

Палеонтологический журнал, 2019, № 5, стр. 63-68
Новые ручейники семейства Dysoneuridae (Insecta: Trichoptera) и личиночные домики (incertae familiaе) из нижнего мела Забайкалья
И. Д. Сукачева a, *, Д. В. Василенко a, b, *
a Палеонтологический институт им. А. А. Борисяка РАН
Москва, Роcсия
b Череповецкий государственный университет
Череповец, Россия
* E-mail: lab@palaeoentomolog.ru
* E-mail: lab@palaeoentomolog.ru
Поступила в редакцию 18.09.2018
После доработки 26.10.2018
Принята к публикации 01.04.2019
Аннотация
Описан новый род с тремя видами из семейства Dysoneuridae: Khasurtia kopylovi gen. et sp. nov., Kh. alexeii sp. nov. и Kh. lukashevichae sp. nov. из нижнего мела Забайкалья (Россия, местонахождение Хасуртый). Дается обновленная определительная таблица родов и видов семейства Dysoneuridae. Кроме имаго, описаны два вида личиночных домиков ручейников индузирода Terrindusia: Terrindusia (s. str.) khasurtica sp. nov. и T. (s. str.) buriatica sp. nov.
Семейство Dysoneuridae было описано из верхней юры (кимеридж) хребта Каратау (Казахстан). К настоящему времени его представители найдены в юрских и юрско-меловых отложениях Забайкалья, Якутии, на юге Сибири, в Монголии и Англии, а также в меловом бирманском янтаре (Сукачева, 1968, 1982, 1990; Novokshonov et al., 1995; Sukatsheva, Jarzembowski, 2001; Сукачева, Василенко, 2013; Wichard et al., 2018). Описание и последующее переописание семейства (Сукачева, Василенко, 2013) основаны исключительно на жилковании сохранившихся передних крыльев с характерным отсутствием апикальных развилков F1 и F4. Однако новые находки в бирманском янтаре дали возможность дополнить характеристику семейства многочисленными морфологическими признаками, хорошо различимыми на голове и теле насекомых (Wichard et al., 2018). Сохранились не только передние, но и задние крылья, а также типичные для этого семейства антенны, максиллярные и лабиальные пальпы, сложные глаза, глазки, гениталии и ноги с характерными шпорами.
Представители семейства Dysoneuridae, описываемые в данной работе, хорошо иллюстрируют эволюцию в направлении редукции жилок, подтверждая, что это не аберрация, а одно из естественных эволюционных направлений.
Новые представители семейства Dysoneuridae собраны в 2004, 2009 и 2014 гг. экспедиционными отрядами Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН из нижнемелового местонахождения Хасуртый (Хасурты). Местонахождение находится в Забайкальском крае (Бурятия, Закаменский р-н, окрестности хут. Хасуртый (25 км на восток от г. Закаменска, N 50°21′, E 103°37′).
Мезозойские отложения в этом районе относятся к нижней подсвите сангинской свиты гусиноозерской серии и датируются верхней юрой – нижним мелом. Всего в местонахождении Хасуртый собрано около 5000 экз. ископаемых насекомых: Ephemeroptera, Odonata, Plecoptera, Orthoptera, Grylloblattodea, ?Psocoptera, Thysanoptera, Homoptera, Heteroptera, Raphidioptera, Neuroptera, Coleoptera, Hymenoptera, Trichoptera, Lepidoptera, Mecoptera, Diptera. К настоящему времени оттуда описаны веснянки (Синиченкова, 2011), жуки (Legalov, 2010; Gratshev, Legalov, 2014), перепончатокрылые (Копылов, 2011, 2012, 2014; Копылов, Расницын, 2017), двукрылые (Blagoderov, Lukashevich, 2013) и мхи (Ignatov, Shcherbakov, 2011). Ручейников найдено около 200 экз. Помимо описываемых в настоящей работе Dysoneuridae, среди них, по предварительному определению, больше всего найдено представителей ископаемого семейства Vitimotauliidae, известного, главным образом, из раннего мела. Кроме того, В.Д. Иванов и С.А. Мельницкий (СПбГУ) определили в коллекции из Хасуртыя представителей некоторых современных семейств, а также древнего семейства Cladochoristidae. Такой состав фауны ручейников уникален для юрских и меловых отложений региона, и для своего объяснения требует более широкого анализа.
Материал, описанный в настоящей работе, хранится в коллекции лаборатории артропод ПИН РАН.
ОТРЯД TRICHOPTERA
ПОДОТРЯД INTEGRIPALPIA (=PHRYGANEINA)
СЕМЕЙСТВО DYSONEURIDAE SUKATSHEVA, 1968
Род Khasurtia Sukatsheva et Vasilenko, gen. nov.
Название рода от местонахождения Хасуртый.
Типовой род – Khasurtia kopylovi sp. nov.
Диагноз. Передние крылья средних размеров. Sc довольно короткая, параллельна R, впадает в передний край крыла на середине третьей четверти длины крыла. Субкостальное поле уже костального. R длинный, с ковшеобразным изгибом у вершины и с горбообразным искривлением перед ним. Начало развилка F2 расположено значительно проксимальнее начала F3. Развилки F1 и F4 отсутствуют. Ячеи DC и TC закрыты. Ячея MC открыта. Жилка cua2–cup короткая, прямая. Ячея TC длинная. Развилок F5 сидячий, длинный и широкий. CuP и A1 впадают в задний край крыла в одной точке в районе середины длины крыла. Жилки A2 и A3 короткие.
Состав. Три новых вида.
Сравнение. См. определительную таблицу.
Khasurtia kopylovi Sukatsheva et Vasilenko, sp. nov.
Табл. Х, фиг. 1 (см. вклейку)
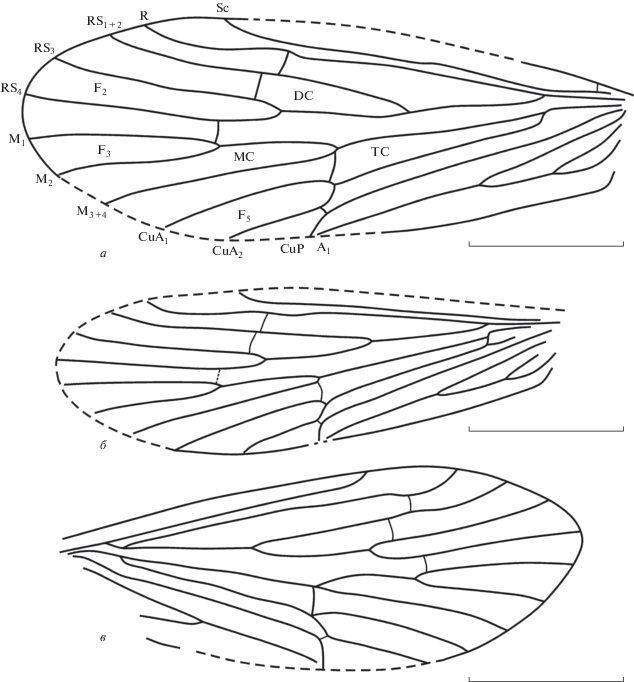
Рис. 1.
Представители семейства Dysoneuridae из местонахождения Хасуртый, нижний мел Бурятии: а – Khasurtia kopylovi gen. et sp. nov., голотип ПИН, № 5026/593; б – Kh. alexeii sp. nov., голотип ПИН, № 5026/578; в – Kh. lukashevichae sp. nov., голотип ПИН, № 5340/1469. Обозначения жилок на рисунке стандартные, кроме F2–F5, обозначающих апикальные развилки. Длина масштабной линейки 2 мм.
Название вида в честь палеоэнтомолога Д.С. Копылова.
Голотип – ПИН, № 5026/593, прямой и обратный отпечатки полного переднего крыла; Бурятия, местонахождение Хасуртый; нижний мел.
Описание (рис. 1, а). Крыло довольно узкое, длина больше ширины в 2.6 раза. Наибольшая ширина крыла на уровне впадения CuA1 в задний край крыла. Вершина крыла слабо вытянута, расположена между окончаниями RS и M1. Поперечная жилка r–rs1 + 2 слабо косая, расположена посередине явственного горбообразного изгиба на R. Поперечная rs1 + 2–rs3 прямая, расположена чуть дистальнее жилки r–rs1 + 2. Ячея DC средней длины и равна по длине ячее MC. RS разветвляется проксимальнее середины длины крыла. Развилок F2 длиннее F3 в 1.3 раза, основание его расположено намного проксимальнее основания развилка F3. Поперечная rs4–m1 прямая. Точка ветвления M расположена значительно дистальнее точки ветвления RS и чуть проксимальнее основания широкого и длинного развилка F5. Точки ветвления R–RS и M–CuA в основании крыла находятся на одном уровне. Слабо дугообразная жилка m3 + 4–cua1, закрывающая ячею TC, расположена чуть дистальнее вершины развилков M и CuA. CuP и A1 впадают в задний край крыла дистальнее середины длины крыла. Анальные жилки A2 и A3 довольно короткие. A2 короче A1 в 1.7 раза, A3 короче A1 в 1.6 раза.
Размеры в мм: длина крыла 8.3, ширина 3.2.
Сравнение. См. определительную таблицу.
Материал. Кроме голотипа, паратип ПИН, № 5340/218± из того же местонахождения.
Khasurtia alexeii Sukatsheva et Vasilenko, sp. nov.
Табл. Х, фиг. 2
Название вида – по имени палеоэнтомолога А.С. Башкуева.
Голотип – ПИН, № 5026/578, прямой и обратный отпечатки полного переднего крыла; Бурятия, местонахождение Хасуртый; нижний мел.
Описание (рис. 1, б). Крыло довольно узкое, длина больше ширины в 3 раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Вершина крыла закруглена и расположена между окончаниями RS4 и M1. Поперечная жилка r–rs1+2 прямая, расположена прямо посередине слабого горбообразного изгиба на R. Ковшеобразный изгиб у вершины R слабый. RS1+2 прямой. RS разветвляется немного проксимальнее середины длины крыла. Поперечные жилки r–rs1 + 2 и rs1 + 2–rs3 образуют одну слабо косую линию напротив середины горбообразного изгиба на R. Поперечная жилка rs4–m1 расположена чуть дистальнее начала развилка F3. Ячея MC открыта. Точка ветвления M значительно дистальнее точки ветвления RS и почти на одном уровне с основанием развилка F5. R–RS и M‒CuA разветвляются на одном уровне в основании крыла. Слабо дугообразная поперечная жилка m3+4–cua1, закрывающая ячею TC, расположена чуть дистальнее вершин развилков M и CuA. CuP и A1 впадают в задний край крыла на середине длины крыла. A2 и A3 довольно короткие. A2 короче A1 в 1.7 раза, A3 короче A1 в 3.0 раза.
Размеры в мм: длина крыла 7.5, ширина 2.5.
Сравнение. См. определительную таблицу.
Материал. Кроме голотипа, паратип ПИН, № 5340/221± из того же местонахождения.
Khasurtia lukashevichae Sukatsheva et Vasilenko, sp. nov.
Табл. Х, фиг. 3
Название вида в честь палеоэнтомолога Е.Д. Лукашевич.
Голотип – ПИН, № 5026/1469, прямой отпечаток полного переднего крыла; Бурятия, местонахождение Хасуртый; нижний мел.
Описание (рис. 1, в). Крыло довольно узкое, длина больше ширины в 2.8 раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Вершина крыла закруглена, находится между окончаниями RS4 и M1. Sc прямая, параллельна R. Поперечная слабонаклонная жилка r–rs1 + 2 расположена сразу за слабым горбообразным искривлением на R. Поперечная rs1 + 2–rs3, закрывающая ячею DC, прямая и расположена под небольшим углом к поперечной жилке r–rs1 + 2. Точка ветвления M расположена значительно дистальнее точки ветвления RS и на одном уровне с основанием довольно короткого и широкого развилка F5. Развилок F2 длиннее F3 в 1.3 раза. Основание его расположено проксимальнее основания F3. Ячея MC открыта. Точки ветвления R–RS и M–CuA в основании крыла расположены на одном уровне. Слабо дугообразная жилка m3+4–cua1 + 2 расположена в вершинах развилков M и CuA. CuP и A1 впадают в задний край крыла на середине длины крыла. A2 и A3 довольно короткие. A2 короче A1 в 1.8 раза. A3 сохранилась не полностью.
Размеры в мм: длина крыла 7.5, ширина 2.5.
Сравнение. См. определительную таблицу.
Материал. Кроме голотипа, паратип ПИН, № 5340/226 из того же местонахождения.
Определительная таблица родов и видов семейства Dysoneuridae по жилкованию передних крыльев и некоторым другим морфологическим деталям11 (Сукачева, Василенко, 2013, обновленная)
1 (10) Длина передних крыльев менее 4 мм.
2 (7) Скапус размером с голову...... Cretopsyche Wichard et al., 2018. 3 вида (в. мел, бирманский янтарь).
3 (4) RS и M разветвляются на одном уровне. Ячеи DC и MC открыты..............C. circula Wichard et al., 2018.
4 (3) RS и M разветвляются на разных уровнях.
5 (6) Ячеи DC и MC открыты..…........…......……………….. ......................................C. insueta Wichard et al., 2018.
6 (5) Ячея DC открыта, ячея MC закрыта....................... ……………………..............C. elegans Wichard et al., 2018.
7 (2) Скапус вдвое длиннее головы……….........…….. Burnapsyshe Wichard et al., 2018. 2 вида (в. мел, бирманский янтарь).
8 (9) Базальные членики жгутиков с дорсальными опушенными выростами.....…….......B. comosa Wichard et al., 2018.
9 (8) Базальные членики жгутиков простые. Максиллярные пальпы раздвоенные..................................... ………………............B. palpsfurcate Wichard et al., 2018.
10 (1) Длина передних крыльев не менее 4 мм.
11 (22) Длина передних крыльев 4–7 мм.
12 (13) M на большом протяжении слита с CuA. Ячея TC короткая.…..….....................Род Palaeoludus Sukatsheva et Jarzembowski, 2001. 1 вид P. popovi Sukatsheva et Jarzembowski, 2001 (н. мел Англии, Дорсет).
13 (12) M не слита с CuA. Ячея TC средней длины.
14 (19) R и RS1 слиты друг с другом; в предвершинной их части у самцов образуется бульбообразное утолщение…………………………...Род Liadotaulius Handlirsch, 1939 (=Oncovena Novokshonov et Sukatsheva, 1995) (syn. Ansorge, 2002). 3 вида.
15 (18) RS4 сливается с M1 на середине длины развилка F2.
16 (17) Поперечная жилка rs4–m1+2 длинная, сильно изогнутая; ячея MC закрытая.................................... ………..…............L. maior (Handlirsch, 1906) (н. юра Германии, Доббертин).
17 (16) Поперечная жилка rs4–m1+2 без изгиба; ячея MC открытая ……….….........L. borealis (Novokshonov et Sukatsheva, 1995) (ср. юра Красноярского края, Кубеково).
18 (15) RS4 не сливается с M1; ячея MC закрыта............. …………………..................L. sharategensis (Ivanov et Novokshonov, 1995) (в. юра Монголии, Шар-Тэг).
19 (14) R и RS1 не сливаются в одну жилку и не образуют у самцов бульбообразное утолщение................... …………………Род Dysoneura Sukatsheva, 1968. 2 вида.
20 (21) Поперечная r–rs1 расположена на уровне середины длины ячеи DC. Ячея TC открыта.................... .....……….....................D. trifurcata Sukatsheva, 1968 (Сукачева, 1968; в. юра Казахстана, Каратау).
21 (20) Поперечная жилка r–rs1 расположена значительно дистальнее середины длины ячеи DC. Ячея TC закрыта......………………….…………… D. zherikhini Sukatsheva et Vassilenko, 2013 (в. юра–н. мел Якутии, Кемпендяй).
22 (11) Длина передних крыльев более 7 мм.
23 (24) Субкостальное поле с поперечной жилкой sc–r в вершинной части крыла. R в последней трети без горбообразного изгиба. Жилка cua2 – cup длинная........ .......……........Prochita Sukatsheva et Vassilenko, 2013 (в. юра – н. мел Забайкалья, Черновские Копи; P. rasnitsyni Sukatsheva et Vassilenko, 2013).
24 (23) Субкостальное поле без поперечной жилки sc‒r в вершинной части крыла. R в последней трети с горбообразным изгибом. Поперечная жилка cua2–cup короткая.
25 (28) Субкостальное поле по ширине равно костальному ………………………….....Utiania Sukatsheva, 1982. 2 вида (в. юра–н. мел Забайкалья).
26 (27) Ячея MC открыта …U. defecta Sukatsheva, 1982 (в. юра–н. мел, Утан).
27 (26) Ячея MC закрыта..………......…….....U. remissa Sukatsheva, 1990 (в. юра–н. мел, Унда).
28 (25) Субкостальное поле уже костального в 2.5–3 раза. Поперечная жилка cua1–cup очень короткая...….……………….………..Khasurtia gen. nov. 3 вида (н. мел Бурятии, Хасуртый).
29 (32) Горбообразный изгиб на R слабый. Коленообразный изгиб на CuP явный.
30 (31) Поперечная жилка m3 + 4–cua1 расположена дистальнее вершин развилков M и CuA.................... …………………….…..........................Kh. alekseii sp. nov.
31 (30) Поперечная жилка m3 + 4–cua1 + 2 расположена в вершинах развилков M и CuA................................. …………………………..............Kh. lukashevichae sp. nov.
32 (29) Горбообразный изгиб на R очень заметный. Коленообразный изгиб на CuP слабый..................... ……………………………….................Kh. kopylovi sp. nov.
Из вышеприведенной определительной таблицы видно, что представитель семейства Dysoneuridae, а именно род Utania, был найден в верхней юре–нижнем мелу Забайкалья (местонахождения Утан и Унда). Этот род интересен сходством жилкования передних крыльев с родом Khasurtia, описываемым в данной работе. Только у двух упомянутых родов встречаются горбообразные изгибы в вершинной части R. Кроме того, они похожи в целом по общему плану жилкования. Такое сходство на родовом уровне может характеризовать как близкий геологический возраст местонахождений Утан, Унда и Хасуртый, так и указывать, что вероятнее, на сходные экологические условия. Необходимо отметить, что для местонахождения Унда принята верхнеюрская датировка, в то время как отложения Утана датируются нижним мелом. Интересно, что систематическая близость форм из Хасуртыя с формами из других местонахождений наблюдается, например, среди перепончатокрылых. В частности, найден вид ихневмонид, общий с нижнемеловым местонахождением Дабейгоу (Китай) – Amplicella shcherbakovi (Kopylov, Zhang, 2015). Вообще, все изученные к настоящему времени перепончатокрылые из Хасуртыя относятся, в основном, к типичным базально-меловым группам (Копылов, Расницын, 2017).
ЛИЧИНОЧНЫЕ ДОМИКИ РУЧЕЙНИКОВ
Индузирод Terrindusia Vialov, 1973
Индузиподрод Terrindusia s. str.
Terrindusia (s. str.) khasurtica Sukatsheva et Vasilenko, sp. nov.
Табл. Х, фиг. 4
Название вида от местонахождения Хасуртый.
Голотип – ПИН, № 5340/1453, прямой и обратный отпечатки личиночного домика; Бурятия, местонахождение Хасуртый; нижний мел.
Описание. Домики мелкие, прямые, узкие или слегка изогнутые, слабо сужающиеся к заднему концу. Построены из мелких (0.1–0.3 мм) песчинок, уложенных плотно и хаотично. Изредка среди них встречаются песчинки диаметром 0.5 мм и мелкие фрагменты растений (0.1–0.5 мм). Соединительного вещества среди строительных частиц мало или совсем нет (не заметно). На некоторых экземплярах к переднему концу домика песчинки увеличиваются до 0.5 мм.
Размеры в мм: длина домиков 7.0–8.0, ширина переднего конца 2.0–2.5, ширина заднего конца 1.0–1.5.
Сравнение. Описываемый вид близок к Terr. (s. str.) minuta Vialov et Sukatsheva, 1976 (н. мел Забайкалья и Монголии) по укладке и размерам строительного материала и размерам самих домиков. Отличается присутствием фрагментов растений.
Балл конструктивного совершенства (БКС) 30.
Материал. Кроме голотипа, из того же местонахождения имеется 13 паратипов ПИН, №№ 5340/149, 150, 151±, 152, 153, 155, 208, 209, 210±, 211±, 1050±, 1454±, 1455±.
Terrindusia (s. str.) buriatica Sukatsheva et Vasilenko, sp. nov.
Табл. Х, фиг. 5
Название вида от названия республики Бурятия.
Голотип – ПИН, № 5340/148, прямой и обратный отпечатки личиночного домика; Бурятия, местонахождение Хасуртый; нижний мел.
Описание. Домик средней величины, прямой, широкий, слабо сужающийся к заднему концу. Построен из мелких (0.1–0.2 мм) песчинок, уложенных плотно и хаотично, с добавлением довольно большого количества растительных фрагментов, особенно заметно увеличивающихся к переднему концу домика до 0.6 мм.
Размеры в мм: длина домика 13.0, ширина переднего конца 4.0, ширина заднего конца 2.5.
Сравнение. Описываемый вид близок к T. (s. str.) minuta и к T. (s. str.) khasurtica sp. nov. по укладке и составу строительного материала, отличаясь большим размером домика и большим количеством растительных частиц на переднем конце домика.
БКС 30.
Материал. Голотип.
Личиночные домики, описанные в этой работе, в соответствии с их конструктивными особенностями (БКС 30), относятся ко II ассоциации, ранее считавшейся характерной для неокома (Сукачева, 1982). К этой же группе относятся и домики ручейников, известные из местонахождения Утан, что не противоречит выводам, сделанным по имаго (см. выше). Домики ручейников, известные из местонахождения Унда (и других местонахождений ундино-даинского комплекса), представлены почти исключительно представителями индузирода Folindusia, поэтому их сравнение с описываемыми здесь Terrindusia затруднительно. Нет достаточных оснований также для отнесения описываемых здесь личиночных домиков к какому-либо семейству ручейников.
* * *
Авторы выражают глубокую благодарность А.П. Расницыну (ПИН РАН), А.Г. Пономаренко (ПИН РАН) и В.Д. Иванову (СПбГУ) за ценные советы и консультации, Д.Е. Щербакову (ПИН РАН) за предоставление актуальных данных о составе энтомофауны местонахождения Хасуртый и Н.С. Гороховой (ООО “Издательство АСТ”) за техническую помощь. Работа поддержана программой Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов” и грантом РФФИ № 18-04-00322.
Список литературы
Копылов Д.С. Ископаемые ихневмониды местонахождения Хасуртый в Забайкалье (Hymenoptera, Ichneumonidae) // Палеонтол. журн. 2011. № 4. С. 49–54.
Копылов Д.С. Новые виды преихневмонид (Hymenoptera, Ichneumonidae, Praeichneumonidae) из нижнего мела Забайкалья // Палеонтол. журн. 2012. № 1. С. 65‒70.
Копылов Д.С. Новые археоцинипиды (Hymenoptera, Cynipoidea, Archaeocynipidae) из нижнего мела Забайкалья // Палеонтол. журн. 2014. №1. С. 60–64.
Копылов Д.С., Расницын А.П. Новые сепульки (Hymenoptera: Sepulcidae) из нижнего мела Азии I. Parapamphilinae и Xyelulinae // Палеонтол. журн. 2017. № 1. С. 66–72.
Синиченкова Н.Д. Новые веснянки (Insecta: Perlida= Plecoptera) из позднемезозойского местонахождения Хасуртый, Забайкалье // Палеонтол. журн. 2011. № 5. С. 76–81.
Сукачева И.Д. Новые юрские ручейники из Каратау (Trichoptera) // Юрские насекомые Каратау. М.: Наука, 1968. С. 175–179.
Сукачева И.Д. Историческое развитие отряда ручейников. М.: Наука, 1982. 112 с. (Тр. ПИН АН СССР. Т. 197).
Сукачева И.Д. Ручейники. Phryganeida // Позднемезозойские насекомые Восточного Забайкалья. М.: Наука, 1990. С. 94–122 (Тр. ПИН АН СССР. Т. 239).
Сукачева И.Д., Василенко Д.В. Новые таксоны ручейников (Insecta, Trichoptera) с редуцированным жилкованием передних крыльев из мезозоя Азии // Палеонтол. журн. 2013. № 1. С. 57–63.
Blagoderov V.A., Lukashevich E.D. New Axymyiidae (Insecta: Diptera) from the Mesozoic of East Siberia // Polskie Pismo Entomol. 2013. V. 82. P. 257–271.
Ignatov M.S., Shcherbakov D.E. Lower Cretaceous mosses from Khasurty (Baikal Area of South Siberia) // Arctoa. 2011. V. 20. P. 19–42.
Gratshev V.G., Legalov A.A. The Mesozoic Stage of Evolution of the Family Nemonychidae (Coleoptera, Curculionoidea) // Paleontol. J. 2014. V. 48. № 8. P. 851–944.
Handlirsch A. Die fossilen Insekten und die Phylogenie der rezenten Formen. Leipzig: Withehn Engelmann Verl., 1906–1908. Bd 1–3. 1430 s.
Kopylov D.S., Zhang H. New ichneumonids (Insecta: Hymenoptera: Ichneumonidae) from the lower Cretaceous of north China // Cret. Res. 2015. V. 52. P. 591–604.
Legalov A.A. Checklist of Mesozoic Curculionoidea (Coleoptera) with description of new taxa // Baltic J. Coleopterol. 2010. V. 10. № 1. P. 71–101.
Novokshonov V., Ivanov V., Sukatsheva I. New Jurassic caddis flies (Insecta, Phryganeida = Trichoptera) from Siberia and Mongolia // Paleontol. J. 1995. V. 29. № 4. P. 157–163.
Sukacheva I., Jarzembovski E. Fossil caddisflies (Insecta: Trichoptera) from the Early Cretaceous of southern England II // Cret. Res. 2001. V. 22. P. 685–694.
Wichard W., Neumann C., Müller P., Wang B. Family Dysoneuridae (Insecta, Trichoptera) in Cretaceous Burmese amber // Cret. Res. 2018. V. 82. P. 138–146.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал