

Палеонтологический журнал, 2020, № 1, стр. 56-65
Новый род и вид скумбриевых рыб (Perciformes, Scombroidei, Scombridae) из нижнего олигоцена Кавказа
a Палеонтологический институт им. А.А. Борисяка РАН, Москва, Россия
* E-mail: aban@paleo.ru
Поступила в редакцию 29.03.2019
После доработки 02.04.2019
Принята к публикации 03.04.2019
Аннотация
Из нижнего олигоцена (нижний майкоп, пшехский горизонт) Северо-Западного Кавказа и Абхазии (реки Белая, Пшеха и Гумиста) описан новый род и вид скумбриевых рыб Caucombrus histiopterygius gen. et sp. nov. Новый таксон отличается от остальных известных Scombridae, как ископаемых, так и современных, высоким парусоподобным первым спинным плавником. Остеологические признаки Caucombrus gen. nov. позволяют сближать его с королевскими макрелями (подсемейство Scomberomorinae). Предполагается, что C. histiopterygius gen. et sp. nov. или близкий к нему вид Caucombrus gen. nov. населял в середине олигоцена и морской бассейн на территории Рейнского грабена (ФРГ).
Из нижнеэоценовых – среднемиоценовых отложений с территории бывшего СССР описано девять родов и 18 видов скумбриевых рыб (семейство Scombridae) и несколько неопределенных до вида форм (Банников, 1985, 2010; Monsch, Bannikov, 2012; Nazarkin, Bannikov, 2014); кроме того, пока остаются неописанными собранные относительно недавно материалы по скумбриевым из базального эоцена р. Хеу (Кабардино-Балкария) и разновозрастных отложений, обнажающихся по р. Пшехе (Апшеронский р-н Краснодарского края): среднеэоценовых (кумский горизонт), нижне-среднемиоценовых (тархан–чокрак) и верхнемиоценовых (средний сармат). Ниже приведено описание нового рода и вида Scombridae, Caucombrus histiopterygius gen. et sp. nov., из нижнего олигоцена (нижний майкоп, пшехский горизонт) Северо-Западного Кавказа и Абхазии (реки Белая, Пшеха и Гумиста).
Семейство Scombridae традиционно помещают в подотряд Scombroidei отряда окунеобразных (Perciformes) (Берг, 1940; Collette et al., 1984; Банников, 1985, 2010; Nelson, 2006 и др.). В последнее время, однако, на смену традиционной классификации организмов зачастую приходит формальная кладистическая систематика, построенная на компьютерной обработке молекулярных данных и часто противоречащая морфологическим свидетельствам. Результат кладистического анализа не всегда отражает реальные родственные связи (Вислобокова, 2019). В кладистической систематике скумбриевидным придается ранг отряда (Wiley, Johnson, 2010; Near et al., 2012, 2013; Betancur-R. et al., 2013, 2017; Nelson et al., 2016 и др.) в “отделе (division) Percomorphacea” (Wiley, Johnson, 2010), “кладе Pelagia” (Miya et al., 2013) или “серии Pelagiaria” (Betancur-R. et al., 2017). Высокие таксоны кладистической иерархии морфологически гетерогенны, что признают и сами кладисты (Miya et al., 2013); это побуждает нас использовать традиционную, естественную систематику. Трудно всерьез отнестись к объединению в одну кладу скумбриевидных и игловидных (Syngnathiformes), предлагаемому кладистами (Near et al., 2012; Betancur-R. et al., 2013, 2017): в морфологии рыб этих групп нет ничего общего. Следует отметить, что еще Э. и И. Ле Дануа (Le Danois, Le Danois, 1963) предприняли попытку выделения “отряда Scombres” на основании гипуростегии хвостового скелета. Однако этот признак связан с активным плаванием рыб, и в “отряде Scombres” оказались объединены многие не родственные семейства (Банников, 1985).
В семействе Scombridae выделяют от двух (Fraser-Brunner, 1950; Collette, Russo, 1979; Collette, Nauen, 1983; Collette et al., 1984) до пяти (Starks, 1910) подсемейств. Монотипический род Gasterochisma Richardson из Южного океана представляет собой сестринскую группу всех других современных скумбриевых (Santini et al., 2013) и обычно выделяется в подсемейство Gasterochismatinae (Fraser-Brunner, 1950; Collette, Russo, 1979; Collette, Nauen, 1983; Collette et al., 1984). Остальные скумбриевые либо подразделяются на подсемейства Scombrinae, Scomberomorinae и Thunninae (Банников, 1985, 2010), либо включаются в единое подсемейство Scombrinae, внутри которого выделяются современные трибы Scombrini, Scomberomorini, Sardini и Thunnini (Collette, Nauen, 1983; Collette et al., 1984) и ископаемая Eocoelopomini (Monsch, Bannikov, 2012). Подсемейство Scomberomorinae (или триба Scomberomorini), помимо нескольких ископаемых родов, основанных на скудном материале (Банников, 1985), включает в себя королевских макрелей (род Scomberomorus Lacepède) и ваху (род Acanthocybium Gill), характеризующихся сжатым с боков (а не веретеновидным) телом и увеличенным (до 64) числом позвонков (Collette, Nauen, 1983; Collette et al., 1984). Mолекулярные данные (Santini et al., 2013) противоречат монофилии Scomberomorini и Sardini, выявляя кладу Acanthocybium + + Gymnosarda.
Описанный ниже Caucombrus histiopterygius gen. et sp. nov. из нижнего олигоцена Кавказа обнаруживает значительное сходство с королевскими макрелями и ваху, но отличается озублением и высоким парусоподобным первым спинным плавником. Опубликованный недавно (Monsch, Micklich, 2018) каталог скумбриевидных рыб из олигоценового местонахождения Грубе Унтерфельд (Фрауэнвайлер; Баден-Вюртемберг, ФРГ), хранящихся в нескольких коллекциях в Германии (преимущественно в Гессенском земельном музее, Дармштадт), содержит сведения о нескольких экземплярах, сходных с новым таксоном. К. Монш и Н. Миклих отнесли этих рыб к Neocybium lingulatum (von Meyer, 1846) (Monsch, Micklich, 2018, c. 101–107, рис. 24, 25 ), отметив, что это единственный олигоценовый вид рода Neocybium Leriche, 1908. Однако Сybium lingulatum von Meyer описан по фрагментарному челюстному материалу и должен относиться к роду Scomberomorus (Банников, 1985, 2010). Что касается рода Neocybium, его типовой вид N. rostratum Leriche, 1908 из олигоцена Бельгии имеет существенно менее крупные зубы (Leriche, 1910), чем Caucombrus histiopterygius gen. et sp. nov. и германские экземпляры с высоким первым спинным плавником. Можно предположить, что среди материала из местонахождения Грубе Унтерфельд, определенного Моншем и Миклихом (Monsch, Micklich, 2018) как “Neocybium lingulatum”, имеются экземпляры, представляющие как род Neocybium, так и Caucombrus gen. nov. Местонахождение Грубе Унтерфельд, сформированное на Рейнском грабене, несколько моложе северокавказских местонахождений: оно датируется зоной NP23 (Maxwell et al., 2016), а последние – зоной NP20–21 (Попов и др., 2019).
Материал, описанный в настоящей статье, в основном собран автором в нескольких экспедициях и хранится в коллекции Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН).
Замечания М.В. Назаркина и Н.В. Зеленкова позволили улучшить статью. Фотографии выполнены С.В. Багировым (ПИН).
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ПОДОТРЯД SCOMBROIDEI
СЕМЕЙСТВО SCOMBRIDAE RAFINESQUE, 1815
Род Caucombrus Bannikov, gen. nov.
Название рода – от Кавказа и рода Scomber L., 1758; м. р.
Типовой вид – Caucombrus histiopterygius gen. et sp. nov.
Диагноз. Тело удлиненное, веретеновидное; его высота существенно уступает длине головы. Хвостовой стебель короткий и тонкий. Голова заостренная спереди, ее длина 2.5–3.9 раза укладывается в стандартной длине тела (SL). Ширина нейрокрания более чем вдвое уступает его длине. Гребень supraoccipitale относительно невысокий. Глаза относительно небольшие, диаметр орбиты не превышает 23% длины головы. Рот крупный, конечный, не выдвижной; верхняя челюсть выступает вперед. Восходящий отросток praemaxillare слит с сочленовным, протяженный в основании; его антеродорсальный край скошен. Сочленение нижней челюсти находится под задней четвертью орбиты. Зубы в челюстях относительно редко сидящие, довольно крупные, заостренные, однорядные, в основном сжатые с боков; несколько передних зубов изогнутые, конические. Praeoperculum слабо изогнуто, его нижняя ветвь короткая, а свободный край ровный (у мальков с шипиками). Позвонков 52–54, из них 24–26 абдоминальные и 28 каудальные; около половины задних туловищных позвонков несут парапофизы, сильно удлиняющиеся назад. Тела позвонков хвостового стебля постепенно укорачиваются к терминальному позвонку и не несут костного латерального киля. Остистые отростки позвонков очень тонкие, несколько передних невральных отростков слегка расширены медиально. Ребра относительно длинные, сильно наклонены назад. Спинные плавники соприкасаются в основании, первый из них протяженный и очень высокий спереди, парусовидный. В первом спинном плавнике около 22 очень тонких колючек; во втором спинном плавнике небольшая колючка и около 11 довольно коротких мягких лучей; дорсальных дополнительных плавничков около 11. В анальном плавнике две небольших колючки и около 10 мягких лучей, вентральных дополнительных плавничков около 11. Анальный плавник сдвинут назад относительно начала второго спинного. Грудные плавники умеренной длины. Брюшные плавники короткие, прикрепляются непосредственно перед грудными. Хвостовой плавник глубоко вырезанный, в нем 17 главных лучей, а также многочисленные верхние и нижние дополнительные (procurrent) лучи. Гипуростегия сильно развита. Чешуя очень мелкая, циклоидная.
Видовой состав. Типовой вид из нижнего олигоцена Кавказа и, возможно, неописанный олигоценовый вид из Германии.
Сравнение. Имея все диагностические признаки семейства Scombridae (см. Банников, 1985, 2010), Caucombrus gen. nov. отличается от остальных известных скумбриевых, как ископаемых, так и современных, высоким парусоподобным первым спинным плавником. Кроме того, новый род отличается от современного Gasterochisma (единственного представителя подсемейства Gasterochismatinaе) очень мелкой, а не крупной чешуей, большим числом позвонков (52–54 против 44 у Gasterochisma) и дополнительных плавничков (11 против 6–7). Автор считает обоснованным подразделение остальных Scombridae на подсемейства Scombrinae, Scomberomorinae и Thunninae (Банников, 1985, 2010). От первого из них Caucombrus gen. nov. резко отличается гораздо бóльшим числом позвонков (52–54 против 31 у Scombrinae), лучей в первом спинном плавнике (22 против не более 13) и дополнительных плавничков. Тунцовые (Thunninae), в отличие от Caucombrus gen. nov., имеют “корсет” из увеличенных чешуй в районе грудных плавников и обычно не более 46 позвонков [лишь современный вид Sarda sarda (Bloch) имеет 50–55 позвонков: Collette, Nauen, 1983]. Кроме того, тела позвонков хвостового стебля нового рода постепенно укорачиваются к терминальному позвонку и не несут костного латерального киля, тогда как у тунцовых второй и третий преуральные позвонки (pu) резко укорочены, а несколько предшествующих им позвонков несут костный латеральный киль.
Таким образом, особенности остеологии Caucombrus gen. nov. указывают на его принадлежность к подсемейству королевских макрелей Scomberomorinae (sensu Банников, 1985, 2010). Это подсемейство включает два современных рода – монотипный Acanthocybium и богатый видами Scomberomorus – и несколько ископаемых родов, известных преимущественно по фрагментарным материалам. По молекулярным данным (Santini et al., 2013), Acanthocybium образует сестринскую группу с родом тунцовых Gymnosarda Gill. Тело ваху (Acanthocybium) несколько более удлиненное, чем у нового таксона, а число позвонков существенно выше (62–64 против 52–54). В то же время, у ваху невысокий первый спинной плавник, меньше дополнительных плавничков (девять против 11) и другое озубление: челюстные зубы относительно мелкие и многочисленные (числом около 50: Devaraj, 1975), треугольные, сжатые с боков, тесно сидящие; длина основания восходящего отростка praemaxillare существенно превышает таковую Caucombrus gen. nov. Новый таксон обнаруживает существенное сходство с родом Scomberomorus в форме тела и меристических признаках; число его позвонков укладывается в пределы изменчивости Scomberomorus (41–56; Collette, Nauen, 1983; Collette, Russo, 1984), но туловищных позвонков больше (24–26, а у Scomberomorus – не более 22). Из 18 современных видов Scomberomorus большинство имеет менее 50 позвонков, но у S. guttatus (Bloch et Schneider) обычно имеется 50–51 позвонок, у S. munroi Collette et Russo – 51, у S. maculates (Mitchill) – 52, а у S. multiradiatus Munro – даже 55 позвонков (Collette, Nauen, 1983; Collette, Russo, 1984). У Scomberomorus меньше, чем у Caucombrus gen. nov., колючек в первом спинном плавнике, но больше лучей в спинном и анальном плавниках (Collette, Russo, 1984, табл. 9–11 ). Наконец, у Scomberomorus более широкий нейрокраний, никогда не бывает парусовидного спинного плавника, а все челюстные зубы сжаты с боков, треугольные, тогда как у нового рода несколько передних зубов изогнутые, конические.
Озубление Caucombrus gen. nov. (сочетание сжатых с боков зубов с коническими спереди) отличает его от ископаемых родов Scomberomorinae, описанных по фрагментарному челюстному и краниальному материалу, у которых неизвестно наличие или отсутствие парусовидного спинного плавника: у Aramichthys Signeux, 1959, Palaeocybium Monsch, 2005 и Neocybium Leriche, 1908 зубы сжаты с боков, треугольные, а у Landanichthys Dartevelle et Casier, 1949, Duplexdens Monsch, 2005, Wetherellus Casier, 1966, Scombramphodon Woodward, 1901 и Sphyraenodus Agassiz, 1844 зубы только конические (Банников, 1985; Monsch, 2005 и др.).
Для некоторых видов двух из трех эоценовых родов, включенных в incertae sedis трибу Eocoelopomini (Monsch, Bannikov, 2012), Eocoelopoma Woodward, 1901 и Palaeothunnus Bannikov, 1978, в дополнение к краниальному материалу, известен посткраний. Число позвонков у эоцелопомовых существенно меньше, чем у Caucombrus gen. nov. (до 41 против 52–54 у Caucombrus), а наличие парусовидного спинного плавника не отмечено. Все три рода Eocoelopomini (Eocoelopoma, Palaeothunnus и Micrornatus Monsch, 2005) имеют конические зубы в челюстях.
Caucombrus histiopterygius Bannikov, sp. nov.
Название вида от ιστίον греч. – парус и πτερόν греч. – перо, по парусоподобному первому спинному плавнику.
Голотип – ПИН, № 5419/14, отпечаток полного скелета с противоотпечатком задней половины тела; Краснодарский край, Апшеронский р-н, р. Пшеха напротив хут. Горный Луч; нижний олигоцен, пшехский горизонт, планорбелловые слои.
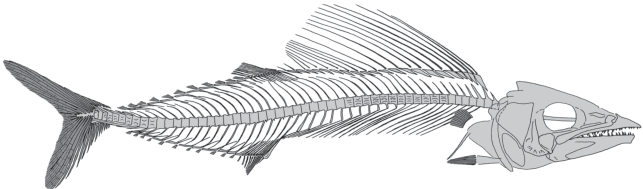
Описание (рис. 1–4). Описание вида в основном основано на двух относительно крупных экземплярах (рис. 1), остальной материал представляет собой молодь и мальков (см. рис. 3). Тело удлиненное, веретеновидное, с коротким и тонким хвостовым стеблем; его высота почти в 1.5 раза уступает длине головы. Голова коническая, заостренная спереди, ее длина до 3.9 раза укладывается в SL. Высота хвостового стебля в 5.5 раза меньше высоты тела. Глаза относительно небольшие, диаметр орбиты не превышает 23% длины головы. Рот крупный, конечный, не выдвижной; верхняя челюсть выступает вперед.
Рис. 1.
Caucombrus histiopterygius gen. et sp. nov.: а – голотип ПИН, № 5419/14, полный скелет; Краснодарский край, Апшеронский р-н, р. Пшеха напротив хут. Горный Луч; нижний олигоцен, пшехский горизонт, планорбелловые слои; б – паратип ПИН, № 1413/826, неполный, частично расчлененный скелет; Адыгея, р. Белая выше ст. Абадзехской; возраст тот же. Масштабная линейка 1 см.

Рис. 2.
Caucombrus histiopterygius gen. et sp. nov., паратип ПИН, № 1413/826, разрозненные нейрокраний (изнутри), нижняя челюсть (сбоку) и часть гиоидной дуги (сбоку); Адыгея, р. Белая выше ст. Абадзехской; нижний олигоцен, пшехский горизонт, планорбелловые слои. Обозначения: an – angulo-articulare, c – первые позвонки, ch – ceratohyale, d – dentale, eh – epihyale, nc – нейрокраний, ptp – posttemporale, uh – urohyale. Масштабная линейка 5 мм.

Рис. 3.
Caucombrus histiopterygius gen. et sp. nov., паратипы: а – ПИН, № 1413/827, молодой экз. без нейрокрания; б – ПИН, № 1413/828, малек, полный скелет; в – ПИН, № 1413/829, малек, скелет без хвостового плавника; Адыгея, р. Белая выше ст. Абадзехской; нижний олигоцен, пшехский горизонт, планорбелловые слои. Масштабная линейка 5 мм.

Нейрокраний невысокий, почти прямой парасфеноид проецируется примерно посередине орбиты. Ширина нейрокрания более чем вдвое уступает его длине (рис. 2). Гребень supraoccipitale невысокий. Этмоидный район черепа умеренно длинный. В орбите имеются окостенения склеротики. Сохранность не позволяет различить подглазничные кости. Нижнечелюстное сочленение расположено под задней четвертью орбиты. В верхней челюсти praemaxillare и maxillare тесно связаны друг с другом и образуют единый блок. Верхняя челюсть лишь слегка короче нижней. Рraemaxillare удлиненное, его восходящий отросток слит с сочленовным, протяженный в основании. Антеродорсальный край восходящего отростка скошен, его длина в 2.5 раза или более уступает длине альвеолярной ветви praemaxillare. Зубы в верхней челюсти однорядные, относительно редко сидящие, довольно крупные, заостренные, в основном сжатые с боков; несколько передних зубов изогнутые, конические. На противоотпечатке паратипа ПИН, № 1413/826 praemaxillare сохранилось наилучше; в нем насчитывается около 20 зубов, а кость покрыта продольной струйчатостью. Наиболее крупные зубы расположены в средней части кости; рострально (вероятно, и каудально) зубы постепенно укорачиваются. Maxillare удлиненное, очень слабоизогнутое, плотно примыкает к praemaxillare дорсо-латерально. Maxillare узкое спереди, слабо расширяется и уплощается дистально, оканчиваясь округленной пластинкой. Supramaxillare различимо плохо. Нижняя челюсть невысокая, ее высота почти в 4 раза уступает длине. Длина нижней челюсти составляет около 62% длины головы и соответствует длине по меньшей мере 10 последних туловищных позвонков. Симфиз невысокий, косой, со слабо намеченным подбородочным выступом. Неподалеку от симфиза на внутренней части dentale имеется отверстие сосуда. Dentale образует бóльшую часть нижней челюсти, с глубокой вырезкой на заднем крае; нижний отросток кости примерно одной длины с верхним. Вырезка на антеровентральном крае dentale (характерная для Scomberomorus: Collette, Russo, 1984, c. 579) едва намечена. Озубление нижней челюсти сходно с таковым верхней челюсти. Angulo-articulare по площади меньше, чем dentale, и плотно прилегает к нему сзади. Сочленовный отросток аngulo-articulare довольно короткий, но крепкий, крючковидный, покрыт продольной струйчатостью; retroarticulare не различимо. Hyomandibulare имеет форму перевернутой буквы L, с удлиненным передним мыщелком (для сочленения со sphenoticum); ось кости очень слабо наклонена вперед от вертикали. Метаптеригоид соединяет hyomandibulare с quadratum и ограничивает орбиту постеро-вентрально. Quadratum умеренно широкое, округло-треугольное, с утолщенным постеровентральным краем и небольшим сочленовным мыщелком. От ectopterygoideum различимы передний стержень и треугольный постеровентральный отросток, которым кость сочленяется с передним краем quadratum. Относительно крупное плоское entopterygoideum образует дно орбиты. От palatinum различима лишь задняя часть на паратипе № 1413/826, где кость выявляет короткую наружную ветвь для сочленения с ectopterygoideum и более длинную плоскую внутреннюю ветвь. Жаберная крышка умеренно широкая. Praeoperculum довольно крупное, плоское, слабо вогнуто вдоль утолщенного переднего края. Нижняя ветвь предкрышки несет следы ветвей предкрышечного канала сейсмосенсорной системы. Свободный край предкрышки шиповатый у мальков (см. рис. 3, б), ровный у более крупных экземпляров. Operculum крупное, плоское, широкое; его нижний край приострен. Передний выступ suboperculum слабый.
Кости гиоидного комплекса сохранились неполно. На паратипе № 1413/826 видны сочлененные между собой ceratohyale, epihyale и urohyale (рис. 2). Ceratohyale – плотная удлиненная кость, вогнутая как по верхнему, так и по нижнему краю, наиболее высокая в задней части. Ceratohyale несет антеровентральный выступ для сочленения с вентральным hypohyale. Если это не артефакт сохранности, ceratohyale имеет в передней части узкую щель параллельно верхнему краю кости (“beryciform foramen”: McAllister, 1968). Epihyale по форме приближается к равностороннему треугольнику, с ceratohyale сочленяется швом в средней части контакта (цератогиально-эпигиальный шов: McAllister, 1968). От пластинчатого удлиненного urohyale распознаются только невысокий передний конец и утолщенный нижний край. Точное число саблевидных, тонких radii branchiostegi неизвестно. Несколько разрозненных, в основном озубленных костей скелета жаберного аппарата сохранились на паратипе № 1413/826.
Позвонков 52–54, из них 24–26 туловищные и 28 каудальные [подсчитано по голотипу и паратипам ПИН, № 1413/826 (туловищные позвонки) и ПИН, № 1413/827 (хвостовые позвонки)]. Длина туловищной части позвоночника в 1.14 раза короче хвостовой его части. Линия позвоночника обычно слегка сигмоидно изогнута, приподнята спереди. Тела позвонков почти квадратные в латеральном аспекте, слегка пережатые посередине. Средние позвонки несут с каждой стороны по два продольных гребня, разделяющих три латеральные впадины (как у Neocybium и Acanthocybium) (то есть имеют “midlateral fossa”: Monsch, Micklich, 2018). Большинство остистых отростков очень тонкие и относительно длинные, прямые или, чаще, изогнутые. Несколько передних невральных отростков слегка расширены медиально. Передние гемальные отростки отходят от передней части позвонков. На передних хвостовых и задних туловищных позвонках сильно развиты гемальные презигапофизы. Около половины задних туловищных позвонков несут прямые узкие парапофизы, сильно удлиняющиеся каудально. Наклон парапофизов меняется с переднего на задний в каудальном направлении. Ребра относительно длинные, тонкие, сильно наклонены назад; передние из них прикрепляются к телам позвонков, а задние – к дистальной части парапофизов. Epineuralia различимы плохо.
Хвостовой скелет почти полностью скрыт гипуростегией (раздвоенными основаниями лучей хвостового плавника). Тела позвонков хвостового стебля постепенно укорачиваются к терминальному позвонку и не несут костного латерального киля. Второй преуральный позвонок заметно укорочен, его невральный гребень редуцирован, а гемальный отросток явно автогенный. Хвостовой плавник умеренно крупный, глубоко вырезанный, состоит из 17 главных лучей (I8-7I). Длина наибольших лучей равна 18–19% SL. Верхние и нижние дополнительные лучи относительно многочисленные, их не менее десяти.
Posttemporale – плоская эллиптическая кость с двумя сильными передними отростками, связывающими грудной пояс с нейрокранием. Cleithrum крупное, изогнутое, вогнутое спереди. Границы coracoideum, scapula и пекторальных radialia не различимы. Postcleithrum узкое, ребровидное. Грудной плавник среднего размера, его полная длина неясна. Точное число лучей в грудном плавнике неизвестно, но их не менее 15.
Тазовые кости узкие, клиновидные. Брюшные плавники короткие, прикрепляются непосредственно перед грудными, каждый состоит из слабой колючки и пяти мягких лучей. У мальков (см. рис. 3, б) брюшные плавники довольно длинные.
Спинные плавники соприкасаются в основании, первый из них протяженный и очень высокий спереди, парусовидный; в нем около 22 очень тонких колючек, постепенно укорачивающихся назад. Высота плавника короче длины его основания, но существенно превышает максимальную высоту тела. У голотипа дистальные концы колючек первого спинного плавника не сохранились, а у паратипа № 1413/826 высота плавника равна длине 17.5 задних туловищных позвонков. Самые длинные шипы первого спинного плавника – четвертый или пятый. Первая колючка сверхштатная, она примерно в 1.4 раза короче самого длинного шипа. Птеригиофоры первого спинного плавника узкие, клиновидные, с косо ориентированной проксимальной частью, едва доходящей до концов невральных отростков позвонков. Число птеригиофоров примерно соответствует числу невральных отростков позвонков, расположенных под первым спинным плавником.
Второй спинной плавник у голотипа начинается на вертикали третьего хвостового позвонка, в нем один жесткий и около 11 довольно коротких мягких лучей. Лучи расположены очень тесно, в задней части плавника несколько более разрежены. Передний мягкий луч самый длинный, каудально лучи укорачиваются. Высота второго спинного плавника превышает длину его основания. Дорсальных дополнительных плавничков у голотипа 11. Птеригиофоры второго спинного плавника очень узкие. В каудальном направлении птеригиофоры постепенно укорачиваются, и на них появляется все более удлиняющийся дистальный горизонтальный участок. Межневральные промежутки под вторым спинным плавником вмещают проксимальные концы двух птеригиофоров каждый. У птеригиофоров дополнительных плавничков дистальная часть сопоставима по длине с проксимальной, при этом кзади птеригиофоры распрямляются. Под задним концом птеригиофора последнего дополнительного плавничка расположен еще один узкий свободный птеригиофор.
Анальный плавник сходен со вторым спинным по форме и размерам, но сдвинут назад относительно начала второго спинного. В анальном плавнике две слабые колючки (передняя – сверхштатная, короче задней) и около 10–11 мягких лучей. Передний ветвящийся луч самый длинный, каудально лучи укорачиваются. Анальный плавник без резкой границы переходит в дополнительные плавнички, которых у голотипа насчитывается 11. Птеригиофоры анального плавника очень узкие, сходны по форме с противолежащими птеригиофорами второго спинного плавника и также укорачиваются спереди назад. Первый анальный птеригиофор заметно толще остальных. Проксимальные концы двух–трех птеригиофоров входят в межгемальные промежутки над анальным плавником. Птеригиофоры нижних дополнительных плавничков сходны с противолежащими птеригиофорами верхних дополнительных плавничков.
Чешуя очень мелкая, тонкая, циклоидная, образует сплошной покров на теле. Боковая линия не прослеживается.
Размеры в мм. SL голотипа – 171, длина головы – около 47. SL самых маленьких экземпляров: паратип ПИН, № 1413/828 – 20; экз. ПИН, № 3363/192 – 19.
В процентах к SL голотипа: длина головы – 26, высота тела – 17, первое предорсальное расстояние – 30, второе предорсальное расстояние – 65, преанальное расстояние – 67, вентроанальное расстояние – 37, длина наибольших лучей первого спинного плавника – >19, длина наибольших лучей хвостового плавника – 18.
Возрастные изменения. У мальков относительная длина головы существенно больше, чем у взрослых рыб (2.5 раза укладывается в SL). С возрастом уменьшается относительная длина брюшных плавников, а сочленение нижней челюсти перемещается назад под заднюю четверть орбиты; в онтогенезе утрачиваются шипы на свободном крае рraeoperculum.
Сравнение. Caucombrus gen. nov. известен по одному виду.
Замечания. Каталог скумбриевидных рыб из олигоценового местонахождения в Германии Грубе Унтерфельд (Фрауэнвайлер) (Monsch, Micklich, 2018) содержит сведения о нескольких экземплярах, сходных с новым таксоном. Монш и Миклих (Monsch, Micklich, 2018, c. 101–107, рис. 24, 25 ) отнесли этих рыб к Neocybium lingulatum (von Meyer). Однако вид lingulatum должен относиться к роду Scomberomorus (см. выше), а для рода Neocybium характерны значительно менее крупные зубы, чем описаны в Каталоге. Германский материал обычно в той или иной степени расчленен, что затрудняет его определение. Однако сохранность по меньшей мере трех изображенных экземпляров (Monsch, Micklich, 2018, рис. 24 с, d; 25c, d) позволяет различить высокий, парусообразный спинной плавник, подобно Caucombrus histiopterygius gen. et sp. nov. При описании этих экземпляров авторы указывают близкую к кавказскому таксону позвонковую формулу (Monsch, Micklich, 2018, с. 105, 106) (25–26 + 25–26, против 24–26 + 28 у C. histiopterygius) и наличие “midlateral fossa” на средних позвонках, как у C. histiopterygius. Судя по фотографиям, зубы у германских рыб имеют сходные с кавказским таксоном размеры; Монш и Миклих (Monsch, Micklich, 2018) описывают их как острые, несколько сжатые латерально и загнутые внутрь, то есть схожие с таковыми C. histiopterygius. У последнего несколько передних зубов изогнутые, конические, что не отмечено Моншем и Миклихом для германских рыб; впрочем, для распознавания этого признака необходима очень хорошая сохранность передней части челюстей, как у паратипа ПИН, №№ 1413/826.
Таким образом, можно предположить, что Caucombrus histiopterygius gen. et sp. nov. или близкий к нему вид рода Caucombrus gen. nov. населял в середине олигоцена (зона NP23: Maxwell et al., 2016) морской бассейн на территории Рейнского грабена.
Распространение. Северо-Западный Кавказ и Абхазия; нижний олигоцен, пшехский горизонт.
Материал. Помимо голотипа, четыре паратипа ПИН, №№ 1413/826–829 из планорбеллового слоя р. Белой (Адыгея) и два экземпляра мальков из эолискового слоя: экз. ПИН, № 1413/830 с р. Белой (Адыгея) и экз. ПИН, № 3363/192 с р. Гумиста (Абхазия).
Список литературы
Банников А.Ф. Ископаемые скумбриевые СССР // Тр. Палеонтол. ин-та АН СССР. 1985. Т. 210. 111 с.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. LXI + 244 с.
Берг Л.С. Система рыбообразных и рыб, ныне живущих и ископаемых // Ежегодн. Зоол. музея АН СССР. 1940. Т. 5. С. 87–517.
Вислобокова И.А. О новых подходах изучения эволюции макротаксонов и концепции экона // Палеонтол. журн. 2019. № 1. С. 3–12.
Попов С.В., Табачникова И.П., Банников А.Ф. и др. Лектостратотип майкопской серии по р. Белая выше г. Майкопа (Западное Предкавказье) в его олигоценовой части // Стратигр. Геол. корреляция. 2019. Т. 27. № 3. С. 70–92.
Betancur-R. R., Broughton R.E., Wiley E.O. et al. The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. 2013. P. 1–41.
Betancur-R. R., Wiley E.O., Arratia G. et al. Phylogenetic classification of bony fishes // BMC Evol. Biol. 2017. 17:162. P. 1–40. https://doi.org/10.1186/s12862-017-0958-3
Collette B.B., Nauen C.E. FAO species catalogue. Vol. 2. Scombrids of the World. An annotated and illustrated catalogue of tunas, mackerels, bonitos, and related species known to date. Rome: FAO, 1983. VII + 137 p. (FAO Fish. Synop. № 125. V. 2).
Collette B.B., Potthoff T., Richards W.J. et al. Scombroidei: development and relationships // Ontogeny and systematics of fishes/ Ed. Moser H.G., Richards W.J., Cohen D.M. et al. Lawrence: Allen Press, 1984. P. 591–620 (Amer. Soc. Ichthyol. Herpetol. Spec. Publ. № 1).
Collette B.B., Russo J.L. An introduction to the Spanish mackerels, genus Scomberomorus // Publ. Gulf States Mar. Fish. Comm. 1979. V. 4. P. 3–16.
Collette B.B., Russo J.L. Morphology, systematics, and biology of the Spanish mackerels (Scomberomorus, Scombridae) // Fish. Bull. 1984. V. 82. № 4. P. 545–692.
Devaraj M. Osteology and relationships of the Spanish mackerels and seerfishes of the tribe Scomberomorini // Ind. J. Fish. 1975. V. 22. № 1/2. P. 1–67.
Fraser-Brunner A. The fishes of the family Scombridae // Ann. Mag. Natur. Hist. Ser. 12. 1950. V. 3. № 26. P. 131–163.
Le Danois E., Le Danois Y. L’ordre des Scombres // Mém. Inst. Franç. Afr. Noire. 1963. № 68. P. 153–192.
Leriche M. Les poissons oligocènes de la Belgique // Mém. Mus. Roy. Hist. Natur. Belg. 1910. T. 5. P. 229–363.
Maxwell E.E., Alexander S., Bechly G. et al. The Rauenberg fossil Lagerstätte (Baden-Württemberg, Germany): A window into early Oligocene marine and coastal ecosystems of Central Europe // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2016. V. 463. P. 238–260.
McAllister D.E. The evolution of branchiostegals and associated opercular, gular, and hyoid bones, and the classification of teleostome fishes, living and fossil // Bull. Nat. Mus. Can. 1968. V. 221. 239 p.
Miya M., Friedman M., Satoh T.P. et al. Evolutionary origin of the Scombridae (tunas and mackerels): Members of a Paleogene adaptive radiation with 14 other pelagic fish families // PLOS One. 2013. V. 8. № 9. P. e73535:1–19.
Monsch K.A. Revision of the scombroid fishes from the Cenozoic of England // Trans. Roy. Soc. Edinburgh: Earth Sci. 2005 (2004). V. 95. P. 445–489.
Monsch K.A., Bannikov A.F. New taxonomic synopses and revision of the scombroid fishes (Scombroidei, Perciformes), including billfishes, from the Cenozoic of territories of the former USSR // Earth and Environ. Sci. Trans. Roy. Soc. Edinburgh. 2012 (2011). V. 102. P. 253–300.
Monsch K.A., Micklich N. Catalogue of the scombroid fishes from the Grube Unterfeld (“Frauenweiler”) fossil site at Rauenberg (Baden-Württemberg, S. Germany) // Current Research in Vertebrate Paleontology 4 / Eds Gruber G., Micklich N. Darmstadt, 2018. P. 7–154 (Kaupia. Darmst. Beitr. Naturges. H. 22).
Nazarkin M.V., Bannikov A.F. Fossil mackerel (Actinopterygii: Scombridae: Scomber) from the Neogene of south-western Sakhalin, Russia // Zoosyst. Ross. 2014. V. 23. № 1. P. 158–163.
Near T.J., Dornburg A., Eytan R.I. et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes // Proc. Nat. Acad. Sci. USA. 2013. V. 110. P. 12738–12743.
Near T.J., Eytan R.I., Dornburg A. et al. Resolution of ray-finned fish phylogeny and timing of diversification // Proc. Nat. Acad. Sci. USA. 2012. V. 109. P. 13698–13703.
Nelson J.S. Fishes of the World. 4th ed. Hoboken, NJ: Wiley, 2006. 601 p.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken, NJ: Wiley, 2016. 707 p.
Santini F., Carnevale G., Sorenson L. First molecular scombrid timetree (Percomorpha: Scombridae) shows recent radiation of tunas following invasion of pelagic habitat // Ital. J. Zool. 2013. V. 80. P. 210–221.
Starks E.C. The osteology and mutual relationships of the fishes belonging to the family Scombridae // J. Morphol. 1910. V. 21. P. 77–100.
Wiley E.O., Johnson G.D. A teleost classification based on monophyletic groups // Origin and Phylogenetic Interrelationships of Teleosts / Eds Nelson J.S., Schultze H.-P., Wilson M.V.H. Munich: Verl. Dr. F. Pfeil, 2010. P. 123–182.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал