкие стержневидные, раздваивающиеся (бифуркация) примерно в верхней трети своей длины.
Видовой состав. Типовой вид и N. elegans sp. nov.
Сравнение. От родов Cancellosphaera Afanasieva, 2000, Russirad Afanasieva, 2000 и Arcoclathrata Afanasieva et Amon, 2016, которые характеризуются внешней решетчатой и внутренней пористой оболочками скелета и наличием шести основных трехгранных игл, новый род отличается двумя решетчатыми оболочками скелета и шестью основными стержневидными иглами. От рода Retientactinosphaera Afanasieva, 2011, у которого развита только одна основная игла трехгранной формы, новый род отличается наличием шести основных стержневидных игл.
Natgorella hirsuta Afanasieva, sp. nov.
Табл. I , фиг. 4–6
Название вида от hirsutus лат. – взъерошенный, мохнатый.
Голотип – ПИН, № 5369/327-М1021; Волго-Уральский бассейн, Мелекесская впадина, скв. Мелекесс Р-1 (инт. 1973.9–1974.7 м); верхний девон, среднефаменский подъярус, лебедянский горизонт, зоны конодонтов Late marginifera и trachytera.
Описание. Раковина сферическая, очень большая (D = 231–248 мкм), состоящая из двух решетчатых оболочек с шестью основными стержневидными иглами и многочисленными шипиками. Внешняя оболочка состоит из переплетения тонких прутьев (D/tw = 34.4–42.8), образующих очень крупные (D/dp = 8.3–10.2) полигональные поровые отверстия примерно одинакового размера. Внутренняя сфера большая (D/Ds = 3.1–3.3). относительно тонкостенная (Ds/ts = 23.3–25.7) и составляет 30.3–31.9% от диаметра внешней сферы. Она сформирована переплетением прутьев. обрамляющих очень крупные (Ds/ds = 5.9) полигональные поровые отверстия. Продолжением лучей шестилучевой спикулы являются шесть относительно коротких (L/D = 0.7–1.0) и тонких (L/wL = 7.8–13.0) основных стержневидных игл. На поверхности прутьев внешней сферы развиты шипики двух типов: (1) малочисленные, но относительно длинные (D/ht1 = 15.4–20.1) конические шипики; (2) очень многочисленные и очень длинные (D/ht2 = 2.3–3.4) стержневидные тонкие шипики, разветвляющиеся (двойная бифуркация) примерно на середине своей длины. Длина шипиков составляет 29.1–44.4% от диаметра внешней сферы, а двойная бифуркация многочисленных тонких шипиков (табл. I , фиг. 5б) создает иллюзию взъерошенного, мохнатого создания, что отражено в названии вида.
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | tw | Ds | ds | ts | L | wL | ht1 | ht2 |
|---|---|---|---|---|---|---|---|---|---|---|
| 5369/327-М1016 (голотип) |
248 | 21 | 5 | – | – | – | 245 | 20 | 14 | 110 |
| 5369/327-М1017 | 241 | 29 | 6 | 77 | 13 | 3 | 221 | 17 | 12 | 70 |
| 5369/327-М1023 | 231 | 23 | 6 | 70 | – | 3 | 155 | 20 | 15 | 73 |
| Экз. № | D/tw | D/dp | L/D | L/wL | D/ht1 | D/ht2 | D/Ds | Ds/ds | Ds/ts |
|---|---|---|---|---|---|---|---|---|---|
| 5369/327-М1016 (голотип) |
42.8 | 10.2 | 1.0 | 12.3 | 17.7 | 2.3 | – | – | – |
| 5369/327-М1017 | 34.4 | 8.3 | 0.9 | 13.0 | 20.1 | 3.4 | 3.1 | 5.9 | 25.7 |
| 5369/327-М1023 | 38.5 | 10.0 | 0.7 | 7.8 | 15.4 | 3.2 | 3.3 | – | 23.3 |
Сравнение. От N. elegans sp. nov., характеризующегося малочисленными тонкими стержневидными шипиками, только иногда раздваивающимися в верхней трети своей длины, и многочисленными очень маленькими коническими шипиками на поверхности внутренней сферы, отличается наличием очень многочисленных тонких стержневидных шипиков, часто дважды разветвляющихся.
Материал. 3 экз. из типового местонахождения.
Natgorella elegans Afanasieva, sp. nov.
Табл. I , фиг. 1–3
Название вида elegans лат. – изящный.
Голотип – ПИН, № 5369/328-М1014; Волго-Уральский бассейн, Мелекесская впадина, скв. Мелекесс Р-1 (инт. 1961.9–1963.9 м); верхний девон, среднефаменский подъярус, лебедянский горизонт, зоны конодонтов Late marginifera и trachytera.
Описание. Раковина сферическая, очень крупная (D = 206–219 мкм), состоящая из двух решетчатых оболочек с шестью основными стержневидными иглами и многочисленными шипиками. Внешняя оболочка состоит из переплетения тонких прутьев (D/tw =36.5–42.0), образующих очень крупные (D/dp = 7.5–10.3) полигональные поровые отверстия примерно одинакового размера. Внутренняя сфера большая (D/Ds = 2.8–3.2), относительно толстостенная (Ds/ts = 22.7–25.0), и составляет 31.1–35.9% от диаметра внешней сферы. Она сформирована переплетением прутьев, обрамляющих очень крупные (Ds/ds = 5.7–6.8) полигональные поровые отверстия. На поверхности прутьев внутренней сферы обнаружены многочисленные очень маленькие конические шипики высотой от 2 до 5 мкм (табл. I , фиг. 3б). Шесть основных стержневидных игл являются продолжением внутренней шестилучевой спикулы: иглы относительно короткие (L/D = 0.5–0.9) и тонкие (L/wL = 10.2–13.4). На поверхности прутьев внешней сферы имеются шипики двух типов: (1) очень многочисленные длинные (D/ht1 = 12.2–13.7) конические шипики; (2) малочисленные. но очень длинные (D/ht2 = = 3.8–4.3) стержневидные тонкие шипики, иногда раздваивающиеся (бифуркация) примерно в верхней трети своей длины.
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | tw | Ds | ds | ts | L | wL | ht1 | ht2 |
|---|---|---|---|---|---|---|---|---|---|---|
| 5369/328-М1014 (голотип) | 210 | 28 | 5 | 75 | 11 | 3 | 201 | 15 | 17 | 50 |
| 5369/328-М1020 | 219 | 23 | 5 | 68 | 12 | 3 | – | 11 | 18 | 58 |
| 5369/328-М1021 | 206 | 20 | 5 | 74 | 13 | 3 | 102 | 10 | 15 | 48 |
| Экз. № | D/tw | D/dp | L/D | L/wL | D/ht1 | D/ht2 | D/Ds | Ds/ds | Ds/ts |
|---|---|---|---|---|---|---|---|---|---|
| 5369/328-М1014 (голотип) | 42.0 | 7.5 | 0.9 | 13.4 | 12.4 | 4.2 | 2.8 | 6.8 | 25.0 |
| 5369/328-М1020 | 36.5 | 9.5 | – | – | 12.2 | 3.8 | 3.2 | 5.7 | 22.7 |
| 5369/327-М1021 | 41.2 | 10.3 | 0.5 | 10.2 | 13.7 | 4.3 | 2.8 | 5.7 | 24.7 |
Материал. 3 экз. из типового местонахождения.
КЛАСС SPHAERELLARIA
ОТРЯД ENTACTINIATA
СЕМЕЙСТВО ENTACTINIIDAE RIEDEL, 1967
ПОДСЕМЕЙСТВО ENTACTINIINAE RIEDEL, 1967
Род Spinoalius Afanasieva, gen. nov.
Название рода от spina лат. – игла и alius лат. – разный, различный; м. р.
Типовой вид – Spinoalius melekessensis sp. nov.
Диагноз. Раковина сферическая, пористая. Продолжением лучей внутренней шестилучевой
спикулы являются шесть основных игл: одна доминирующая четырехгранная игла с  -образным сечением основания и пять стержневидных игл с округлым сечением основания.
Около основания стержневидных игл присутствуют шесть поровых отверстий лепестковидной
формы (табл. II
, фиг. 1а, 1б).
-образным сечением основания и пять стержневидных игл с округлым сечением основания.
Около основания стержневидных игл присутствуют шесть поровых отверстий лепестковидной
формы (табл. II
, фиг. 1а, 1б).
Видовой состав. Типовой вид.
Сравнение. От наиболее близкого рода Borisella Afanasieva, 2000, который характеризуется
пористым скелетом с шестью основными стержневидными иглами, новый род отличается наличием
одной доминирующей четырехгранной иглы с  -образным поперечным сечением и пяти стержневидных игл.
-образным поперечным сечением и пяти стержневидных игл.
Spinoalius melekessensis Afanasieva, sp. nov.
Табл. II , фиг. 1–4
Название вида по местонахождению в Мелекесской впадине Волго-Уральского бассейна.
Голотип – ПИН, № 5369/329-М1027; Волго-Уральский бассейн, Мелекесская впадина, скв. Мелекесс Р-1, инт. 1973.9 –1974.7 м; верхний девон, среднефаменский подъярус, лебедянский горизонт, зоны конодонтов Late marginifera и trachytera.
Описание. Раковина сферическая, очень крупная (D = 214–252 мкм), состоящая из одной
пористой оболочки. Внешняя сфера толстостенная (D/tw = 19.4–25.6) с округлыми, большими
порами примерно одного размера (D/dp = 12.2–14.4). Межпоровые перегородки приостренные
и относительно узкие (dp/wp = 4.5–5.7). Продолжением лучей внутренней шестилучевой
спикулы являются шесть основных игл: (1) одна доминирующая четырехгранная игла с  -образным поперечным сечением, относительно длинная (L1/D = 0.7–1.1) и тонкая (L1/wL1 = 7.3–11.7); (2) пять стержневидных игл с округлым поперечным сечением, иглы короткие
(L2/D = 0.4–0.6) и относительно толстые (L2/wL2 = 3.9–6.3). Около основания стержневидных игл присутствуют шесть поровых отверстий
лепестковидной формы (табл. II
, фиг. 1а, б). Межпоровые перегородки осложнены скульптурными элементами в форме очень
высоких приостренных шипиков, иногда бифуркирующих в дистальной части (табл. II
, фиг. 3, 4).
-образным поперечным сечением, относительно длинная (L1/D = 0.7–1.1) и тонкая (L1/wL1 = 7.3–11.7); (2) пять стержневидных игл с округлым поперечным сечением, иглы короткие
(L2/D = 0.4–0.6) и относительно толстые (L2/wL2 = 3.9–6.3). Около основания стержневидных игл присутствуют шесть поровых отверстий
лепестковидной формы (табл. II
, фиг. 1а, б). Межпоровые перегородки осложнены скульптурными элементами в форме очень
высоких приостренных шипиков, иногда бифуркирующих в дистальной части (табл. II
, фиг. 3, 4).
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | wp | tw | L1 | wL1 | L2 | wL2 | ht |
|---|---|---|---|---|---|---|---|---|---|
| 5369/329-М1027 (голотип) | 230 | 16 | 3 | 9 | 363 | 31 | 86 | 22 | 54 |
| 5369/329-М1022 | 231 | 19 | 4 | 11 | 220 | 30 | 136 | 28 | 52 |
| 5369/329-М1025 | 252 | 18 | 4 | 13 | – | 32 | 125 | 20 | 30 |
| 5369/329-М1026 | 214 | 17 | 3 | 9 | – | 27 | 102 | 18 | 40 |
| Экз. № | D/dp | D/tw | dp/wp | L1/D | L1/wL1 | L2/D | L2/wL2 | D/ht |
|---|---|---|---|---|---|---|---|---|
| 5369/329-М1027 (голотип) | 14.4 | 25.6 | 5.3 | 1.6 | 11.7 | 0.4 | 3.9 | 4.3 |
| 5369/329-М1022 | 12.2 | 21.0 | 4.8 | 1.0 | 7.3 | 0.6 | 4.9 | 4.4 |
| 5369/329-М1025 | 14.0 | 19.4 | 4.5 | – | – | 0.5 | 6.3 | 8.4 |
| 5369/329-М1026 | 12.6 | 23.8 | 5.7 | – | – | 0.5 | 5.7 | 5.4 |
Замечание. От видов наиболее близкого рода Borisella Afanasieva, 2000, характеризующегося
шестью стержневидными иглами, новый вид отличается наличием одной доминирующей четырехгранной
иглы с  -образным поперечным сечением и пяти стержневидных игл.
-образным поперечным сечением и пяти стержневидных игл.
Материал. Четыре экз. из типового местонахождения.
ПОДСЕМЕЙСТВО BIENTACTINOSPHAERINAE AFANASIEVA, 1999
Род Diversiacus Afanasieva, gen. nov.
Название вида от diversus лат. – разнообразный, и acus лат. – игла; м.р.
Типовой вид – Bientactinosphaera zuraevi Afanasieva et Amon, 2011 из верхнего девона, нижнефаменского подъяруса, конодонтовых зон triangularis и crepida Полярного Урала, и среднефаменского подъяруса, лебедянского горизонта, конодонтовых зон Late marginifera и trachytera Волго-Уральского бассейна.
Диагноз. Раковина сферическая с двумя пористыми оболочками. Внутренний каркас в форме
6-лучевой спикулы (табл. II
, фиг. 8
), продолжением лучей которой являются 6 основных игл. Одна основная игла имеет трехгранную
форму с 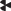 -образным поперечным сечением иглы и лопастьевидным сечением каждой грани. Пять основных
игл стержневидные с округлым поперечным сечением. Около основания стержневидных игл
формируются шесть поровых отверстий лепестковидной формы (табл. II
, фиг. 7
).
-образным поперечным сечением иглы и лопастьевидным сечением каждой грани. Пять основных
игл стержневидные с округлым поперечным сечением. Около основания стержневидных игл
формируются шесть поровых отверстий лепестковидной формы (табл. II
, фиг. 7
).
Видовой состав. Типовой вид.
Сравнение. От рода Bientactinosphaera Afanasieva, 2000, характеризующегося шестью основными трехгранными иглами, новый род отличается наличием одной основной иглы трехгранной формы и пятью основными стержневидными иглами.
Замечания. Изначально вид D. zuraevi был описан в составе рода Bientactinosphaera на основании наличия двух пористых сферических оболочек скелета (Afanasieva, Amon, 2011). Однако особенности морфологии скелета позволили пересмотреть таксономическую позицию вида и установить на его основе новый род Diversiacus gen. nov. Кроме этого, установлено конвергентное сходство внешнего облика скелетов новых родов Diversiacus и Spinoalius (рис. 5). Скелеты этих родов характеризуются наличием внешней пористой сферы с пятью основными стержневидными иглами и одной основной граненой иглой (четырехгранной или трехгранной) (табл. II ). При этом скелеты рода Spinoalius характеризуются (рис. 5, а; табл. II , фиг. 2а) только одной внешней сферой (подсем. Entactiniinae), а Diversiacus отличается (рис. 5, б; табл. II , фиг. 5, 6, 8 ) двумя сферами: внешней и внутренней (подсем. Bientactinosphaerinae).
Список литературы
Афанасьева М.С. Позднепалеозойские радиолярии месторождения Карачаганак и их фациальная приуроченность // Стратиграфия и палеонтология Прикаспийской впадины / Ред. Ильин В.Д., Замилацкая Т.К. М.: ВНИГНИ, 1987. С. 26–47.
Афанасьева М.С. Атлас радиолярий палеозоя Русской платформы. М.: Научный Мир, 2000. 480 с.
Афанасьева М.С. Систематическое положение рода Entactinosphaera Foreman, 1963 и новый род Retientactinosphaera gen. nov. (радиолярии палеозоя) // Палеонтол. журн. 2011. № 2. С. 3–13.
Афанасьева М.С., Амон Э.О. Новая классификация радиолярий // Палеонтол. журн. 2003. № 6. С. 72–86.
Афанасьева М.С., Амон Э.О. Радиолярии. М.: ПИН РАН, 2006. 320 с.
Афанасьева М.С., Амон Э.О. Биостратиграфия и палеобиогеография радиолярий девона России. М.: ПИН РАН, 2012. 280 с.
Быкова Е.В. Фораминиферы и радиолярии девона Волго-Уральской области и Центрального девонского поля и их значение для стратиграфии // Фораминиферы, радиолярии и остракоды девона Волго-Уральской области. Л.: Гостоптехиздат, 1955. С. 5–190 (Тр. ВНИГРИ. Нов. сер. Вып. 87).
Вишневская В.С., Седаева К.М. Ревизия некоторых таксонов фораминифер отряда Parathuramminoidea и вопросы эволюции фораминифер и радиолярий // Палеонтол. журн. 2002. № 6. С. 15–24.
Кручек С.А., Назаров Б.Б. Верхнедевонские радиолярии Русской платформы // Докл. АН СССР. 1977. Т. 237. № 6. С. 1445–1448.
Майр Э. Популяции, виды и эволюция. М.: Мир, 1974. 460 с.
Назаров Б.Б. Радиолярии палеозоя. Л.: Недра, 1988. 231 с. (Практическое руководство по микрофауне СССР. Т. 2).
Палеонтология и палеоэкология: Словарь-справочник / Ред. Макридин В.П., Барсков И.С. М.: Недра, 1995. 494 с.
Родионова Г.Д., Умнова В.Т., Кононова Л.И. и др. Девон Воронежской антеклизы и Московской синеклизы. М.: ЦРГЦ, 1995. 265 с.
Седаева Г.М., Вишневская В.С. Ревизия надсемейства Parathuramminoidae – фораминиферы или радиолярии // Вестн. МГУ. Сер. 4. Геол. 2002. № 5. С. 15–19.
Седаева К.М., Вишневская В.С. О природе проблематичных микрофоссилий из верхнего девона и нижнего карбона // Геол. и геофизика. 2008. Т. 49. Нов. палеонтол. стратигр. Вып. 10–11. С. 152–152.
Стратиграфический кодекс России. Изд. 3-е. СПб.: Изд-во ВСЕГЕИ, 2006. 96 с.
Фортунатова Н.К., Зайцева Е.Л., Бушуева М.А. и др. Стратиграфия верхнего девона Волго-Уральского субрегиона (по материалам актуализации стратиграфических схем). М.: ВНИГНИ, 2016. 176 с.
Фортунатова Н.К., Зайцева Е.Л., Кононова Л.И. и др. Литолого-фациальная и биостратиграфическая характеристика верхнедевонских отложений опорной скважины Мелекесс Р-1 (Мелекесская впадина, Волго-Уральский субрегион) // Бюлл. МОИП. Отд. геол. 2018. Т. 93. Вып. 5–6. С. 3–49.
Afanasieva M.S., Amon E.O. Devonian radiolarians of Russia // Paleontol. J. 2011. V. 45. № 11. P. 1313–1532.
Afanasieva M.S., Amon E.O., Agarkov Yu.V., Boltovskoy D.S. Radiolarians in the geological record // Paleontol. J. 2005. V. 39. Suppl. 3. P. 135–392.
Braun A., Amon E.O. Fluoritisierte Radiolarien aus Kieselkalk-Banken des Mittel–Viseum (Unterkarbon) des Rheinischen Schiefergebirges (Deutschland) // Paläontol. Z. 1991. Bd 65. № 1–2. S. 25–33.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал