

Палеонтологический журнал, 2020, № 1, стр. 6-13
Копролит с конхостраками из терминальной перми Центральной России – палеобиологическое и стратиграфическое значение
А. Г. Сенников a, b, *, В. В. Буланов a, b, **, Ф. Шольце b, c, ***
a Палеонтологический институт им. А.А. Борисяка РАН, Москва, Россия
b Казанский федеральный университет, Казань, Россия
c Геологический институт, Технический университет Фрайбергская горная академия, Фрайберг, Германия
* E-mail: sennikov@paleo.ru
** E-mail: bulanov@paleo.ru
*** E-mail: Frank.Scholze@geo.tu-freiberg.de
Поступила в редакцию 24.12.2018
После доработки 05.02.2019
Принята к публикации 05.02.2019
Аннотация
Впервые описан копролит с конхостраками из местонахождения вязниковского фаунистического комплекса Быковка (терминальная пермь Владимирской обл., Россия). Наиболее вероятными продуцентами подобных копролитов могли быть сеймуриаморфы рода Microphon (Kotlassiidae), недавно обнаруженные в этом местонахождении. Состав комплекса конхострак дает дополнительное независимое биостратиграфическое подтверждение наиболее высокого положения местонахождения Быковка в пермской части разреза Вязников.
ВВЕДЕНИЕ
В Центральном регионе Европейской России, на востоке Владимирской области в районе гг. Гороховец и Вязники описаны наиболее полные, непрерывные разрезы пограничных континентальных отложений перми и триаса, охарактеризованные органическими остатками как ниже, так и выше границы. В соответствии с региональной стратиграфической шкалой Восточной Европы положение границы определяется по смене комплексов тетрапод (вязниковского на спасский), различных групп рыб, беспозвоночных и растений. В настоящее время имеющиеся данные позволяют детально проследить историю развития отдельных групп организмов и всей биоты на рубеже палеозоя и мезозоя в данном регионе (Сенников, 1995, 2017; Sennikov, 1996; Ивахненко и др., 1997; Sennikov, Golubev, 2005, 2006; Сенников, Голубев, 2006, 2012, 2017; Голубев и др., 2012; Lebedev et al., 2015; Шишкин и др., 2018).
ПОЗДНЕПЕРМСКОЕ МЕСТОНАХОЖДЕНИЕ БЫКОВКА В КОМПЛЕКСЕ МЕСТОНАХОЖДЕНИЙ ВЯЗНИКИ
В г. Вязники и его окрестностях было открыто несколько позднепермских и раннетриасовых местонахождений фауны и флоры. Местонахождение Быковка расположено на северо-западной окраине г. Вязники в песчаном карьере, в его юго-восточной части, где представлена самая верхняя часть разреза верхнепермских отложений в районе Вязников (Sennikov, Golubev, 2006; Lebedev et al., 2015; Сенников, 2017).
Разрез местонахождения Быковка приводится ниже (описание слоев снизу вверх):
P3v Слой 1. Пески красновато-желтые полимиктовые разнозернистые с косослоистыми сериями до 1 м мощностью с резким наклоном слойков; косая слоистость сильно срезанная, разнонаправленная. Местами в тонких (1–3 см) алевритистых прослоях встречаются редкие фрагменты костей позвоночных и копролиты. Видимая мощность – 5–6 м.
Слой 2. Линзовидное переслаивание красновато-желтых горизонтальнослоистых, в отдельных линзовидных прослоях полого-косослоистых, полимиктовых мелкозернистых песков, пестроцветных алевритов и алевритистых глин с редкой галькой пестрых глин, многочисленными копролитами, чешуей рыб, реже – костями тетрапод и рыб. Линзовидные прослои песка срезают нижележащие напластования внутри слоя 2 и залегают на них с размывом. В основании таких песчаных прослоев местами наблюдаются пятна более темноокрашенных песков, обогащенных органическими остатками. По простиранию к северо-западу песчаные прослои замещаются переслаиванием пестроцветных алевритистых глин и алевритов. Нижняя граница слоя четкая, неровная, с размывом. Мощность – 0.5–2 м.
Q Слой 3. Суглинки глинистые буровато-красные с валунами. Слой наблюдается по северо-восточному борту карьера. Видимая мощность – 2–3 м.
Местонахождение Быковка имеет аллювиальный генезис. Слой 1 представляет собой русловые отложения, а слой 2 – прибрежно-отмельные (пески и алевриты) и пойменные (глины и алевриты). Разрез местонахождения демонстрирует верхнюю, завершающую часть аллювиального седиментационного цикла, и сформирован при заполнении русла в фазе затухания силы потока. Захоронение костей и копролитов в алевролитовых прослоях слоя 1, очевидно, совпадало с кратковременными эпизодами ослабления течения в основном русле и заиления дна. Слой 2 образован на участке мелководья в остаточной протоке с непостоянной интенсивностью течения, в депрессиях дна которых после непродолжительного переноса и сортировки накапливались и захоранивались остатки позвоночных и их копролиты. Не исключено, что данный участок прибрежной, краевой части русла временами подвергался частичному осушению, так как кости тетрапод и копролиты иногда демонстрируют признаки изменений при нахождении в субаэральных условиях: для первых – это трещины усыхания, а для вторых – рыхлая консистенция, приобретенная в результате мацерации.
Местонахождение Быковка занимает наиболее высокое положение в пермской части разреза Вязников, непосредственно ниже границы перми и триаса, что подтверждают и данные по позвоночным (Sennikov, Golubev, 2006; Миних и др., 2014; Lebedev et al., 2015; Сенников, 2017). Однако на северо-западной окраине Вязников, где расположено местонахождение, нижнетриасовых отложений не обнаружено. К востоку от Вязников начинается участок распространения нижнетриасовых (вохминских) отложений, охарактеризованных в местонахождении Федурники органическими остатками (остракоды, конхостраки, рыбы, тетраподы). Для определения положения границы перми и триаса в данном регионе детальная фаунистическая характеристика местонахождения Быковка имеет особо важное значение.
КОПРОЛИТЫ КАК ИСТОЧНИК ИНФОРМАЦИИ О БИОТИЧЕСКИХ КОМПЛЕКСАХ НА РУБЕЖЕ ПЕРМИ И ТРИАСА
Копролиты позвоночных в большом количестве были обнаружены в терминальнопермских местонахождениях органических остатков в районе гг. Вязники и Гороховец во время систематических полевых работ Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН), проводимых с 1999 г. по настоящее время, а также в результате совместных Польско-Российских палеонтологических экспедиций 2008, 2010 и 2013 гг. Копролиты были подробно описаны, выделено девять их морфотипов (“A”–“I”), представленных наиболее полно в местонахождении Быковка. Каждый из морфотипов предположительно соотнесен с определенной группой рыб или тетрапод (Owocki et al., 2012; Niedźwiedzki et al., 2016; Bajdek et al., 2016, 2017; Сенников, 2017). Копролиты морфотипов “A”–“H” содержат чешую рыб и обломки костей. Копролиты морфотипов “C” и “D” со следами спирального клапана принадлежат, вероятно, хищным рыбам, скорее всего, акулам или, может быть, двоякодышащим и лучеперым (Niedźwiedzki et al., 2016). Продуцентами копролитов морфотипа “A” были, наиболее вероятно, хищные зверообразные рептилии – тероцефалы, а морфотипа “B” – базальные архозавры протерозухиды (Owocki et al., 2012; Niedźwiedzki et al., 2016). Копролиты морфотипов “E”–“H” принадлежат, вероятно, хищным амфибиям или рептилиям, может быть, темноспондильным лабиринтодонтам, микрозаврам, сеймуриаморфам или хрониозухиям (Niedźwiedzki et al., 2016). Только копролиты одного особого морфотипа “I” без костных остатков, но с редкими остатками растений, принадлежат растительноядным тетраподам, скорее всего, дицинодонтам (Niedźwiedzki et al., 2016; Сенников, 2017).
КОПРОЛИТ С КОНХОСТРАКАМИ ИЗ МЕСТОНАХОЖДЕНИЯ БЫКОВКА
Конхостраки имеют большое значение для стратиграфии позднего палеозоя и мезозоя, для корреляции континентальных отложений этого возраста и для более точного обоснования границы перми и триаса в частности. Богатая и разнообразная фауна конхострак обнаружена в ряде позднепермских и триасовых местонахождений в районе Вязников и Гороховца (Scholze et al., 2015, 2019). Однако наиболее высокая, сложенная преимущественно песками часть разреза позднепермских отложений не была охарактеризована конхостраками. В частности, в местонахождении Быковка найти определимых конхострак до сих пор не удавалось. Только в копролите из этого местонахождения (слой 2) была обнаружена единственная раковина конхостраки плохой сохранности, определения для которой приведено не было (Owocki et al., 2012).
Во время полевых работ ПИН РАН в 2014 г. на местонахождении Быковка были собраны многочисленные копролиты. Один из копролитов морфотипа “F” из слоя 2 содержит раковины конхострак (экз. ПИН, № 5078/10; рис. 1, 2). Он имеет несколько неправильную коротко-овальную форму, со слегка суженным одним концом и немного расширенным противоположным; длина копролита – около 55 мм, максимальная толщина – 20 мм. Весь копролит заполнен многочисленными и хорошо сохранившимися раковинами конхострак, как целыми раковинами и створками, так и раздавленными, а также их обломками, так что некоторые раковины немного выступают на его поверхности (рис. 1, а, б). При извлечении из вмещающей породы копролит раскололся почти пополам, а край его относительно узкого конца слегка обломался. Слом проходит почти по средней плоскости вдоль копролита. Поверхность копролита несет следы изменений в субаэральных условиях – небольшие трещины усыхания и неровности. На поверхности хорошо различимы ходы беспозвоночных (рис. 1, а, б). Представлены два типа ходов: 1) крупные, около 1–1.2 мм в поперечнике и 2) мелкие – около 0.3 мм в поперечнике. Для последних местами наблюдается вхождение их внутрь копролита в виде округлых норок; на сломе копролита также можно наблюдать округлый ход (рис. 1, г). Очевидно, эти ходы и норки образованы различными копрофагами или их личинками разного размерного класса.
Рис. 1.
Копролит морфотипа “F”, экз. ПИН, № 5078/10: a, б – вид снаружи с противоположных сторон; в, г – поверхности продольного слома с раковинами конхострак; д – фрагмент поверхности продольного слома с раковинами конхострак; Владимирская обл., г. Вязники, местонахождение Быковка; верхняя пермь, вятский ярус, верхневятский подъярус.
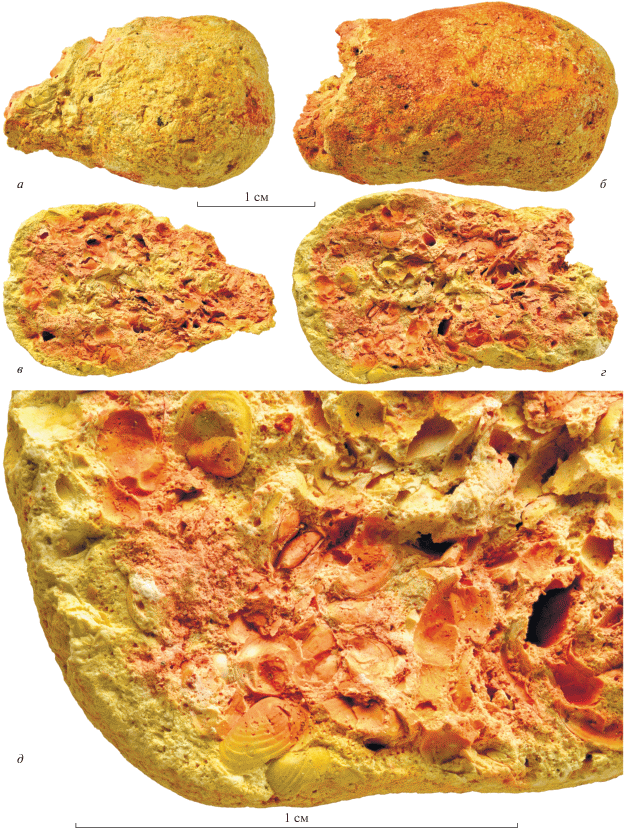
Рис. 2.
Раковины конхострак Pseudestheria cf. chatangensis (Novozhilov, 1946) в копролите морфотипа “F”, экз. ПИН, № 5078/10. Фотографии сделаны на СЭМ Tescan Vega XMU.
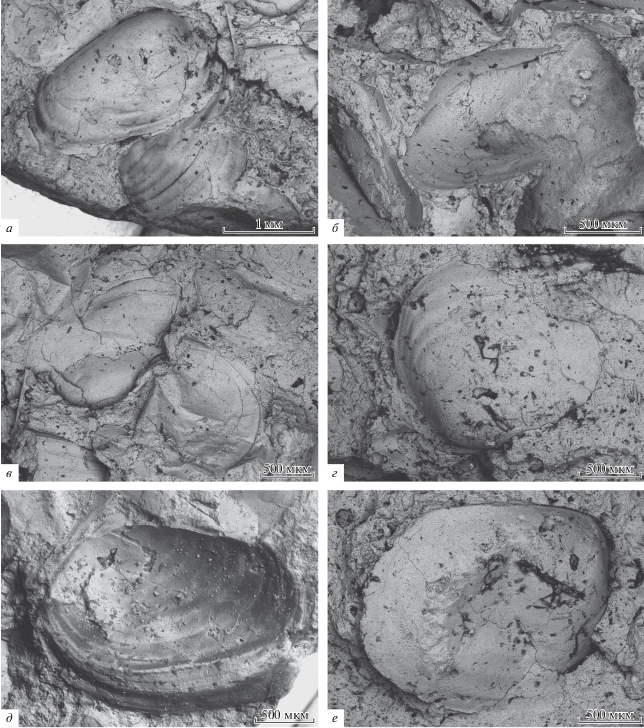
Копролит, целиком заполненный раковинами конхострак, впервые встречен в пермо-триасовых отложениях Восточной Европы.
КОНХОСТРАКИ ИЗ МЕСТОНАХОЖДЕНИЯ БЫКОВКА И ИХ СТРАТИГРАФИЧЕСКОЕ ЗНАЧЕНИЕ
Заполняющие описываемый копролит (экз. ПИН, № 5078/10) раковины и фрагменты раковин конхострак несколько различаются по степени сохранности, но в целом детальное изучение и описание их скульптуры затруднительно. Кроме того, даже целые створки раковин на поверхности слома копролита выступают из вмещающей массы в большей или в меньшей степени и могут наблюдаться в разных положениях, что также затрудняет их измерение и описание.
Благодаря тому, что в ряде верхнепермских и нижнетриасовых местонахождений в окрестностях Вязников и Гороховца конхостраки хорошей сохранности представлены в большом количестве (Scholze et al., 2015, 2019), имеется возможность сравнить остатки конхострак в рассматриваемом копролите с этими материалами. В вышеупомянутых публикациях подробно рассматриваются вопросы таксономии изучаемых родов и видов. Для верхней части вятского яруса в этих разрезах характерен род Pseudestheria Raymond, 1946, полностью исчезающий на границе перми и триаса и сменяющийся другими родами конхострак (Scholze et al., 2015, 2019). Доминирующей формой в верхневятских отложениях (нефедовский и жуковский горизонты) в окрестностях Вязников и Гороховца является P. cf. chatangensis (Novozhilov, 1946).
Заполняющих описываемый копролит конхострак по форме, общим пропорциям и, насколько можно судить, по скульптуре створок также можно определить как Pseudestheria sp., а раковины наилучшей сохранности, для которых есть возможность сделать измерения, – как P. cf. chatangensis (Novozhilov, 1946) (рис. 2). Соотношение длины (L) и высоты (H) створок для них следующее:
L = 2.1–2.4 мм
H = 1.5–1.7 мм
H/L = 0.63–0.76
Для сравнения для всей выборки створок P. chatangensis (Novozhilov, 1946) из позднепермских отложений в окрестностях Вязников и Гороховца измерения следующие (Scholze et al., 2019):
L = 2.2–4.1 мм
H/L = 0.63–0.80
В описаниях P. chatangensis (Novozhilov, 1946) упомянуты и еще более крупные экземпляры (Novozhilov,1958; Defretin-Lefranc, 1965):
L = 2.1–5.6 мм
H = 1.6–4.1 мм
H/L = 0.67–0.74
Таким образом, все определимые раковины конхострак из копролита (ПИН, № 5078/10) можно отнести к P. cf. chatangensis. По своим размерам и пропорциям все раковины конхострак в копролите укладываются в диапазон изменчивости створок P. chatangensis (Novozhilov, 1946), однако представлены лишь мелкими экземплярами. Для объяснения этого факта можно предположить, что выборка из описываемого копролита соответствует моменту их поимки продуцентом копролита в сезон размножения и формирования новой генерации в популяции конхострак, когда в данном месте были представлены почти исключительно мелкие ювенильные особи. Альтернативное объяснение – выборочность по размерному классу в питании конхостраками продуцента копролита – кажется крайне маловероятной. Не исключено, что конхостраки в описываемом копролите представлены хотя и взрослыми, но мелкими экземплярами, небольшой размер которых можно было бы объяснить какими-то не вполне благоприятными условиями существования.
Упомянутую выше раковину конхостраки плохой сохранности в копролите из местонахождения Быковка (Owocki et al., 2012) также можно определить как Pseudestheria sp.
ВОЗМОЖНЫЙ ПРОДУЦЕНТ КОПРОЛИТА С КОНХОСТРАКАМИ ИЗ МЕСТОНАХОЖДЕНИЯ БЫКОВКА
В местонахождении Быковка (с учетом материалов из раскопок 2018 г.) определены следующие тетраподы: сеймуриаморфы (котлассииды) Microphon sp. nov., хрониозухии (хрониозухиды) Uralerpeton tverdochlebovae Golubev, 1998, брахиопоиды (двинозавриды) Dvinosaurus sp., парейазавры Elginiidae gen. indet., протерозухии (протерозухиды) Archosaurus rossicus Tatarinov, 1960, дицинодонты Dicynodontinae gen. indet., тероцефалы мосховайтсииды Moschowhaitsia (?) sp. и наниктидопиды Purlovia (?) sp., Cynodontia gen. indet. Здесь известны также рыбы Toyemia blumentalis A. Minich, 1995, Toyemia sp., Strelnia sp., Isadia sp., Evenkia (?) sp. Кроме того, найдены единичные зубы, ранее соотносимые с Saurichthys (?) sp. Последние, вероятно, принадлежат представителям нового таксона рыб (Миних и др., 2014; Lebedev et al., 2015).
Морфотип “F”, к которому принадлежит рассматриваемый копролит, ранее был соотнесен с парарептилиями или амфибиями (скорее всего, сеймуриаморфными) (Niedźwiedzki et al., 2016). Парарептилии в вязниковских местонахождениях представлены единичными находками покровных костей растительноядных Elginiidae (Буланов, Яшина, 2005); таким образом, наиболее вероятным продуцентом копролита является одна из форм амфибий. Кроме вышеперечисленных земноводных, обнаруженных в Быковке, к числу вероятных продуцентов могут быть также отнесены определенные из других вязниковских местонахождений Karpinskiosauridae (Seymouriamorpha) и Microsauria.
Более точному определению продуцента может способствовать необычное заполнение копролита исключительно раковинками конхострак: активное (не исключено – факультативное) питание объектами с требующими раздавливания плотными покровами подразумевает наличие соответствующих адаптаций в строении зубной системы и челюстного аппарата.
Для хрониозухий возможность питания мелкоразмерными раковинными объектами крайне маловероятна ввиду их крупного размера и очевидных адаптаций к облигатной ихтиофагии, выражающихся в сильно удлиненном ростральном отделе черепа, большом количестве остроконических челюстных (и такой же формы увеличенных небных) зубов, гомодонтном характере построения маргинального озубления в целом (Ивахненко, Твердохлебова, 1980; Голубев, 2000; Witzmann, Schoch, 2018).
Обычно ихтиофагия как основная трофическая специализация подразумевается для неотенических брахиопоидов рода Dvinosaurus (Ивахненко, 2001), в целом наследующих архитектуру зубной системы типичных рыбоядных темноспондил (предковых Trimerorhachidae), в том числе фиксирующие клыковые пары на нижней челюсти и латеральных элементах небного комплекса. С этим положением, однако, плохо согласуются данные тафономического анализа: в отличие от Chroniosuchia, в известных местонахождениях пермских тетрапод находки описанных видов Dvinosaurus не имеют прямой количественной корреляции с остатками сопутствующей ихтиофауны (Голубев, 2009), что позволяет трактовать Dvinosauridae как водных хищников с широким спектром трофических преференций, к которым, однако, исходя из особенностей организации зубной системы, вряд ли могут быть отнесены мелкие раковинные объекты.
Облигатное питание мелкими беспозвоночными с плотными наружными покровами маловероятно также для Karpinskiosauridae. У видов рода Karpinskiosaurus крючковидная форма коронок маргинальных зубов свидетельствует о фиксации активной добычи, в качестве которой могла выступать молодь рыб или сравнительно крупные подвижные беспозвоночные (Буланов, 2006). Строение челюстного сустава K. secundus, допускавшего продольное скольжение articulare относительно иммобилизованного в черепную конструкцию quadratum, позволяло осуществлять проталкивание добычи внутрь ротовой полости за счет ретракционных движений нижней челюсти (Буланов, 2016).
Одной из типичных составляющих вязниковской фауны тетрапод являлись лепоспондилы отряда Microsauria. Первые материалы по этой группе из терминальнопермских отложений района Вязников в виде многочисленных элементов посткраниума были получены Б.П. Вьюшковым в 1955–1956 гг. при раскопках ныне недоступного для изучения местонахождения Вязники-1 (Шишкин и др., 2018) и трактовались как остатки рептилий неясного систематического положения (Вьюшков, 2018). Строение зубной системы описанного разнообразия микрозавров (Carroll, Gaskill, 1978) позволяет констатировать для многих видов питание мелкими беспозвоночными с плотными покровами, однако остатки этой группы в местонахождении Быковка пока не обнаружены. Кроме того, размер копролита ПИН, № 5078/10 достаточно велик для небольших по размеру вязниковских микрозавров.
Наиболее вероятным продуцентом является представитель нового вида рода Microphon (Kotlassiidae, Seymouriamorpha), описание которого вторым автором находится в печати. Положение crista paraquadrata на чешуйчатых костях, слабое развитие покровного орнамента и некоторые особенности строения челюстных зубов позволяют сближать данную форму с Microphon gracilis.
Как и у Karpinskiosauridae, челюстные зубы нового вида Microphon имеют отчетливый изгиб вершинок внутрь ротовой полости; зубы такого типа могли осуществлять эффективное удержание подвижной добычи, но мало пригодны для успешного манипулирования и раздавливания округлых раковин конхострак. Тем не менее, можно предполагать, что, как и других видов Microphon, зубная система нового вида представляла собой комплексное специализированное образование, позволявшее потреблять значительно более широкий спектр трофических объектов (Bulanov, 2003).
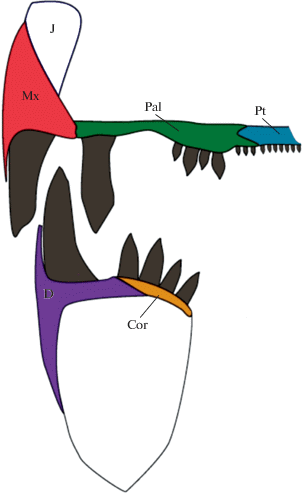
Так, зубная система предполагаемого предкового таксона (M. gracilis) была способна обеспечивать питание как растительным ресурсом (главным образом за счет стригущих движений поликуспидных маргинальных зубов преорбитального отдела черепа), так и склерированными или тонкораковинными беспозвоночными (Буланов, 2006), раздавливаемыми при совокупном участии маргинальных боковых зубов, увеличенных зубов небного комплекса и зубов короноидной серии на участке между хоаной и аддукторной полостью (рис. 3). Последнее подтверждается сильной прижизненной деструкцией как челюстных, так и небных зубов у M. gracilis, которая проявляется уже на ранних стадиях онтогенеза (длина черепа менее 40 мм; Bulanov, 2003). Следует также отметить, что в типовом местонахождении остаткам M. gracilis (Бабинцево, Оренбургская обл.) сопутствуют многочисленные раковины конхострак.
Рис. 3.
Схема организация зубной системы позднепермского котлассиида Microphon gracilis Bulanov, 2003; поперечное сечение озубленных элементов верхнечелюстной дуги, неба и нижней челюсти на уровне передней половины орбиты. Обозначения: Cor – сoronoideum, D – dentale, J – jugale, Mx – maxillare, Pt – pterygoideum, Pal – palatinum.
ЗАКЛЮЧЕНИЕ
Впервые в пермо-триасовых отложениях Восточной Европы встречен копролит, целиком заполненный раковинами конхострак. Его наиболее вероятным продуцентом является котлассиидный сеймуриаморф нового вида рода Microphon, представленный в том же местонахождении многочисленными изолированными остатками. Находка свидетельствует в пользу предположений о принадлежности копролитов морфотипа “F” из вязниковских местонахождений сеймуриаморфным амфибиям (Niedźwiedzki et al., 2016) и о присутствии конхострак в рационе котлассиид рода Microphon (Буланов, 2006). Состав комплекса конхострак указывает на терминальнопермский возраст местонахождения Быковка и, тем самым, дает дополнительное независимое биостратиграфическое подтверждение наиболее высокого положения этого местонахождения в пермской части разреза Вязников. Смена фаун тетрапод, рыб и конхострак происходила в данном регионе синхронно и приходится на границу перми и триаса.
Работа выполнена при финансовой поддержке РФФИ в рамках научных проектов №№ 17-04-00410, 17-04-01937, 17-54-10013, а также за счет средств субсидии, выделенной в рамках государственной поддержки Казанского (Приволжского) федерального университета в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров.
Список литературы
Буланов В.В. Трофические адаптации сеймуриаморф (Parareptilia) и положение группы в структуре водных сообществ конца палеозоя // Эволюция биосферы и биоразнообразия. М.: Тов-во научн. изданий КМК, 2006. С. 394–415.
Буланов В.В. Принцип функционирования челюстного сустава у сеймуриаморфных амфибий // Матер. LXII сессии Палеонтологического общества. СПб.: ВСЕГЕИ, 2016. С. 230–231.
-
Буланов В.В., Яшина О.В. Элгиниидные парейазавры Восточной Европы // Палеонтол. журн. 2005. № 4. С. 85–90.
Вьюшков Б.П. Местонахождение пермских наземных позвоночных в окрестностях г. Вязники // Палеонтол. журн. 2018. № 2. С. 58–65.
Голубев В.К. Пермские и триасовые хрониозухии и биостратиграфия верхнетатарских отложений Восточной Европы по тетраподам. М.: Наука, 2000. 174 с. (Тр. Палеонтол. ин-та РАН. Т. 276).
Голубев В.К. Реконструкция трофических связей в позднепермском сообществе тетрапод Восточной Европы // Палеострат-2009. Годичн. собр. секции палеонтологии МОИП и Моск. отделения Палеонтологического об-ва. Москва, 26–27 января 2009 г. Прогр. и тез. докладов. М.: ПИН, 2009. С. 10–11.
Голубев В.К., Миних А.В., Балабанов Ю.П. и др. Опорный разрез перми и триаса в Жуковом овраге у г. Гороховец, Владимирская область // Бюлл. региональной межведомственной стратиграфической комиссии по центру и югу Русской платформы. Вып. 5 / Ред. Алексеев А.С. М.: РАЕН, 2012. С. 49–82.
Ивахненко М.Ф. Тетраподы Восточно-Европейского плакката – позднепалеозойского территориально-природного комплекса. Пермь: Тип. “Книга”, 2001. 200 с.
Ивахненко М.Ф., Твердохлебова Г.И. Систематика, морфология и стратиграфическое значение верхнепермских хрониозухов востока Европейской части СССР. Саратов: Изд-во Саратов. ун-та, 1980. 69 с.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Миних А.В., Миних М.Г., Андрушкевич С.О. Ихтиофауна терминальной перми в окрестностях г. Вязники Владимирской области // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2014. Т. 14. Вып. 2. С. 91–96.
Сенников А.Г. Ранние текодонты Восточной Европы. М.: Наука, 1995. 142 с. (Тр. Палеонтол. ин-та РАН. Т. 263).
Сенников А.Г. Новые данные по синэкологии вязниковского наземного сообщества (терминальная пермь, Центральная Россия) // Палеонтол. журн. 2017. № 4. С. 65–77.
Сенников А.Г., Голубев В.К. Вязниковская фауна: черты экологического кризиса // Природа. 2006. № 7. С. 39–48.
Сенников А.Г., Голубев В.К. К фаунистическому обоснованию границы перми и триаса в континентальных отложения Восточной Европы. 1. Гороховец–Жуков овраг // Палеонтол. журн. 2012. № 3. С. 88–98.
Сенников А.Г., Голубев В.К. Последовательность пермских фаун тетрапод Восточной Европы и пермо-триасовый экологический кризис // Палеонтол. журн. 2017. № 6. С. 30–41.
Шишкин М.А., Сенников А.Г., Голубев В.К. О статье Б.П. Вьюшкова “Местонахождение пермских наземных позвоночных в окрестностях г. Вязники” // Палеонтол. журн. 2018. № 2. С. 66–77.
Bajdek P., Qvarnsröm M., Owocki K. et al. Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia // Lethaia. 2016. V. 49. № 4. P. 455–477.
Bajdek P., Owocki K., Niedźwiedzki G. et al. Residues from the Upper Permian carnivore coprolites from Vyazniki in Russia – key questions in reconstruction of feeding habits // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2017. V. 482. P. 70–82.
Bulanov V.V. Evolution and systematics of seymouriamorph parareptiles // Paleontol. J. 2003. V. 37. Suppl. 1. 105 p.
-
Carroll R.L., Gaskill P. The order Microsauria // Mem. Amer. Philos. Soc. Philad. 1978. V. 126. 211 p.
Defretin-Lefranc S. Etude et révision de Phyllopodes Conchostracés en provenance d’U.R.S.S // Ann. Soc. Géol. Nord. 1965. V. 85. P. 15–48.
Lebedev O.A., Sennikov A.G., Golubev V.K. et al. The first find of Permian ceratodontids (Dipnoi, Osteichthyes) in Russia // Paleontol. J. 2015. V. 49. № 10. P. 1112–1124.
Niedźwiedzki G., Bajdek P., Qvarnström M. et al. Reduction of vertebrate coprolite diversity in association to the end-Permian extinction event in Vyazniki region, Central Russia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2016. V. 450. P. 77–90.
Novozhilov N. Conchostraca du Permien et du Trias du littoral de la Mer des Laptev et de la Toungounzka inférieure // Ann. Serv. Inform. Géol. Bureau Rech. Geol., géophys., minières. 1958. V. 26. P. 15–80.
Owocki K., Niedźwiedzki G., Sennikov A.G. et al. Upper Permian vertebrate coprolites from Vyazniki and Gorokhovets, Vyatkian regional stage, Russian platform // Palaios. 2012. V. 27. № 12. P. 867–877.
Scholze F., Golubev V.K., Niedzwiedzki G. et al. Early Triassic Conchostracans (Crustacea: Branchiopoda) of the terrestrial Permian-Triassic boundary sections in the Moscow syncline // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 429. P. 22–40.
Scholze F., Golubev V.K., Niedźwiedzki G. et al. Late Permian conchostracans (Crustacea, Branchiopoda) from continental deposits in the Moscow syneclise, Russia // J. Paleontol. 2019. V. 93. № 1. P. 72–97.
Sennikov A.G. Evolution of the Permian and Triassic tetrapod communities of Eastern Europe // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1996. V. 120. № 3–4. P. 331–351.
Sennikov A.G., Golubev V.K. Unique Vyazniki biotic complex of the terminal Permian from the central Russia and the global ecological crisis at the Permo-Triassic boundary // New Mexico Museum Natur. Hist. Sci. Bull. 2005. № 30. P. 302–304.
Sennikov A.G., Golubev V.K. Vyazniki biotic assemblage of the terminal Permian // Paleontol. J. 2006. V. 40. Suppl. 4. P. 475–481.
Witzmann F., Schoch R.R. Skull and postcranium of the bystrowianid Bystrowiella schumanni from the Middle Triassic of Germany, and the position of chroniosuchians within Tetrapoda // J. Syst. Palaeontol. 2018. V. 16. № 9. P. 711–739.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал