

Палеонтологический журнал, 2019, № 5, стр. 14-21
Cephalonega, новое родовое название и система вендских проартикулят
А. Ю. Иванцов a, *, М. А. Федонкин a, А. Л. Наговицын b, М. А. Закревская a
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Архангельский краеведческий музей
Архангельск, Россия
* E-mail: ivancov@paleo.ru
Поступила в редакцию 29.12.2017
После доработки 09.11.2018
Принята к публикации 14.02.2019
Аннотация
В связи с омонимией для рода вендских ископаемых организмов Onega Fedonkin, 1976 предложено новое замещающее название Cephalonega Fedonkin, nom. nov. Приводится уточненный диагноз рода, обоснована его принадлежность к вымершему типу многоклеточных животных Proarticulata. Впервые дана развернутая характеристика типа и всех классов проартикулят.
ВВЕДЕНИЕ
Первое крупное местонахождение окаменелостей эдиакарского типа в естественных обнажениях верхнего венда на северо-западе Восточно-Европейской платформы было обнаружено на Онежском полуострове по берегам р. Сюзьмы в 1972 г. (Келлер и др., 1974). Уже на начальных этапах изучения сюзьминского местонахождения было найдено несколько отпечатков небольшого организма, описанного в качестве монотипического рода Onega Fedonkin, 1976 (Келлер, Федонкин, 1976). Малые размеры, наличие признаков сегментации и, в то же время, наблюдаемая простота строения позволили считать это существо ювенильной формой членистых животных: членистоногих (Келлер, Федонкин, 1976; Glaessner, 1979) или аннелид (Федонкин, 1981). В дальнейшем М.А. Федонкин отнес Onega к проартикулятам – установленному им типу многоклеточных животных, вымерших, по-видимому, еще в докембрии (Федонкин, 1985). Открытие и изучение новых местонахождений макрофоссилий в Юго-Восточном Беломорье, в том числе крупнейшего в мире Зимнегорского, надолго отвлекло внимание исследователей от сюзьминских находок. Только в начале текущего столетия изучение новых материалов, полученных в результате масштабных поисковых работ, позволило уточнить морфологию организма и расширить известный диапазон его стратиграфического распространения (Иванцов, 2007). Тогда же выяснилось, что название Onega преоккупировано родом современных южноамериканских цикадок семейства Cicadellidae – Onega Distant, 1908 (Takiya, Cavichioli, 2004).
В настоящей статье дается новое родовое название для сюзьминского ископаемого организма, модифицируются диагноз рода и описание вида. В целях обоснования систематического положения таксона сформулированы уточненные характеристики типа Proarticulata и подчиненных таксонов высокого ранга.
ОБСУЖДЕНИЕ
Тип Proarticulata выделен в составе вендской фауны на основе изучения билатерально-симметричных животных, обладающих симметрией скользящего отражения и первоначально объединенных в классы Vendiamorpha и Dipleurozoa (Федонкин, 1985). В настоящей работе мы обосновываем выделение третьего класса проартикулят – Cephalozoa. За прошедшие годы, благодаря систематическим раскопкам и детальному исследованию представительного ископаемого материала (Иванцов, 2007, 2008, 2011), проартикуляты обрели статус группы вендских беспозвоночных наибольшего видового разнообразия, хотя их распространение ограничено только верхним вендом Восточной Европы (включая Урал) и эдиакарием Южной Австралии. Dickinsonia Sprigg, 1947, одно из классических ископаемых неопротерозоя, описанию и интерпретации которого посвящено множество статей, является типичным представителем проартикулят.
Проартикуляты обладали низким и широким метамерным телом, но не имели боковых или вентральных придатков, способных выполнять функции конечностей. Известны следы питания проартикулят, позволяющие считать их многоклеточными животными, способными к активному передвижению и питанию (Иванцов, Малаховская, 2002; Fedonkin, 2002; Gehling et al., 2005; Иванцов, 2011). Одной из характерных черт организации проартикулят является их “дочленистость” – разделение тела на два ряда “полусегментов” (изомеров), смещенных друг относительно друга по обе стороны сагиттальной плоскости (Федонкин, 1985; Иванцов, 2008). На ископаемых остатках проартикулят всех известных видов чередующееся расположение изомеров (симметрия скользящего отражения) обычно выражено достаточно четко, но иногда встречаются экземпляры, у которых изомеры расположены супротивно. В качестве примера этого приводится ювенильная форма Dickinsonia costata (Gehling et al., 2005; Evans et al., 2017; Hoekzema et al., 2017; Dunn et al., 2018). Следует отметить, что супротивное расположение наблюдается, и то далеко не всегда, у небольших экземпляров, обладающих тонкими изомерами. У проартикулят, имеющих широкие изомеры, случаев его не известно. Тем не менее, некоторые исследователи именно супротивное расположение считают исходным и описывают дикинсоний, соответственно, как нормально сегментированные организмы (Glaessner, Wade, 1966; Wade, 1972; Jenkins, 1992; Gehling et al., 2005; Evans et al., 2017; Hoekzema et al., 2017; Dunn et al., 2018). Здесь мы приводим изображения большого и маленького экземпляров одного из видов дикинсоний, обладающих тонкими изомерами, чередующееся расположение которых видно вполне явственно (табл. III , фиг. 2, 3 ). Четкость и правильность отпечатков, на наш взгляд, свидетельствует об отсутствии деформаций тела и, соответственно, о первичности именно такого расположения изомеров. Небольшого посмертного продольного сдвига той или другой стороны тела было бы достаточно для того, чтобы у маленького образца (таблица III , фиг. 3 ) изомеры встали супротивно (или наоборот). Однако трудно объяснить сдвигом отсутствие непрерывности гребней, отмечающих границы между изомерами, наблюдаемое на оси тела крупного образца (табл. III , фиг. 2). Конечно, демонстрацией все новых и новых экземпляров доказать первичность какого-либо положения трудно. Но сама возможность разделения на отпечатках «сегментов» на две части строго по оси тела и смещение этих половин друг относительно друга, чем бы они ни были вызваны, свидетельствует о наличии в теле животного некоей продольной структуры: мембраны, тяжа или другой линейной зоны, с которой смыкаются проксимальные концы изомеров. Присутствие узкой продольной преграды, делящей тело на две достаточно независимые половины, противоречит плану строения фанерозойских членистых животных и не отмечается на их ископаемых остатках. Таким образом, тип Proarticulata может быть противопоставлен всем остальным одиночным сегментированным многоклеточным животным.
Объяснение к таблице III
Проартикуляты из отложений верхнего венда Юго-Восточного Беломорья; фиг. 1–5 – естественные отпечатки, фиг. 6–10 – латексные слепки; длина масштабного отрезка: фиг. 1–3 – 1 см, фиг. 5–9 – 0.5 см, фиг. 10 – 0.3 см.
Фиг. 1–4 . Dickinsonia cf. menneri (Keller, 1976): 1 – экз. ПИН, № 4716/5148, один из самых крупных экземпляров вида; 2 – экз. ПИН, № 4716/5165, деталь, демонстрирующая характерное для проартикулят чередующееся расположение изомеров; 3 – экз. ПИН, № 4716/5150, экземпляр среднего размера (большая величина головного отдела сравнительно с толщиной изомеров, долго сохраняющаяся в онтогенезе – диагностический признак вида); 4 – экз. ПИН, № 4716/5146, экземпляр с прижизненным повреждением заднего конца тела, приведшим к расщеплению зоны образования новых изомеров и последующему росту в двух направлениях: 4а – общий вид, 4б – деталь; Лямицкий берег, Лямицкое местонахождение, локальное скопление L2(XII); усть-пинежская свита, архангельские слои.
Фиг. 5–10 . Cephalonega stepanovi (Fedonkin, 1976): 5 – голотип ПИН, № 3992/5049; 6 – экз. ПИН, № 3992/5005 (стрелка указывает на нитевидный вырост); 7 – экз. ПИН, № 3992/5041; р. Сюзьма, местонахождение Сюзьминское, локальное скопление SZ1(XX); сюзьминские слои; 8 – экз. ПИН, № 3993/6795; 9 – экз. ПИН, № 3993/6397; Зимний берег Белого моря, местонахождение Зимнегорское, локальное скопление Z11(XXII); зимнегорские слои; 10 – экз. ПИН, № 4853/600; р. Солза, местонахождение Солзинское, локальное скопление SL1(VII); сюзьминские слои.

Расчлененная структура, формирующая характерный отпечаток проартикулят, по-видимому, представляет лишь часть тела организма. Это тонкостенное образование облекало сверху и снизу внутренние области тела (Ivantsov et al., 2019) и состояло из вещества, более плотного и устойчивого к разложению, чем другие материалы, составлявшие тело проартикулят (Bobrovskiy et al., 2019). Снаружи она была покрыта тканью иного рода, на спинной стороне тела формировавшей щит, иногда не расчлененный. Внешняя поверхность щита несла бугорки и иные выросты. Предполагается, что расчлененная структура представляла собой базальную мембрану, а располагавшееся поверх нее образование – эктодермальные покровы (Ivantsov et al., 2019).
По мнению Е. Дзика, у Dipleurozoa (таксон высокого ранга, помимо Dickinsonia, включающий в себя также несколько родов Cephalozoa предлагаемой здесь системы) внутри тела над несущим выросты прямым кишечным трактом располагалось особое метамерное образование, состоявшее из серии мускульных камер. В анатомическом строении диплеврозой Дзик прослеживает гомологии с немертинами и некоторыми вторичноротыми (Dzik, Ivantsov, 1999; Dzik, Martyshyn, 2015). Однако дорсальное расположение метамерного образования не подтверждается ископаемым материалом, а предложенная в указанных работах интерпретация некоторых слабо выраженных на отпечатках структур, как, например, мускулистой глотки или сериально расположенных гонад, кажется слабо обоснованной.
В противоположность таким представлениям, Федонкин (Fedonkin, 2002) интерпретировал метамерную структуру Dickinsonia, Ovatoscutum и Andiva как ребристый, тонкий и гибкий дорсальный покров или панцирь, что оставляет открытым вопрос о расчлененности нижележащей части тела проартикулят. Однако нам известны следы питания некоторых видов, в том числе D. costata, которые представляют собой, по сути, отпечатки вентральной поверхности этих организмов. Судя по следам, верхняя и нижняя стороны проартикулят были расчленены одинаковым образом (Иванцов, 2011).
Рост тела в онтогенезе проартикулят реализовывался двумя процессами: приращением количества изомеров (или их пар, которые отдельные авторы считают сегментами) и увеличением их линейных размеров (Runnegar, 1982; Ivantsov, 2004; Hoekzema et al., 2017). Обычно считается, что зона роста располагалась на приостренном конце тела проартикуляты, где происходило добавление новых маленьких изомеров (Runnegar, 1982; Sperling, Vinther, 2010; Gold et al., 2015; Evans et al., 2017). По альтернативной гипотезе зона роста у дикинсоний находилась между непарной лопастью и остальным телом, и новые изомеры добавлялись путем вставки (Hoekzema et al., 2017; Dunn et al., 2018). Приостренный конец, несущий маленькие изомеры, обычно интерпретируется как задний (Glaessner, Wade, 1966; Wade, 1972; Runnegar, 1982), но определенным это стало лишь после находок ориентированных следов перемещения и питания животного вместе с отпечатками тела (Иванцов, Малаховская, 2002; Gehling et al., 2005; Иванцов, 2011). В нашей коллекции имеется экземпляр D. cf. menneri, у которого впервые наблюдается интересное тератологическое явление – прижизненное расщепление оси тела надвое с формированием дополнительной линии чередующихся изомеров (табл. III , фиг. 4). Данный экземпляр подтверждает предположение о заднем терминальном расположении зоны роста у проартикулят. На противоположном (переднем) конце тела у многих проартикулят располагается непарная лопасть, нерасчлененный отдел, который у дикинсоний имеет субтреугольные очертания. Этот отдел выражен в разной степени среди таксонов Dipleurozoa и Cephalozoa; он интерпретируется здесь как элемент начального эволюционного развития цефализации Metazoa. Вполне вероятно, что у активно передвигавшихся животных на этом переднем конце концентрировались сенсорные клетки (механорецепторы, хеморецепторы, терморецепторы и другие), реагировавшие на различные формы раздражения. У маленьких и ювенильных проартикулят головной отдел имеет относительно большие размеры и может охватывать туловище не только спереди, но и с боков (Иванцов, 2008). Рост и переход к более взрослым формам у дикинсоний сопровождается скачкообразным увеличением числа изомеров, что может свидетельствовать о существовании в онтогенезе проартикулят особой личиночной стадии (Zakrevskaya, Ivantsov, 2017).
Сходство Cephalonega nom. nov. с проартикулятами и, прежде всего, с ювенильными экземплярами некоторых представителей класса Cephalozoa, определяется наличием головного отдела и характером расчленения тела, чередующимся расположением поперечных элементов туловищного отдела и мелкобугорчатой скульптурой наружной поверхности. Удивительной структурой, выявленной только у сюзьминской проартикуляты, являются две тонкие нити, отходящие от заднего конца расчлененной области (Ivantsov et al., 2019). Судя по тому, что они наблюдаются у экземпляров с хорошо видимыми бугорками, и их присутствие никак не сказывается на размерах и форме близлежащих изомеров, эти нити имеют эктодермальную природу.
Обращает на себя внимание массовое появление в захоронениях маленьких цефалонег примерно одного размера (рис. 1). Оно было обусловлено, скорее всего, биологическими, а не тафономическими причинами, и не гранулометрическим составом пород, поскольку во всех случаях организмы были захоронены на месте обитания без переноса и сортировки по размеру, а размер зерен песчаника, который теоретически мог бы повлиять на возможность различения отпечатков, во всех местонахождениях был разным. Явление может свидетельствовать о начале определенной фазы индивидуального развития животного, например, оседания плавающей стадии на дно.
Рис. 1.
Пропорции тела Cephalonega stepanovi (Fedonkin, 1976): а – для вида в целом, б – по отдельным захоронениям.
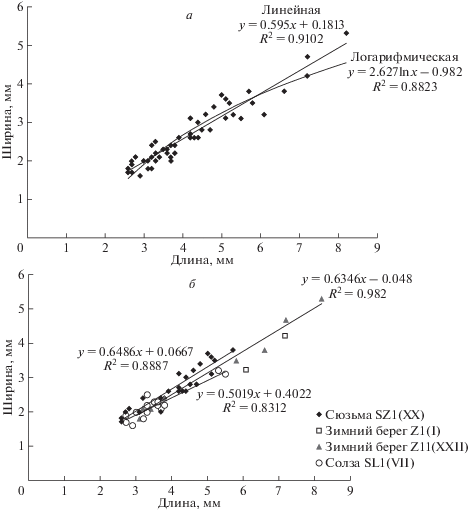
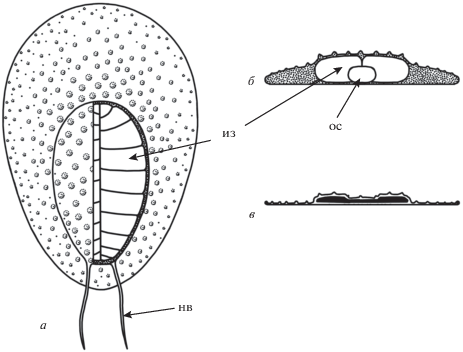
Рис. 2.
Схематическая реконструкция Cephalonega stepanovi (Fedonkin, 1976): а – вид сверху, эктодермальный покров на правой половине туловищного отдела удален, б – поперечное сечение, в – то же, для коллапсировавшего тела. Oбозначения: из – изомер, ос – осевая структура, нв – нитевидные выросты.
Ниже приводим уточненные морфологические характеристики типа Proarticulata и подчиненных таксонов высокого ранга. Авторы осознают трудности и условность надродовой систематики проартикулят, поскольку, несмотря на обильный ископаемый материал, очень многое остается неясным, особенно в части их анатомического строения, раннего онтогенеза, гистологии и физиологии.
Изученный материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН) (колл. №№ 3992, 3993, 4716, 4852, 4853). Полевые работы и подготовка каменного материала проведены при поддержке РФФИ, грант № 17-05-02212-a. Лабораторные и теоретические исследования поддержаны субсидией на выполнение фундаментальных исследований по теме госзадания ПИН РАН “Появление жизни, становление биосферы и развитие древних биот”.
Тип Proarticulata Fedonkin, 1985. Многоклеточные животные с низким широким телом, поделенным полностью или частично на два ряда однотипных элементов – изомеров, расположенных в чередующемся относительно продольной оси тела порядке. На переднем конце тела у многих представителей обособляется нерасчлененный головной отдел. У небольших форм он может окружать с боков расчлененный (туловищный) отдел.
Внутренние структуры представлены комплексами каналов. Один из них состоит из крупного осевого канала, одиночного или двойного, и серии более мелких боковых каналов, иногда дихотомически ветвящихся. Этот комплекс интерпретируется как пищеварительно-распределительная система. Другой комплекс состоит из множества тонких ветвящихся каналов, расположенных преимущественно в головном отделе. Он интерпретируется как секреторный орган, также, вероятно, связанный с пищеварением.
Судя по найденным вереницам следов в ассоциации с остатками тел следообразователей, по крайней мере, три вида проартикулят передвигались и питались верхним слоем микробных матов, разрушая его своей вентральной поверхностью. Пищеварение было либо наружное, либо происходило в особых карманах пищеварительно-распределительной системы, щелевидно открывавшихся на вентральную сторону. Протяженность следов свидетельствует о способности проартикулят относительно быстро перемещаться на значительные расстояния, что говорит о наличии у них развитой мускулатуры.
Рост проартикулят осуществлялся путем увеличения размеров тела и умножения числа изомеров. Прибавление новых изомеров происходило на заднем конце тела в течение всей жизни или только на начальных этапах онтогенеза. Для некоторых видов доказано существование личинки, характеризовавшейся относительно крупным головным и мало расчлененным туловищным отделами и, вероятно, имевшей иные, нежели у взрослого организма, экологические предпочтения.
Симметрия скользящего отражения, определяющая архитектонику ряда групп вендского бентоса, включая прикрепленные модулярные формы и подвижные билатеральные проартикуляты, интерпретируется как результат спирального роста тела (возможно, и спирального дробления яйца), что допускает предположение о филогенетической связи между этими группами (Fedonkin, 2017).
Класс Vendiamorpha Fedonkin, 1985. Проартикуляты преимущественно небольших размеров. Головной отдел отсутствует. Внешние концы изомеров обращены вбок и назад. Осевая структура состоит из осевого канала и боковых ответвлений, на один изомер приходится одно ответвление. К этому классу относятся роды Paravendia Ivantsov, 2004, Vendia Keller, 1969 и, возможно, Karakhtia Ivantsov, 2004.
Класс Dipleurozoa Harrington et Moore, 1955. Проартикуляты от небольших до очень крупных размеров. На ранней стадии развития организма его головной отдел имеет форму полукруга или широкого треугольника, а внешние концы изомеров обращены вбок и назад. На средних и поздних стадиях изомеры расположены субрадиально, а головной отдел по форме и размерам приближается к передним изомерам. Осевая структура состоит из одиночного или парного осевого канала и боковых ответвлений, число которых может быть существенно меньше числа изомеров. Класс включает в себя один род Dickinsonia Sprigg, 1947.
Класс Cephalozoa Ivantsov, 2004. Проартикуляты от небольших до крупных размеров. Головной отдел четко обособлен на всех стадиях онтогенеза, имеет форму от полукруглой до серповидной. Внешние концы изомеров обращены назад. Осевая структура состоит из осевого канала и боковых ответвлений, либо из одних ответвлений; на один изомер приходится одно ответвление. На спинной стороне ювенильных экземпляров иногда развита скульптура в виде мелких бугорков. Класс объединяет большинство родов проартикулят, в том числе: Andiva Fedonkin, 2002, Archaeaspinus Ivantsov, 2007, Cephalonega Fedonkin, nom. nov., Cyanorus Ivantsov, 2004, Ivovicia Ivantsov, 2007, Lossinia Ivantsov, 2007, Marywadea Glaessner, 1976, Ovatoscutum Glaessner et Wade, 1966, Podolimirus Fedonkin, 1983, Praecambridium Glaessner et Wade, 1966, Spriggina Glaessner, 1958, Tamga Ivantsov, 2007, Yorgia Ivantsov, 1999.
КЛАСС CEPHALOZOA
ОТРЯД И СЕМЕЙСТВО НЕ УСТАНОВЛЕНЫ
Род Cephalonega Fedonkin, nom. nov.
Onega (non Distant, 1908): Келлер, Федонкин, 1976, с. 42; Glaessner, 1979, c. 104; Федонкин, 1981, с. 20; Вендская система, 1985, с. 56; Федонкин, 1987, с. 104; Иванцов, 2007, с. 4, 5.
Название рода – от родового названия Onega и κεφαλή греч. – голова.
Типовой вид – Onega stepanovi Fedonkin, 1976 из позднего венда Архангельской обл.
Диагноз. Тело преимущественно с овоидальным контуром, с расширенным и слегка притупленным передним концом; подразделяется на два отдела: нерасчлененный головной и туловищный, поделенный на два ряда изомеров; при этом головной отдел окружает туловищный со всех сторон. Изомеры имеют вид резких овальных выступов, небольших, слегка удлиненных в поперечном к оси тела направлении, собранных в компактную группу. Спинная поверхность покрыта мелкими бугорками. От заднего края туловищного отдела отходят два тонких нитевидных выроста, направленные назад.
Сравнение. Род имеет сходство с проартикулятами мелких размеров и ювенильными формами крупных цефалозой полным охватом головным отделом области с изомерами (Tamga, Yorgia, Ovatoscutum) и бугорчатой скульптурой спинной поверхности (Yorgia, Archaeaspinus, Lossinia). Отличается овоидным контуром тела и маленькими, собранными в компактную группу изомерами и наличием нитевидных выростов.
Замечания. Формальный диагноз рода впервые был опубликован М. Глесснером (Glaessner, 1979). Положение переднего конца тела определено по аналогии с другими проартикулятами.
Состав. Типовой вид.
Cephalonega stepanovi (Fedonkin, 1976)
Табл. III , фиг. 5–10
Onega stepanovi: Келлер, Федонкин, 1976, с. 42, табл. III , фиг. 3, 6 ; Федонкин, 1981, с. 71, табл. XIII , фиг. 5, 6 ; Федонкин, 1985, с. 93, табл. XIX , фиг. 1, 2, 4, 8, 9 ; Иванцов, 2007, с. 4, табл. III , фиг. 7, 8 , рис. 3 , в.
Голотип – ПИН, № 3992/5049 (ранее ГИН, № 4464/57В), отпечаток; Архангельская обл., Летний берег Белого моря, прав. берег р. Сюзьма в 5.5 км выше устья, местонахождение Сюзьминское; верхний венд, усть-пинежская свита, сюзьминские слои, локальное скопление SZ1(XX) (Келлер, Федонкин, 1976, табл. III , фиг. 6 ; Федонкин, 1981, табл. XIII , фиг. 6 ; Вендская система, 1985, с. 93, табл. XIX , фиг. 8 ).
Описание. Контур тела у маленьких экземпляров эллиптический, у более крупных – овоидальный; максимальная ширина наблюдается вблизи переднего конца туловищного отдела. Длина переднего участка головного отдела (предшествующего туловищному) вдвое больше, чем заднего (находящегося позади от туловищного отдела), и чем его ширина с боков; и лишь немногим меньше длины туловищного отдела. Изомеры овальные, ориентированы субрадиально. Размер изомеров увеличивается от первой пары до четвертой, затем постепенно уменьшается. Видимое число изомеров – 8–10 пар.
Наблюдаются два варианта сохранности туловищного отдела. В первом варианте его поверхность ровная по высоте или слегка прогнутая в осевой части, изомеров не видно (табл. III , фиг. 10 ), либо их границы представлены тонкими бороздками; иногда заметна скульптура, состоящая из равномерно рассеянных бугорков (табл. III , фиг. 8, 9 ). Изомеры могут хорошо выделяться как на краях, так и на оси отдела, и демонстрировать характерное для проартикулят чередование (табл. III , фиг. 9 ). Во втором варианте на оси отдела располагается широкий и глубокий желобок, а изомеры имеют вид коротких, в той или иной степени деформированных валиков, упирающихся своими внутренними концами в осевую ложбину (табл. III , фиг. 6, 7 ). По всей видимости, эти два варианта сохранности отражают различные стадии разложения вещества туловищного отдела, сопровождавшегося коллапсом наименее плотных участков – внутренних частей изомеров и осевой структуры (рис. 2).
Головной отдел по внешнему краю окружен узкой каймой, наиболее рельефно выступающей на его переднем конце. Спинная поверхность покрыта мелкими коническими бугорками, размер которых немного увеличивается от внешнего края в сторону центра. От заднего края туловищного отдела отходят два тонких нитевидных выроста, направленные назад (табл. III , фиг. 6 ). Их длина составляет не менее половины длины туловищного отдела (Ivantsov et al., 2019, рис. 3 ).
Размеры (рис. 1). Минимальная измеренная длина составляет 2.5 мм, максимальная – 8.2 мм; имеется также один существенно больший по размерам, но плохо сохранившийся фрагментарный экземпляр (ПИН, № 4853/1120), предположительно относящийся к описываемому виду, неполная длина которого достигает 17 мм.
Распространение. Верхний венд, усть-пинежская свита, сюзьминские, зимнегорские, мезенская свита, ергинские слои; Юго-Восточное Беломорье, Архангельская обл.
Материал. 119 экз., в том числе: местонахождение Сюзьминское, локальное скопление SZ1(XX) – 80 экз. (ПИН, №№ 3992/401, 504, 505, 5003, 5005, 5006, 5008, 5016–5023, 5025–5046, 5049–5051, 5074, 5075, 5078, 5081–5089, 5102–5107, 5111–5131, 5133); местонахождение Карахтинское Южное, локальное скопление K2(VI) – 4 экз. (ПИН, №№ 4852/103, 111, 270, 276); местонахождение Солзинское, локальное скопление SL1(VII) “Основное” – 17 экз. (ПИН, №№ 4853/94, 158, 598, 600–603, 840–846, 1042, 1043, 1096), не проиндексированное скопление – 2 экз. (ПИН, №№ 4853/1120, 1121); местонахождение Зимнегорское, обнажение Еловое Южное – 1 экз. (ПИН, № 3993/6439), локальное скопление Z11(XXII) “Верхний желтый” – 10 экз. (ПИН, №№ 3993/5682, 5690, 6395-97, 6415, 6663–6666), локальное скопление Z1(I) “Кимберелловые линзы” – 5 экз. (ПИН, №№ 3993/5617, 5618, 5685, 5689, 6893).
Список литературы
Иванцов А.Ю. Мелкие вендские поперечно-рассеченные ископаемые // Палеонтол. журн. 2007. № 2. С. 3–10.
Иванцов А.Ю. Проартикуляты – вымерший в докембрии тип многоклеточных животных // Эволюционная морфология животных: к столетию со дня рождения акад. А.В. Иванова. Часть I / Ред. Мамкаев Ю.В. СПб.: Изд-во СПбГУ, 2008. С. 32–42 (Тр. СПб. об-ва естествоиспыт. Сер. 1. Т. 97).
Иванцов А.Ю. Следы питания проартикулят – вендских многоклеточных животных // Палеонтол. журн. 2011. № 3. С. 3–13.
Иванцов А.Ю., Малаховская Я.Е. Гигантские следы вендских животных. // Докл. Акад. наук. Сер. геол. 2002. Т. 385. № 3. С. 382–386.
Келлер Б.М., Меннер В.В., Степанов В.А., Чумаков Н.М. Новые находки Metazoa в вендомии русской платформы // Изв. АН СССР. Сер. геол. 1974. № 12. С. 130–134.
Келлер Б.М., Федонкин М.А. Новые находки окаменелостей в валдайской серии докембрия по р. Сюзьме // Изв. АН СССР. Сер. геол. 1976. № 3. С. 38–44.
Федонкин М.А. Беломорская биота венда (докембрийская бесскелетная фауна севера Русской платформы). М.: Наука, 1981. 99 с. (Тр. Геол. ин-та АН СССР. Т. 342).
Федонкин М.А. Систематическое описание вендских Metazoa // Вендская система. Историко-геологическое и палеонтологическое обоснование. Т. 1. Палеонтология. М.: Наука, 1985. С. 70–106.
Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции Metazoa. М.: Наука, 1987. 174 с. (Тр. ПИН АН СССР. Т. 226).
Bobrovskiy I., Krasnova A., Ivantsov A. et al. Simple sediment rheology explains the Ediacara biota preservation // Nature Ecol. & Evol. 2019. V. 3. № 4. P. 582–589.
Dunn F.S., Liu A.G., Donoghue P.C.J. Ediacaran developmental biology // Biol. Rev. 2018. V. 93. № 2. P. 914–932.
Dzik J., Ivantsov A.Yu. An asymmetric segmented organism from the Vendian of Russia and the status of the Dipleurozoa // Histor. Biol. 1999. V. 13. P. 255–268.
Dzik J., Martyshyn A. Taphonomy of the Ediacaran Podolimirus and associated dipleurozoans from the Vendian of Ukraine // Precambr. Res. 2015. V. 269. P. 139–146.
Evans S.D., Droser M.L., Gehling J.G. Highly regulated growth and development of the Ediacara macrofossil Dickinsonia costata // Plos One. 2017. V. 12. № 5. P. 1–15.
Fedonkin M.A. Andiva ivantsovi gen. et sp. nov. and related carapace-bearing Ediacaran fossils from the Vendian of the Winter Coast, White Sea, Russia // Ital. J. Zool. 2002. V. 69. P. 175–181.
Fedonkin M.A. Symmetry of gliding reflection in the Vendian (Ediacaran) invertebrates: a hint to the spiral mode of body growth, and probably, to the spiral cleavage of an egg in early metazoan evolution? // Abstr. IV Intern. Congr. Invertebr. Morphol. M.: Pero, 2017. P. 12.
Glaessner M.F. Precambrian // Treatise on Invertebrate Paleontology, Part A. Lawrence: Univ. Kansas Press, 1979. P. 79–117.
Gehling J.G., Droser M.L., Jensen S.R., Runnegar B.N. Ediacara organisms: relating form to function // Evolving form and function: fossils and development. New Haven: Yale Univ. Press, 2005. P. 43–66.
Glaessner M.F., Wade M. The Late Precambrian fossils from Ediacara, South Australia // Palaeontol. 1966. V. 9. P. 599–628.
Gold D.A., Runnegar B., Gehling J.G., Jacobs D.K. Ancestral state reconstruction of ontogeny supports a bilaterian affinity for Dickinsonia // Evol. Devel. 2015. V. 17. P. 315–324.
Hoekzema R.S., Brasier M.D., Dunn F.S., Liu A.G. Quantitative study of developmental biology confirms Dickinsonia as a metazoan // Proc. Roy. Soc. B. 2017. V. 284. P. 1–9.
Ivantsov A.Yu. Vendian animals in the phylum Proarticulata // Abstr. Intern. conf. IGSP Project 493 “The rise and fall of the Vendian biota”. Padova, 2004. P. 52.
Ivantsov A.Yu., Zakrevskaya M.A., Nagovitsyn A.L. Morphology of integuments of the Precambrian animals, Proarticulata // Invertebr. Zool. 2019. V. 16. № 1. P. 19–26.
Jenkins R.J.F. Functional and ecological aspects of Ediacaran assemblages // Origin and early evolution of the Metazoa / Eds. Lipps J., Signor P. N.Y.: Plenum Press, 1992. P. 131–176.
Runnegar B. Oxygen requirements, biology and phylogenetic significance of the late Precambrian worm Dickinsonia, and the evolution of the burrowing habit // Alcheringa. 1982. V. 6. P. 223–239.
Sperling E.A., Vinther J. A placozoan affinity for Dickinsonia and the evolution of late Proterozoic metazoan feeding modes // Evol. Devel. 2010. V. 12. P. 201–209.
Takiya D.M., Cavichioli R.R. A review of the Neotropical sharpshooter genus Onega Distant, 1908 (Hemiptera: Cicadellidae: Cicadellini) // Zootaxa. 2004. V. 718. № 1. P. 1–19.
Wade M. Dickinsonia: polychaete worms from the Late Precambrian Ediacara fauna, South Australia // Mem. Queensl. Museum. 1972. V. 16. № 2. P. 171–190.
Zakrevskaya M.A., Ivantsov A.Yu. Dickinsonia costata – the first evidence of neoteny in Ediacaran organisms // Invertebr. Zool. 2017. V. 14. № 1. P. 92–98.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал