

Палеонтологический журнал, 2019, № 5, стр. 33-38
Новые данные о биогеографии пермского семейства аммоноидей Perrinitidae
Т. Б. Леонова *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: tleon@paleo.ru
Поступила в редакцию 25.01.2019
После доработки 30.01.2019
Принята к публикации 12.02.2019
Аннотация
Представлены новые данные по биогеографическому распространению в раннепермских бассейнах аммоноидей семейства Perrinitidae Miller et Furnish, 1940 в связи с первой находкой представителя рода Properrinites Elias, 1938 на Урале. Обсуждаются возможные пути расселения этой, как ранее считалось, типично экваториальной группы аммоноидей. Предлагается схема, согласно которой перринитиды из области своего возникновения – экваториальных бассейнов Америки – проникли в центральные области океана Тетис, а затем оттуда расселились в западные и восточные части Тетической области и в Уральский палеобассейн.
ВВЕДЕНИЕ
Раннепермское семейство Perrinitidae Miller et Furnish, 1940 включает в себя восемь родов аммоноидей, отличающихся очень сложно рассеченной лопастной линией. Форма раковины у всех представителей пахиконовая, отличается от рода к роду небольшими вариациями в соотношениях высоты и ширины оборота и диаметра умбилика (Leonova, 2002).
Впервые перринитиды были найдены в пермских отложениях юго-западных штатов Америки, и номинативный род Perrinites был описан Э. Бёзе 100 лет назад (Böse, 1917). Примерно в это же время другой представитель этого семейства был обнаружен на о. Тимор. Он был определен и описан К. Ганиэлем (Haniel, 1915) как Cyclolobus subcumminsi. Несколькими годами позже Дж. Смит (Smith, 1927) описал с о. Тимор новый вид Perrinites brouweri и переописал вид Ганиэля как P. subcumminsi. Тем самым он де-факто установил присутствие перринитид в восточном полушарии. Тем не менее, еще довольно долгое время перринитиды считались семейством, характерным исключительно для западных штатов Америки и Мексики, и генозоны Properrinites и Perrinites широко использовались в биостратиграфии нижнепермских отложений этих областей (Böse, 1917; Plummer, Scott, 1937; Miller, Furnish, 1940 и др.).
О первых перринитидах с территории СССР стало известно по работам О.Г. Туманской (1937). Она описала несколько видов, найденных на Памире, и сравнила их с тиморскими, выделив для этих форм новый род Paraperrinites с типовым видом P. subcumminsi (Haniel). Более полные сведения о памирских и дарвазских перринитидах приведены в работах Туманской (1963) и Т.Б. Леоновой (1983; Леонова, Дмитриев, 1989; Левен и др., 1992).
В 80-х гг. прошлого века американский палеонтолог Д. Тэрэлсон при детальном изучении перринитид нашел четкие морфологические отличия между строением внутреннего отрезка лопастной линии в двух филогенетических ветвях, и на этом основании выделил два подсемейства: Perrinitinae и Paraperrinitinae (Tharalson, 1984). Дальнейший биогеографический анализ показал, что представители первого подсемейства были характерны только для западного полушария, а второго – для обоих на начальной стадии филогенеза, а на более поздних стадиях – для восточного полушария (Леонова, 1996). С этим выводом был не согласен китайский исследователь Цзоу Цурен (Zhou, Liengjarern, 2004), тогда как японский специалист М. Эхиро (Ehiro et al., 2005) в целом принял эту концепцию, но с некоторыми оговорками.
При этом все исследователи сходились во мнении, что аммоноидеи семейства Perrinitidae были обитателями экваториальных бассейнов, а в Бореальной области их не существовало (Tharalson, 1984; Zhou, Liengjarern, 2004; Ehiro et al., 2005; Leonova, 2011; Korn, Baets, 2015 и др.). В. Нассичук, описавший Properrinites с территории Юкона (Nassichuk, 1971), писал об этом местонахождении: “This is the most northerly occurrence of Properrinites and tends to support the hypothesis that lower Sakmarian strata in the northern Yukon were deposited in an equatorial setting” (Nassichuk, 1995, с. 232).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
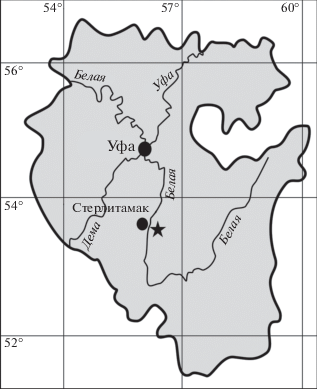
Во время полевых работ 2017 г. сотрудниками лаборатории моллюсков ПИН РАН А.В. Мазаевым и М.С. Бойко на раннепермском биогерме Шах-Тау в районе г. Стерлитамак в Башкортостане (рис. 1) был впервые найден представитель ассельско-сакмарского рода Properrinites Elias, 1938. Эта находка стала совершенно неожиданной, поскольку до настоящего времени бытовало общее убеждение, что представители семейства были провинциально ограниченными организмами, и их никогда ранее не находили на территории Урала и Предуралья.
Рис. 1.
Схема расположения местонахождения с Properrinites в республике Башкортостан, Урал. Звездочкой показано местоположение шихана Шах-Тау.
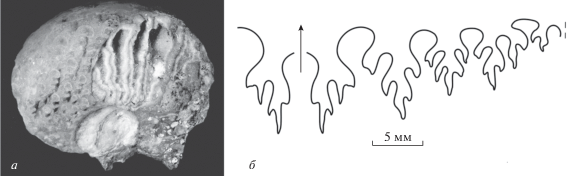
В карьере, разрабатываемом на месте шихана Шах-Тау, был обнаружен единственный экземпляр Properrinites, который представлен обломком довольно крупной раковины (диаметр около 60 мм) с хорошо сохранившейся наружной частью лопастной линии. Форма раковины пахиконовая, типичная для этого рода. Наиболее важным диагностическим признаком является сохранившаяся наружная часть лопастной линии. Она состоит из вентральной, трех боковых сложно-рассеченных лопастей и двураздельной умбиликальной лопасти (рис. 2). Степень рассечения лопастей в данной группе указывает на эволюционную ступень исследуемого таксона. Наиболее близкой формой, как по возрасту, так и по географическому положению, можно считать памирского Properrinites dmitrievi Ruzhencev, 1978 из ассельско-сакмарских отложений Памира.
Рис. 2.
Properrinites sp., экз. ПИН, № 5615/1: a – раковина, ×1; б – наружная часть лопастной линии; Башкортостан, шихан Шах-Тау; верхнеассельский подъярус (по: Leonova, Boiko, 2018).
Таксономический состав позднеассельского сообщества аммоноидей, обитавшего в районе распространения рифов, отличается от одновозрастных бассейновых сообществ на территории Уральского палеоокеана. Ранее здесь были найдены такие экстра-эндемики, как Shikhanites singularis Ruzhencev и Protopopanoceras sublahuseni Gerassimov, которые никогда не были встречены в других местонахождениях.
За все время изучения с 1930-х гг. в асселько-сакмарских известняках Стерлитамакских шиханов Шах-Тау и Тра-Тау были обнаружены следующие виды аммоноидей: Neopronorites rotundus (Maximova), Sakmarites asaphus (Ruzhencev), S. postcarbonarius tetragonus (Karpinsky), Artinskia subartiensis (Gerassimov), Medlicottia subdorbignyi Gerassimov, Shikhanites singularis Ruzhencev, Agathiceras uralicum (Karpinsky), Prothalassoceras biforme (Gerassimov), Somoholites shikhanensis Ruzhencev, Paragastrioceras sterlitamakense Gerassimov, Protopopanoceras sublahuseni Gerassimov (Герасимов, 1937; Руженцев, 1951; Королюк, 1985). Теперь этот список пополнился следующими формами: Properrinites sp., Neopronorites tenuis (Karpinsky) и множеством наутилоидей – около 20 видов (Барсков, Бойко, 2016; Leonova, Boiko, 2018).
Таким образом, уральский поднеассельский-раннесакмарский рифовый комплекс аммоноидей включает в себя 13 видов 11 родов. При сравнении его с сообществами аммоноидей, включающими в себя примитивных перринитид Properrinites и Subperrinites из других районов, выясняется следующее. Наиболее близким по родовому составу было раннесакмарское сообщество, остатки которого обнаружены на Юконе (Nassichuk, 1971). Здесь вместе с Subperrinites furnishi (Nassichuk) обнаружены виды родов Medlicottia, Prothalassoceras, Eoasianites, Uraloceras, Somoholites, Tabantalites, а также наутилоидов Liroceras и Titanoceras. Нужно отметить, что представители рода Tabantalites достаточно часто встречаются в ассельско-сакмарских отложениях Южного Урала (Руженцев, 1951). Основным аргументом в пользу раннесакмарского возраста местонахождения Нассичук (Nassichuk, 1971, 1995) считает присутствие примитивного Medlicottia, т. к. этот род раньше был известен, только начиная с позднесакмарского времени. Последняя находка примерно такого же примитивного Medlicottia на Урале позволяет предположить одновозрастность юконского и шахтаусского местонахождений.
Сообщество с Properrinites dmitrievi Ruzhencev из ташказыкской свиты Памира (позднеассельско-раннесакмарского возраста) включает в себя также виды родов Boesites, Metapronorites, Vanartinskia, Agathiceras, Prothalassoceras, Glaphyrites, Eoasianites, Svetlanoceras, Somoholites, Almites, Cardiella, Emilites, Tabantalites, Prostacheoceras, Martoceras (Руженцев, 1978). В этом случае родовой состав более богатый: кроме форм, известных из Башкирии, он содержит больше пролеканитид и видриоцератид, а также маратонитид.
На Тиморе Properrinites deroeveri Gerth найден вместе с видами родов Metapronorites, Akmilleria, Agathiceras, Somoholites, Juresanites, Metalegoceras, Kargalites, Propopanoceras (Haniel, 1915; Smith, 1927; Gerth, 1950). Особенностью этого сообщества является наличие металегоцератид, которые известны и в других районах, но в местонахождениях без перринитид.
В Техасе, Нью Мексико, Канзасе и Неваде примитивные перринитиды: Subperrinites bakeri (Plummer et Scott), S. denhami (Miller et Furnish), S. plummeri (Elias), S. moorae (Miller et Furnish) и Properrinites boesei (Plummer et Scott), P. nevadaensis Miller, Furnish et Clark встречены в отложениях формаций Bursum, Neal Ranch, Putnam, Wolfcamp, Alta, Lenox Hills, Council Grove, нижней части Hueco и нижней части Cibolo вместе с представителями родов Akmilleria, Aristoceras, “Stenolobulites”, Kargalites, Crimites, Propopanoceras (Böse, 1917; Plummer, Scott, 1937; Miller, Furnish, 1940; Miller et al., 1957 и др.).
В Южном Китае Properrinites dmitrievi Ruzhencev найден в отложениях формации Мапин (Maping) вместе с видами родов Boesites, Metapronorites, Agathiceras, Eoasianites, Subkargalites, Kargalites, Almites, Emilites (Zhou, 1987). В своей последней работе Цзоу (Zhou, 2017) переопределил Properrinites dmitrievi как новый вид P. gigantus Zhou, но это не меняет сути дела, особенно если учесть, что изученный материал очень плохой сохранности. В целом это сообщество очень близко памирскому из ташказыкской свиты даже на видовом уровне.
Во всех перечисленных областях в более молодых отложениях артинского–роудского возраста найдены более продвинутые роды перринитид: Metaperrinites и Perrinites (разрезы Техаса, Нью-Мексико, Канзаса, Оклахомы, а также Мексики, Гватемалы, Венесуэлы и Колумбии) и Metaperrinites, Perrimetanites и памирские роды-эндемики в восточном полушарии. В остальных регионах (Крым, Таиланд и Япония) перринитиды найдены только в артинском и кунгурском ярусах и представлены родами Metaperrinites, Perrimetanites и, возможно, Perrinites (Glenister et al., 1990; Леонова, 1996; Zhou, Liengjarern, 2004; Ehiro et al., 2005). В Таиланде (Ishibashi et al., 1996) указывали Properrinites bosei, но при повторном изучении этот образец был переопределен как Metaperrinites ishibashii (Zhou, Liengjarern, 2004)
Цзоу (Zhou, Liengjarern, 2004) пишет о возможности проникновения перринитид на территорию современного Таиланда через морские пути, проходящие вдоль геосинклиналей через Северо-Восточную Пацифику, Внутреннюю Монголию и Дальний Восток. Это может быть лишь отчасти справедливо. Нужно еще раз повторить, что на территории Японии не обнаружено примитивных перринитид в ассельских и сакмарских отложениях, т.е., первые перринитиды Тетической области появились не в северо-восточных и восточных частях Тетической области, а в ее центральных районах. Приводя в качестве аргумента отсутствие перринитид в ряде других бассейнов, в том числе и в Уральском, Цзоу утверждает, что эти аммоноидеи существовали лишь в зоне “активных поясов геосинклиналей” (in the active linear belts – geosynclines, the “open-sea”) (Zhou, Liengjarern, 2004, с. 322). Кроме этого, он считает, что находки перринитид в районах западной части Мидконтинента Америки связаны с посмертным переносом раковин, а не с обитанием этих организмов в платформенных бассейнах. С таким утверждением трудно согласиться. Во-первых, в этих районах (Сев. Техас, Канзас и Оклахома) известны находки представителей рода Shumardites, который считается предковым родом для семейства Perrinitidae (Руженцев, 1950), а также находки наиболее древнего из перринитид (Sub-per-rin-ites bakeri) (Tharalson, 1984). Во-вторых, перринитиды там обнаружены совместно с другими характерными членами аммоноидных сообществ, такими же, как и в геосинклинальных областях. В-третьих, последняя находка Properrinites в рифовых фациях Уральского бассейна также опровергает гипотезу Цзоу (Zhou, Liengjarern, 2004).
Относительно морского пути обмена фаунами между американской биогеографической областью и Тетическим океаном через северную часть Палеопацифики также имеются возражения. Перринитиды никогда не были найдены ни во Внутренней Монголии, ни в Приморье; наоборот, в этих областях в нижнепермских разрезах присутствуют типично арктические сообщества аммоноидей (Ehiro et al., 2005; Leonova, 2007).
В соответствии с концепцией М. Эхиро (Ehiro et al., 2005), обмен фаунами между экваториальными Американскими областями [или East Pacific Realm в соответствии со схемой районирования Д. Корна и К. де Байетса (Korn, Baets, 2015)] происходил через Пацифику, благодаря теплым течениям, которые доходили до восточных границ океана Тетис, где находились бассейны Таиланда и Японии. Затруднения в применении этой гипотезы связаны с тем, что в этих районах найдены лишь артинские–кунгурские перринитиды, а ранние, ассельско-сакмарские более примитивные формы указываются не из восточных районов, а из центральной части Тетис – о. Тимор, Южного Китая, Памира.
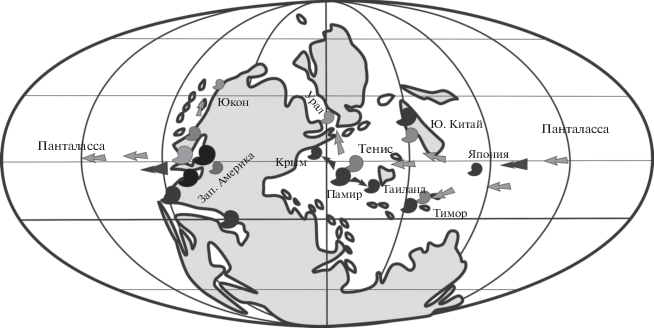
Учитывая все приведенные выше данные, мы предлагаем следующую схему. Из юго-западных районов Американского бассейна – центра возникновения перринитид, где обнаружены древнейшие формы [Subperrinites bakeri из формации Бурсум (Furnish, Glenister, 1971)], а также предположительно предковые формы перринитид, виды Properrinites через Пацифику проникли в центральную часть океана Тетис и обосновались в бассейнах Тимора, Южного Китая и Памира. Там они продолжали свое развитие и дали ряд более продвинутых форм (рис. 3). В Тиморском регионе Properrinites дал начало Paraperrinites, а в Южно-Китайском и Памирском регионах от Properrinites возникли Metaperrinites и Perrimetanites, а также специфические памирские формы. Perrimetanites, по-видимому, по сложности своего развития соответствовал уровню Perrinites и занял в тетических сообществах место последнего.
Рис. 3.
Схема расселения аммоноидей семейства Perrinitidae в раннепермскую эпоху (палеогеографическая реконструкция по: Ziegler et al., 1996). Серыми значками и стрелками показано распространение ассельско-сакмарских перринитид, черными значками и стрелками показано распространение артинско-кунгурских перринитид.
Присутствие представителей Perrinites в кунгурских отложениях Таиланда, как и Южного Китая, не является доказанным. Поскольку в работе Цзоу (Zhou, Liengjarern, 2004) отсутствует необходимая информация для определения рода, т.е. нет изображения внутренней части лопастной линии, и из текста следует, что она не была изучена, нельзя быть уверенным в правильности идентификации этой формы. Из приведенного рисунка наружной части лопастной линии (Zhou, Liengjarern, 2004, рис. 17, 3–4 ) можно заключить, что третья наружная боковая лопасть разделилась на две лопасти не полностью, что характерно для рода Perrimetanites. На большое сходство таиландского перринита с памирским Perrimetanites progressus Leonova указывали Б. Гленистер и У. Ферниш (Glenister et al., 1990). Вероятно, Perrinites действительно найден в Японии в Южном Китаками (Southern Kitakami Massif), что подтверждается исследованиями Эхиро (Ehiro et al., 2005). Независимо от наличия или отсутствия рода Perrinites в местонахождениях восточного полушария, можно рассматривать две возможности заселения этих бассейнов перринитидами во второй половине ранней перми.
С одной стороны, имеются неоспоримые доказательства присутствия позднеассельских–раннесакмарских Properrinites в местонахождениях Тимора, Памира, Южного Китая, а теперь еще и Урала, появившихся в самом начале раннепермской эпохи и развивавшихся в этих акваториях на протяжении нескольких геологических веков. Перринитиды достигли максимума разнообразия во второй половине ранней перми (в позднеартинское-кунгурское время), поэтому вполне логичным представляется вывод о том, что именно в это время они расселились в другие части океана Тетис: на запад (местонахождения Крыма) и на восток (местонахождения Японии и Таиланда). Вместе с тем, по нашему мнению, в конце ранней перми, благодаря теплым течениям, вполне могли происходить и параллельные миграции Perrinites из экваториальных вод Америки к восточным границам океана Тетис, как это предполагает Эхиро (Ehiro et al., 2005).
Присутствие Properrinites в верхнеассельских–раннесакмарских известняках рифа Шах-Тау подтверждает то, что в начале пермской эпохи морские связи между океаном Тетис и Уральским палеобассейном были вполне устойчивыми (Leonova, 2018). Строго говоря, Уральский бассейн не может считаться частью Бореальной биогеографической области в начале ранней перми, как полагали многие исследователи. Между Уралом и океаном Тетис происходили миграции не только в южном направлении, но и в сторону Уральской области. И только в середине артинского века морские связи с тетическими бассейнами были полностью прерваны, поскольку произошло полное закрытие южной части Уральского палеоокеана. Во второй половине ранней перми распространение перринитид происходило только в пределах экваториальных областей.
Список литературы
Барсков И.С., Бойко М.С. Сакмарские (ранняя пермь) наутилиды рифа Шах-Тау (Башкирия) // Сб. трудов всеросс. научн. конф. “Золотой век российской малакологии”. М. – Саратов, 2016. С. 207–211.
Герасимов Н.П. Уральский отдел пермской системы // Уч. зап. Казанск. гос. ун-та. 1937. Т. 97. Кн. 3–4. Геология. Вып. 8–9. С. 3–68.
Королюк И.К. Методы и результаты изучения пермского рифогенного массива Шахтау (Башкирское Приуралье). М.: Наука, 1985. С. 1–112.
Леонова Т.Б. Перринитиды Памира (Ammonoidea) // Палеонтол. журн. 1983. № 3. С. 39–53.
Леонова Т.Б. О систематике пермских аммоноидей семейства Perrinitidae // Палеонтол. журн. 1996. № 2. С. 25–33.
Леонова Т.Б., Дмитриев В.Ю. Раннепермские аммоноидеи Юго-Восточного Памира. М.: Наука, 1989. 198 с. (Тр. ПИН АН СССР. Т. 235).
Левен Э.Я., Леонова Т.Б., Дмитриев В.Ю. Пермь Дарваз-Заалайской зоны Памира: фузулиниды, аммоноидеи, стратиграфия. М.: Наука, 1992. 203 с. (Тр. ПИН РАН. Т. 253).
Руженцев В.Е. Верхнекаменноугольные аммониты Урала. М.: Изд-во АН СССР, 1950. 220 с. (Тр. ПИН АН СССР. Т. 29).
Руженцев В.Е. Нижнепермские аммониты Южного Урала. 1. Аммониты сакмарского яруса. М.: Изд-во АН СССР, 1951. 188 с. (Тр. ПИН РАН СССР. Т. 33).
Руженцев В.Е. Ассельские аммоноидеи на Памире // Палеонтол. журн. 1978. № 1. С. 36–52.
Туманская О.Г. О нахождении представителей рода Pеrrinites в СССР // Ежегодн. Всеросс. палеонтол. об-ва. 1937. Т. 11. С. 148.
Туманская О.Г. Пермские аммонеи Центрального Памира и их стратиграфическое значение. М.: Изд-во АН СССР, 1963. 119 с.
Böse E. The Permo-Carboniferous ammonoids of the Glass Mountains, west Texas, and their stratigraphical significance // Bull. Texas Univ. 1917. № 1762. P. 1–241.
Ehiro M., Hasegawa H., Misaki A. Permian ammonoids Prostacheoceras and Perrinites from the Southern Kitakami Massif, Northest Japan // J. Paleontol. 2005. V. 79. № 6. P. 1222–1228.
Furnish W.M., Glenister B.F. Permian Gonioloboceratidae (Ammonoidea) // Smithson. Contrib. Paleobiol. 1971. V. 3. P. 301–312.
Gerth H. Die Ammonoideen des Perms von Timor und ihre bedeutung fur die stratigraphische Gliederung der Permformation // N. Jb. Miner., Geol., Paläontol. Abh. Abt. B. 1950. Bd 91. H. 2. S. 233–320.
Glenister B.F., Furnish W.M., Zhou Z., Malai P. Ammonoid cephalopods from the Lower Permian of Thailand // J. Paleontol. 1990. V. 64. № 3. P. 479–480.
Haniel C.A. Die Cephalopoden der Dyas von Timor // Paläontol. von Timor. 1915. Lief. 3. P. 1–153.
Ishibashi T., Fujikawa M., Nakornsri N. Permian ammonoids from Loei area, northcentral Thailand // Mem. Fac. Sci. Kyushu Univ. Ser. Earth and Planetary Sci. 1996. V. 29. № 2. P. 175–196.
Korn D., De Baets K. Biogeography of Paleozoic ammonoids // Ammonoid Paleobiology: from Macroevolution to Paleobiogeography / Eds. Klug Ch. et al. Dordrecht: Springer, 2015. P. 145–161 (Topics in Geobiology 44).
Leonova T.B. Permian ammonoids: classification and phylogeny // Paleontol. J. 2002. V. 36. Suppl. 1. P. 1–114.
Leonova T.B. Correlation of the Kazanian of the Volga-Urals with the Roadian of the global Permian scale // Paleoworld. 2007. V. 16. P. 246–253.
Leonova T.B. Permian ammonoids: biostratigraphic, biogeographical and ecological analysis // Paleontol. J. 2011. V. 45. № 10. P. 1206–1312.
Leonova T. Biogeography of the Permian ammonoids of the Uralian paleoocean // Proc. of Kazan Golovkinsky Stratigraphic Meeting, 2017. Advances in Devonian, Carboniferous and Permian Research: Stratigraphy, Environments, Climate and Resources / Ed. Nurgaliev D. Bologna. Filodiritto Editore – Proceeding. 2018. P. 305–309.
Leonova T., Boiko M. A unique find of Perrinitids (Ammonoidea) in the Early Permian Shakh-Tau reef (Bashkortostan) // Proc. of Kazan Golovkinsky Stratigraphic Meeting, 2017. Advances in Devonian, Carboniferous and Permian Research: Stratigraphy, Environments, Climate and Resources / Ed. Nurgaliev D. Bologna. Filodiritto Editore – Proceeding. 2018. P. 163–166.
Miller A.K., Furnish W.M. Permian ammonoids of the Guadalupe Mountain region and adjacent areas // Spec. Pap. Geol. Soc. Amer. 1940. № 26. P. 1–242.
Miller A.K., Furnish W.M., Clark D.L. Permian ammonoids from western United States // J. Paleontol. 1957. V. 31. № 6. P. 1057–1068.
Nassichuk W.W. Permian ammonoids and nautiloids, South-eastern Eagle Plaine, Yukon Territory // J. Paleontol. 1971. V. 45. № 4. P. 1001–1021.
Nassichuk W.W. Permian ammonoids in the Arctic Regions of the world // The Permian of Northern Pangea / Eds. Scholle P.A., Peryt T.M., Ulmer-Scholle D.S. Berlin: Springer, 1995. P. 210–235.
Plummer F.B., Scott G. Upper Paleozoic ammonites in Texas // Bull. Univ. Texas. 1937. № 3701. P. 1–516.
Smith J.P. Permian ammonoids of Timor // Jaarb. Mijnwesen Ned. Indie. Verh. 1927. Bd 1. P. 1–58.
Tharalson D.B. Revision of the Early Permian ammonoid family Perrinitidae // J. Paleontol. 1984. V. 58. № 3. P. 804–833.
Zhou Z. First discovery of Asselian ammonoid fauna in China // Acta palaeontol. sin. 1987. V. 26. № 2. P. 131–148.
Zhou Z. Permian basinal ammonoid sequence in Nanpanjiang area of South China–possible overlap between basinal Guadalupian and platform-based Lopingian // J. Paleontol. 2017. V. 91. Mem. 74. P. 1–95.
Zhou Z., Liengjarern M. Lower Permian perrinitid ammonoids faunas from Thailand // J. Paleontol. 2004. V. 78. № 2. P. 317–339.
Ziegler A.M., Hulver M.L., Rowley D.B. Permian World Topography and Climate // Late glacial and Postglacial Environment Changes – Quaternery, Carboniferous-Permian and Proterozoic / Ed. Martini I.P. N.Y.: Oxford Univ. Press, 1996. (maps).
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал