

Палеонтологический журнал, 2020, № 1, стр. 78-88
Раннеплейстоценовые винторогие антилопы (Artiodactyla, Bovidae) из пещеры Таврида (Крым, Россия)
И. А. Вислобокова a, *, В. В. Титов b, А. В. Лавров a, Д. О. Гимранов c, d, Д. Б. Старцев e, К. К. Тарасенко a
a Палеонтологический институт им. А.А. Борисяка РАН, Москва, Россия
b Южный научный центр РАН, Ростов-на-Дону, Россия
c Институт экологии растений и животных УрО РАН, Екатеринбург, Россия
d Уральский федеральный университет, Екатеринбург, Россия
e Крымский федеральный университет им. В.И. Вернадского, Симферополь, Россия
* E-mail: ivisl@paleo.ru
Поступила в редакцию 05.02.2019
После доработки 07.02.2019
Принята к публикации 12.02.2019
Аннотация
Дано описание ископаемых остатков двух видов винторогих антилоп Gazellospira torticornis (Aymard, 1854) и Pontoceros ambiguus Vereschagin, Alexejeva, David et Baigusheva, 1971, найденных в 2018 г. в нижнеплейстоценовых отложениях пещеры Таврида в Центральном Крыму. Эти находки – первое свидетельство присутствия этих таксонов на территории полуострова. Данная ассоциация коррелируется с поздневиллафранкскими фаунами Европы и зоной млекопитающих MNQ18 (в интервале 1.8–1.5 млн л. н.). G. torticornis из местонахождения Таврида подтверждает тенденцию к уменьшению размеров поздних представителей антилоп этого вида. P. ambiguus из Тавриды – одни из наиболее ранних представителей вида на юге Европы. Находки винторогих антилоп в Тавриде важны для уточнения особенностей их морфологической эволюции, географического и стратиграфического распространения.
Местонахождение плейстоценовых позвоночных пещера Таврида открыто в 2018 г. в Центральном Крыму в районе пос. Зуя в 15 км к востоку от Симферополя при прокладке федеральной автомобильной трассы Симферополь–Керчь. Особый интерес представляет изучение найденных в пещере остатков млекопитающих псекупского комплекса, относящихся ко времени раннего расселения гоминин рода Homo в Евразии.
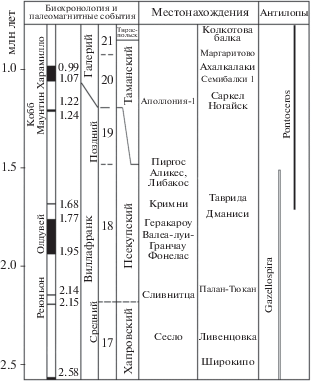
Пещера Таврида находится на Внутренней гряде Крымских гор в междуречье рек Бештерек и Фундуклы и заложена в палеогеновых известняках. Костеносный слой в южной галерее пещеры представлен красно-бурыми суглинками субаэрального типа. Из точки “Грот гиен” пещеры происходят остатки зайца Hypolagus brachygnathus (Kormos, 1930), дикобраза Hystrix (Acanthion) vinogradovi Argyropulo, 1941, мелкого волка Canis sp., гигантской гиены Pachycrocuta brevirostris (Gervais, 1850), саблезубой кошки Homotherium crenatidens (Fabrini, 1890), меридионалоидного слона Archidiskodon meridionalis (Nesti, 1825), двух таксонов стеноноподобных лошадей, носорогов Elasmotherium sp. и Stephanorhinus sp., верблюда Paracamelus gigas Schlosser, 1903, оленя Arvernoceros verestchagini David, 1992, бычьих Leptobos sp. и Bison (Eobison) sp., а также двух винторогих антилоп. Предварительный анализ биохронологических данных показывает, что фауна Тавриды относится к псекупскому комплексу млекопитающих Восточной Европы, позднему виллафранку, зоне млекопитающих MNQ18 в интервале 1.8–1.5 млн л. н. (Лопатин и др., 2019). По данным изотопного (40Ar/39Ar) анализа материалов из местонахождений Франции, диапазон зоны MNQ18 примерно соответствует интервалу от палеомагнитного субхрона Реюньон, 2.19–2.14 млн л. н. (фауна Сенез) до 1.47 млн л. н. (фауна Пейроль, MNQ19) (см. Nomade et al., 2014).
Изучение разных групп Bovidae становится одним из актуальных направлений исследований эволюции плейстоценовых фаун, так как палеонтологические и археологические данные позволяют предполагать связь миграций этих животных и гоминин. В данной статье мы приводим первое описание винторогих антилоп из пещеры Таврида. Диагностика этих антилоп основывается на особенностях строения роговых стержней и типе их скручивания (гомонимном или гетеронимном), а также особенностях костей посткраниального скелета. Ископаемый материал хорошей сохранности из “Грота гиен” свидетельствуют о присутствии в фауне Тавриды двух видов винторогих антилоп, относящихся к родам Gazellospira Pilgrim et Schaub, 1939 и Pontoceros Vereschagin, Alexejeva, David et Baigusheva, 1971. Находка винторогой антилопы рода Pontoceros в пещере Таврида представляет особый интерес, так как, вероятно, отражает первое появление этого рода в раннем плейстоцене Европы. Pontoceros близкого возраста известен из местонахождения Дманиси в Грузии (Vekua, 2012), в котором присутствуют остатки ранних Hоmо (Lordkipanidze et al., 2013). Кроме того, Таврида – одно из немногих местонахождений, в котором найдены ископаемые остатки поздних представителей характерного для виллафранка Европы и Турции вида Gazellospira torticornis (Aymard, 1854) и первое местонахождение этого вида на юге Восточной Европы.
В работе приняты следующие обозначения для промеров роговых стержней и метаподий: длина (L), поперечный диаметр (DT), переднезадний диаметр (DAP), проксимальный (prox), дистальный (dist), диафиз (diaph).
Используются следующие сокращения: ПИН – Палеонтологический ин-т им. А.А. Борисяка РАН, ИЭРЖ – Ин-т экологии растений и животных УрО РАН.
Исследование частично выполнено за счет средств Программы фундаментальных исследований Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов”, а также в рамках научного проекта РФФИ № 19-05-00982 и реализации госзадания ЮНЦ РАН на 2019 г., № проекта 01201363186. Авторы выражают благодарность всем участникам раскопок местонахождения Таврида.
Gazellospira torticornis (Aymard, 1854). Материал. Фрагмент правого рогового стержня, ПИН, № 5644/57; фрагмент основания правого рогового стержня с частью лобной кости, ИЭРЖ, № 727/47; правая пястная кость, ИЭРЖ, № 727/44; правая плюсневая кость, ИЭРЖ, № 727/45.
Описание и сравнение (рис. 1). Роговые стержни гетеронимно скручены примерно на половину оборота у экз. ПИН, № 5644/57, с двумя спиральными килями, передним и задненаружным, последний развит сильнее. Сечение рогового стержня в основании у экз. ИЭРЖ, № 727/47 почти округлое, слегка заостренное у задненаружного киля; сечение рогового стержня экз. ПИН, № 5644/57 – эллипсовидное (слабо овальное). Промеры в мм: экз. ИЭРЖ, № 727/47: L – 108, DT × DAP –27.9 × 28.7.
Рис. 1.
Gazellospira torticornis (Aymard, 1854): а–в – ПИН, № 5644/57, неполный правый роговой стержень: а – спереди, б – изнутри, в – сзади; г, д – ИЭРЖ, № 727/47, фрагмент рогового стержня: г – спереди, д – сзади; е–з – ИЭРЖ, № 727/44, правая пястная кость: е – спереди, ж – сзади, з – сверху; и–л – ИЭРЖ, № 727/45, плюсневая кость: и – спереди, к – сзади, л – сверху; Крым, пещера Таврида; нижний плейстоцен.
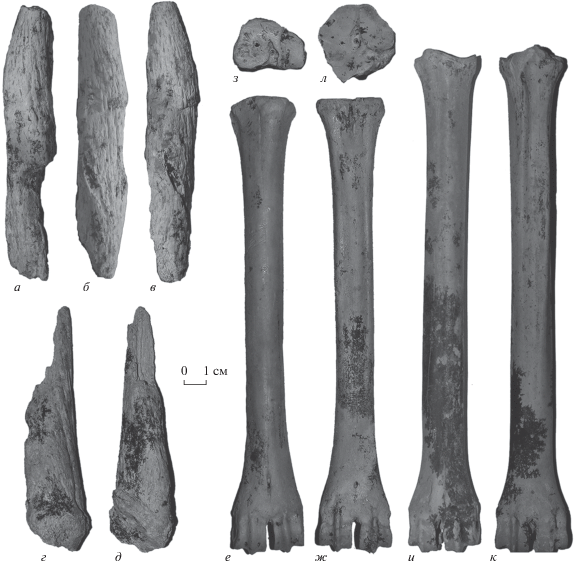
Пястная и плюсневая кости тонкие, стройные, постепенно расширяющиеся к проксимальному и дистальному эпифизам. Промеры пястной кости ИЭРЖ 727/44 в мм: L – 219, DTprox × DAPprox – 32.5 × 23.7, DTdist × DAPdist – 34.7 × 22.9, DTdiaph × DAPdiaph 18.7 × 17.1; индекс массивности (DTdiaph/L) – 0.09; индекс ширины дистального эпифиза (DTdist/L) – 0.14. Промеры плюсневой кости ИЭРЖ 727/45 в мм: L – 247, DTprox × DAPprox – 31 × 31.7, DTdist × DAPdist – 35 ×26.2, DTdiaph × DAPdiaph – 19.5 × 22.9 мм; индекс DTdiaph/L – 0.08; индекс DTdist/L – 0.14.
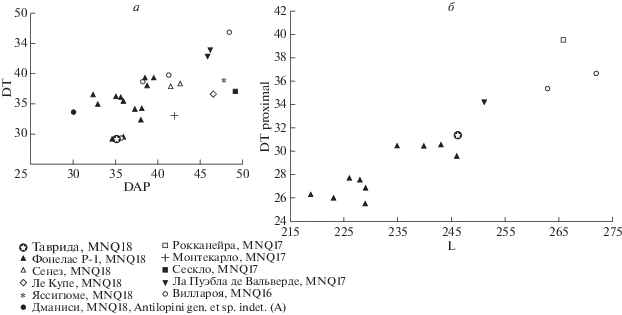
Морфология и размеры роговых стержней, а также пястной и плюсневой костей этой антилопы из Тавриды соответствуют таковым G. torticornis из позднего виллафранка Европы (рис. 2). По промерам диаметров DT и DAP основания роговых стержней антилопа из Тавриды близка к наиболее мелким формам G. torticornis. Судя по длине и пропорциям пястной и плюсневой костей, G. torticornis из Тавриды была сравнительно небольшой и изящной. Промеры длины этих костей у нее близки к максимальным величинам у G. torticornis hispanica Garrido, 2008, мелкой формы из местонахождения Фонелас P-1 (~2 млн л. н., MNQ18) в Испании. По значениям индексов DTdiaph/L и DTdist/L G. torticornis из Тавриды и Фонеласа P-1 тоже примерно совпадают. У голотипа из Фонеласа P-1: L – 235, DTprox × DAPprox – 30.49 × 33.31, DTdist × DAPdist – 32.47 × 22.71, DTdiaph – 16.96; индекс DTdiaph/L – 0.07; индекс DTdist/L – 0.14; размах изменчивости длины пястной кости у антилопы из Фонеласа P-1 (L min–max) – 219–246 (Garrido, 2008). От G. gromovae Dmitrieva, 1975 из Куруксая (MNQ17), Таджикистан (Дмитриева, 1977), отличается более тонкими роговыми стержнями и более мелкими общими размерами.
Рис. 2.
Соотношение параметров роговых стержней (а) и плюсневых костей (б) у Gazellospira torticornis (Aymard, 1854) (c использованием данных: Athanassiou, 2005; Bukhsianidze, 2005; Garrido, 2008; Rodrigo, 2011; Demirel, Mayda, 2014; Bellucci, Sardella, 2015; и др.).
Замечания. Вид G. torticornis довольно полно представлен в ископаемом состоянии в Западной Европе и хорошо изучен (Pilgrim, Schaub, 1939; Heintz, 1966, 1970; Duvernois, Guérin, 1989; Kostopoulos, 1997a; Spassov, Crégut-Bonnoure, 1999; Athanassiou, 2005; Lyras, van der Geer, 2007; Garrido, 2008; Bellucci, Sardella, 2015; и др.). Голотип типового вида G. torticornis – правый рог на фрагменте черепа из местонахождения начала позднего виллафранка Ле Купе (Музей Крозатье, Лэ Пуи-эн-Вэлей, Франция; изображение см.: Dorlhac, 1854, рис. 8 ). Этот вид характеризуется гетеронимным типом скручивания рогов. Лобные кости пневматизированы, роговые стержни без полостей пневматизации (синусов), сидят над глазницами, свернуты в слабый завиток, с двумя спиральными килями, поперечное сечение роговых стержней слабо овальное и округлое, заостренное у заднего, сильнее развитого киля. Исследования показали, что роговые стержни у антилоп этого вида могут различаться как по длине, так и по степени развития килей, а также по числу их оборотов вокруг продольной оси рога. Западноевропейский материал позволил предполагать постепенное уменьшение размеров G. torticornis в виллафранке Испании и Франции (Heintz, 1966; Rodrigo, 2011). Уменьшение общих размеров наиболее заметно у антилоп начала позднего виллафранка, в частности, из местонахождения Фонелас P-1 (~2 млн л. н., MNQ18) в Испании (Garrido, 2008). Популяция G. torticornis из Фонеласа P-1 – одна из самых поздних в Средиземноморье. Она выделена в подвид G. torticornis hispanica Garrido, 2008. Голотипом этого подвида является левая плюсневая кость, FP1-2001-0165, хранящаяся в Музее Ин-та геологии и минералогии Испании в Мадриде (Garrido, 2008, рис. 16F ), а паратипы представлены фрагментами черепов с роговыми стержнями, верхней и нижней челюстью, лучевой и пястной костями. Диагностические признаки подвида включают особенности роговых стержней, зубов и небольшие размеры костей посткраниального скелета. У этого подвида роговые стержни достигают длины около 300 мм, наклонены под углом 35°–60° к крыше черепа и расположены на расстоянии 53–63 мм друг от друга. Угол расхождения рогов – 64°–76°. У основания рогового стержня хорошо развит заднелатеральный киль и иногда выражен передний киль; степень развития этих килей и их положение изменчивы; число полных оборотов килей вокруг продольной оси рога может доходить до двух (Garrido, 2008).
Из местонахождения Куруксай (Таджикистан), коррелируемого с европейскими средневиллафранкскими фаунами, описан другой вид, G. gromovae Dmitrieva, 1975. Эта форма отличается от G. torticornis меньшим расхождением роговых стержней в основании, а также некоторыми особенностями основания черепа и верхних зубов (Дмитриева, 1975, 1977).
Данные из пещеры Таврида свидетельствуют о существовании довольно мелкой формы G. torticornis в Крыму, что соответствует тренду уменьшения размеров G. torticornis в начале позднего виллафранка (MNQ18), прослеживающемуся на материалах из Западной Европы.
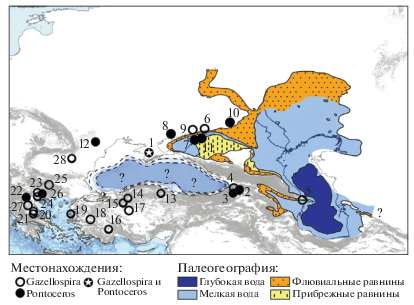
Самые древние Gazellaspira, возможно, присутствуют в плиоценовом местонахождении Галта (MN15) в Турции (Bouvrain, 1998). Достоверные находки представителей этого рода антилоп известны из позднеплиоценовых местонахождений, коррелируемых с зоной MN16b, в частности, Вилларои в Испании (см. Rodrigo, 2011), Квабеби в Грузии (Bukhsianidze, Koiava, 2018) и Гюльязи в Турции (Sickenberg, 1974). В раннем плейстоцене род Gazellaspira был распространен в европейской и азиатской частях Средиземноморской зоогеографической подобласти Палеарктики, а также в некоторых прилегающих регионах. В общем, биохронологический диапазон вида G. torticornis – зоны MN16b–MNQ18. Наиболее широко данный вид был представлен в среднем и в начале позднего виллафранка. В среднем виллафранке (MNQ17) он обитал на территории от Испании до Турции. На рубеже среднего и позднего виллафранка ареал вида начинает сокращаться, и в постолдувейское время в конце дуная/эбурония (начало зоны MNQ19) этот вид исчезает. В Италии последние представители G. torticornis известны из фаун Коста-Сан-Джакомо (MNQ17) (Bellucci, Sardella, 2015). К первой половине позднего виллафранка (MNQ18) относятся находки этого вида в местонахождениях Франции (Ле Купе, Шиллак, Сенез), Испании (Фонелас P-1), Греции (Геракароу, Кримни 1, Аликес, возможно, Пиргос, и др.), Германии (Эрпфингер Хехле), Румынии (Валя-луи-Гранчану), Болгарии (Сливнитца), Черногории (Трлица, TRL11–10) и Турции (Яссигюме, 1.5 млн л. н.) и др. (Kostopoulos, Koufos, 1994; Athanassiou, 1996, 2005; Kostopoulos, 1997a, 2014; Spassov, Crégut-Bonnoure, 1999; Koufos, 2001; Kostopoulos et al., 2002; Spassov, 2003, 2016; Rădulescu et al., 2003; Arribas et al., 2004; Radulesko, 2005; Crégut-Bonnoure, 2007; Garrido, 2008; Rodrigo, 2011; Demirel, Mayda, 2014; Вислобокова, Агаджанян, 2015; Koufos, Kostopoulos, 2016; и др.). Находка G. torticornis в пещере Таврида в Крыму – первая в Восточной Европе в составе фауны позднего виллафранка. Из этого региона антилопы рода Gazellospira были известны лишь из местонахождений хапровского комплекса Ливенцовка в Приазовье и Болград в Одесской области, относящихся к среднему виллафранку (MNQ17), но материал по ним очень скуден. Этих газеллоспир относили к G. torticornis (Алексеева, 1977) или к G. gromovae (Титов, 2008). По данным М. Букхсианидзе, среди антилоп в местонахождении Дманиси (1.75 млн л. н., MNQ18; Грузия), возможно, есть и Gazellospira (Bukhsianidze, 2005). По размерам и пропорциям пястные кости Antilopini gen. et sp. indet. (B) из Дманиси близки к G. torticornis из средневиллафранкских местонахождений (зона MNQ17) Рокканейра (Испания) и Сенез (Франция) (Bukhsianidze, 2005). Отсутствие находок роговых стержней Gazellospira в Дманиси пока оставляет открытым вопрос о присутствии там этих антилоп.
Pontoceros ambiguus Vereschagin, Alexejeva, David et Baigusheva, 1971. Материал. Фрагмент крыши черепа с частью правого рогового стержня, ПИН, № 5644/54; неполный правый роговой стержень, ПИН, № 5644/55; неполный левый роговой стержень, ИЭРЖ, № 727/46; правая пястная кость, ИЭРЖ, № 727/43; дистальный конец плюсневой кости, ПИН, № 5644/58.
Описание и сравнение (рис. 3). Роговые стержни сидят над орбитами. Они гомонимно скручены, трехгранные. Синусы в их основании отсутствуют. Экз. ПИН, № 5644/55 по морфологии и размерам хорошо соответствует верхней половине голотипа вида P. ambiguus из Ногайска, Украина. Хорошо развиты три киля, названные вслед за авторами вида по положению их проксимальных точек (начала килей) у голотипа, а именно: фронтальный (переднемедиальный), заднелатеральный и заднемедиальный. Последний киль уплощен и имеет форму валика, а переднемедиальный и заднелатеральный кили хорошо выражены и сильнее заострены. Рог от основания к вершине закручен примерно на один оборот, у голотипа – на 1.25 оборота. На передненаружной грани рогового стержня экз. ПИН, № 5644/55 вдоль края переднемедиального гребня тянется глубокая бороздка; такая же бороздка на голотипе доходит до основания рога. Поперечное сечение экз. ПИН, № 5644/55 в основании имеет округленно-треугольную форму, близкую к таковой на половине длины рога голотипа; выше сечение становится округленно-треугольным. На экз. ПИН, № 5644/54 роговой стержень расположен над глазницей и наклонен назад под углом около 40° к крыше черепа; в основании роговые стержни, по-видимому, очень слабо расходились в стороны. Промеры роговых стержней в мм: экз. ПИН, № 5644/54 DAP – 46; DT (вычислен) – 48, экз. ПИН, № 5644/55 DAP × DT у нижнего облома – 30 × 42.5; у основания эти промеры могли быть примерно как у голотипа – 46 × 48 или немного больше; экз. ИЭРЖ, № 727/46: L – 177, DT × DAP – 28.6 × 35.3.
Рис. 3.
Pontoceros ambiguus Vereschagin, Alexejeva, David et Baigusheva, 1971: а–в – ПИН, № 5644/55, правый роговой стержень: а – спереди, б – сзади, в – сечение его основания; г – рисунок голотипа ГИН, № 98-1, из Ногайска (по: Верещагин и др., 1971, табл. 34 , фиг. 1), показана форма сечений и линия примерно соответствующая положению основания фрагмента рога из Тавриды; д – ПИН, № 5644/54, фрагмент крыши черепа с частью левого рогового стержня; е, ж – ИЭРЖ, № 727/46, неполный левый роговой стержень: е – снаружи, ж – изнутри; з–к – ИЭРЖ, № 727/43, правая пястная кость: з – сверху, и – спереди, к – сзади; л – ПИН, № 5644/58, дистальный конец плюсневой кости, спереди; Крым, пещера Таврида; нижний плейстоцен.
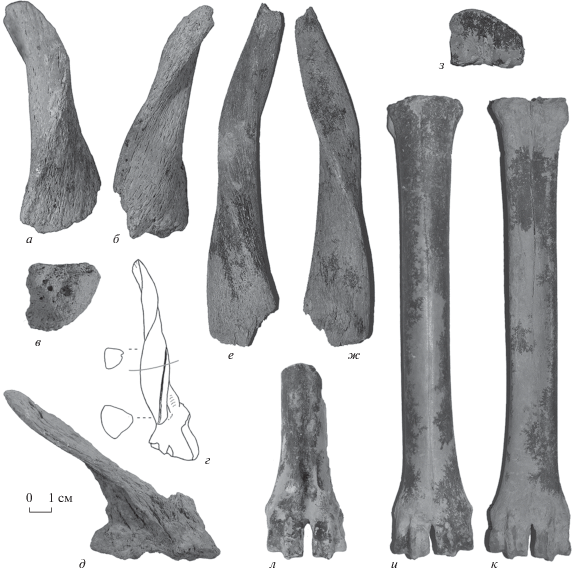
По морфологии, размерам, форме сечения и углу наклона роговые стержни из Тавриды соответствуют голотипу P. ambiguus (рис. 3, г). От P. ambiguus mediterraneus Kostopoulos, 1997 из Аполлонии-1 (Греция; MNQ20, ~1 млн л. н.), отличается более четко выраженной трехгранной формой и немного большими размерами. У P. ambiguus mediterraneus DAP × DT основания рогов – 40.5–41.3 × 43.2–44.6 мм, и их сечение от основания до середины высоты рога эллиптически-субокруглое, так как кили более сглаженные, чем у типичной формы (Kostopoulos, 1997b). Рога у этого подвида наклонены назад под углом 30°–40° к крыше черепа, скручены на один оборот и расходятся в стороны немного сильнее, чем у типичной формы. Кроме того, в передней части лобной кости у P. ambiguus mediterraneus есть многочисленные мелкие синусы, которые отсутствуют у P. ambiguus из Тавриды и Ногайска; усиление пневматизации относится к прогрессивным признакам. От голотипа P. surprine Vekua, 2012 из Дманиси (Грузия; MNQ18, ~1,8 млн л. н.) антилопа из Тавриды отличается немного меньшими размерами, большим наклоном рогов назад, их трехгранной формой и наличием трех килей, а также более мелкими размерами. У голотипа P. surprine стержни массивнее (DT × DAP – 55.5 × 50 мм), повернуты на один оборот; резко развит лишь один (переднелатеральный) киль; рог наклонен назад под углом 50° к крыше черепа; поперечное сечение основания рога неправильно-округленной формы (Vekua, 2012).
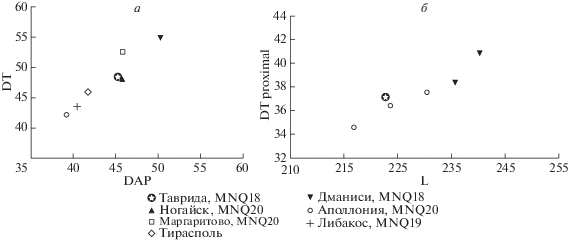
Пястная кость, экз. ИЭРЖ, № 727/44, удлиненная, стройная, но немного более массивная, чем у G. torticornis. По морфологии, размерам и массивности соответствует P. ambiguus (рис. 4). На проксимальной суставной поверхности фасетка для os trapezoideocapitatum почти квадратная, гребень между ней и фасеткой для os hamatum низкий. На пальмарный стороне диафиза желоб sulcus longitudinalis palmaris широкий и длинный, отверстие канала canalis metacarpi distalis маленькое. Расширение проксимальной и дистальной частей диафиза пястной и плюсневых костей к эпифизам довольно резкое; контур дистального эпифиза (при взгляде спереди) напоминает прямоугольник. Дорсальный продольный желоб на плюсневой кости около canalis metatarsi distalis имеет высокие острые гребни. Эти признаки выражены и на метаподиях из Дманиси, которые определены как Antilopini gen. et sp. indet. (B) (Bukhsianidze, 2005, табл. VII -5), но, возможно, принадлежат Pontoceros. Длина этих костей у Antilopini gen. et sp. indet. (B) из Дманиси немного больше, чем у понтоцероса из Тавриды, но по индексам массивности они близки. Промеры пястной кости экз. ИЭРЖ, № 727/43 в мм: L – 223, DTprox × DAPprox – 37.4 × 26.6, DTdist × DAPdist – 39.3 × 25.9; DTdiaph × DAPdiaph – 23.4 × 20; индекс DTdiaph/L – 0.10; индекс DTdist/L – 0.18. Они примерно соответствуют промерам и индексам пястных костей у P. ambiguus mediterraneus. Величины этих индексов у P. ambiguus mediterraneus – 0.08–0.09 и 0.15–0.17, а у Antilopini gen. et sp. indet. (B) – 0.10 и 0.16 (рис. 4, б).
Рис. 4.
Соотношение параметров роговых стержней (а) и пястных костей (б) у антилоп рода Pontoceros (с использованием данных: Верещагин и др., 1971; Kostopoulos, 1997b; Bukhsianidze, 2005; Vekua, 2012).
Замечания. Род Pontoceros был выделен по материалам из Ногайска и Маргаритово в Приазовье, а также Тирасполя (Колкотова Балка) в Молдавии на основании строения рогов (Верещагин и др., 1971). Голотипом типового вида P. ambiguus является правый рог с фрагментом черепа, экз. ГИН № 98-1, из нижнего плейстоцена Ногайска, предположительно из нижних слоев разреза (Верещагин и др., 1971, табл. ХХХIV, фиг. 1–2). По данным Л.И. Алексеевой (1977), эти слои залегают ниже отложений с ногайской фауной мелких млекопитающих (MNQ20) и содержат фауну поздней стадии хапровского (=псекупского) комплекса с типичным Archidiskodon meridionalis (=Mammuthus meridionalis). Роговой стержень из Маргаритово в первоописании рода отнесен к таманскому комплексу (Верещагин и др., 1971; Алексеева, 1977), зона MNQ20, а материал из Тирасполя (Колкотова Балка, MNQ21) – к тираспольскому комлексу. Роговой стержень из Маргаритово, судя по его сохранности, может происходить из слоев с фауной, переходной между таманским и тираспольским комплексами млекопитающих (местонахождение Маргаритово 2; MNQ20/MNQ21; см. Tesakov et al., 2007). P. cf. ambiguus присутствует в фауне Семибалки 1 (MNQ20) таманского фаунистического комплекса в Северо-Восточном Приазовье (Байгушева, 2000).
Род Pontoceros существовал примерно от палеомагнитного эпизода Олдувей до начала среднего плейстоцена. В ископаемом состоянии он представлен гораздо беднее, чем Gazellospira (рис. 5). Ареал вида P. ambiguus простирался от Северо-Восточного Приазовья до Греции и, возможно, Израиля. Antilopini gen. et sp. indet. (cf. Pontoceros ambiguus/Spirocerus sp.) входит в состав фауны Убедии (1.6–1.2 млн л. н; Израиль) (Martínez-Navarro et al., 2012). Остатки антилоп рода Pontoceros присутствуют в фаунах Дманиси (MNQ18) (Vekua, 2012) и Ахалкалаки (0.98–0.78 млн л. н., MNQ20) в Грузии (Bukhsianidze еt al., 2014), а также Либакос (MNQ18), Аполлония-1 (MNQ20) и Платаночори-1 (MNQ20) в Греции (Kostopoulos, 1997а, 2006; Konidaris et al., 2015). Pontoceros из Аполлонии-1 морфологически немного отличается от типичной формы из Ногайска (см. выше), и Д. Костопоулос выделил его в подвид Pontoceros ambiguus mediterraneus, к которому отнес и форму из Либакоса (=Antilopinae gen. indet. sp. A.; Steensma, 1988, рис. 81 ) (Kostopoulos, 1997b).
Рис. 5.
Основные местонахождения винторогих антилоп Восточно-Cредиземноморской провинции. Местонахождения: 1 – Таврида, 2 –Дманиси, 3 –Ахалкалаки, 4 – Квабеби, 5 – Палан-Тюкан, 6 – Ливенцовка, 7 – Маргаритово, 8 – Ногайск, 9 – Широкино, 10 – Саркел, 11 – Семибалки, 12 – Колкотова Балка (Тирасполь), 13 – Камышлы, 14 – Сарикoл Тепе, 15 – Галта, 16 – Яссигюме, 17 – Гюльязи, 18 – Бгек, 19 – Ватера, 20 – Сескло, 21 – Аликес, 22 – Либакос, 23 – Геракароу, 24 – Кримни-1, 25 – Волакс, 26 – Аполлония-1, 27 – Дафнеро, 28 – Болград.
Понтоцеросы имели гомонимно закрученные рога, и это хорошо отличает их от антилоп рода Gazellospira. От последних они отличаются и некоторыми особенностями строения пястных и плюсневых костей, в частности, формой проксимального сустава пястной кости и формой фасетки для os trapezoideocapitatum, которые у Gazellospira более округлены с медиальной стороны. По размерам P. ambiguus из Тавриды и Аполлонии-1 были мельче, чем Pontoceros surprine из Дманиси.
Авторы рода Pontoceros отнесли его к подсемейству Tragelaphinae Pilgrim, 1939 (Верещагин и др., 1971), и такое систематическое положение рода принято в ряде работ (Байгушева, 1971; Алексеева, 1977; Дмитриева, 1975, 1977; Титов, 2008). Н.К. Верещагин с соавт. (1971) полагали, что P. ambiguus был крупной антилопой, размером с гну и весом до 120–130 кг, и обликом походил на бонго Tragelaphus eurycerus Ogilby, 1837. Но по гомонимной скрученности рогов Pontocеros отличается от современных представителей трагеляфин; у последних рога закручены гетеронимно. По данным современных филогенетических и генетических анализов, эта группа, близкородственная Bovini, входит в качестве трибы в состав подсемейства Bovinae. Обе группы (Bovini и Tragelaphini) происходят, по-видимому, от миоценовых Boselaphini (Bibi et al., 2009). Некоторые особенности морфологии роговых стержней и зубов понтоцеросов указывают на их сходство с представителями трибы Caprini (Kostopoulos, 1997b, 2006; Vekua, 2012), которые в настоящее время включаются в подсемейство Antilopinae (Bibi et al., 2009). Среди современных видов семейства Bovidae только около 13% имеют гомонимно закрученные рога; они встречаются среди Caprini, Alcelaphini, а также некоторых Bovini, и почти не встречаются среди современных Antilopini, за исключением отдельных особей газелей (Kostopoulos, 2014). На основании строения неполной части черепа и гомонимного типа скручивания рогов у P. surprine А. Векуа отнес род Pontoceros к трибе Oiocerini Sokolov, 1953 подсемейства Caprinae (Vekua, 2012). Для уточнения систематической принадлежности рода Pontoceros нужны новые палеонтологические находки, детальное морфологическое сравнение с ископаемыми и современными видами и данные онтогенеза и филогенеза.
* * *
Таким образом, материалы по винторогим антилопам из пещеры Таврида позволяют уточнить стратиграфическое распространение, особенности эволюции и ареалы видов Gazellospira torticornis и Pontoceros ambiguus. Данные из Тавриды и анализ данных о биостратиграфическом распространении этих винторогих антилоп показывают, что ассоциация Gazellospira torticornis – Pontoceros ambiguus может считаться в Европе характерной для первой половины позднего виллафранка, зоны MNQ18, интервала ~1.8–1.5 млн л. н. Границы этого интервала определены на основе данных о последнем распространении G. torticornis и первом появлении Pоntoceros. Он соответствует первой половине позднего виллафранка Западной Европы (зона MNQ18), дунаю/эбуронию, стадиям MIS 65–57.
Находка G. torticornis в пещере Таврида показала, что в это время ареал вида охватывал и юг Восточной Европы, а не только европейское обрамление Средиземноморья и Малую Азию. Крым, вероятно, являлся одним из немногих поздневиллафранкских рефугиумов этого вида. Кроме того, материалы из Тавриды подтверждают тенденцию к уменьшению размеров у поздних G. torticornis. Присутствие G. torticornis на территориях, примыкающих к европейскому Средиземноморью, в Крыму и Турции в местонахождениях интервала ~2–1.5 млн л. н. (рис. 5, 6) говорит о существовании сплошного ареала этого вида в пределах Средиземноморской зоогеографической подобласти Палеарктики в начале позднего виллафранка. Антилопы Pontoceros ambiguus из местонахождения Таврида – одни из самых ранних представителей рода. Pontoceros появились на фоне усиливающейся глобальной тенденции к похолоданию и заметных палеобиогеографических изменений (см. Kahlke et al., 2011). Ареал рода Pontoceros приурочен к Восточно-Средиземноморской провинции и охватывал Грецию, юг Восточной Европы и Кавказ, а возможно, и Левант. В Греции фауны Либакос и Аликес близки по возрасту, и находки Pontoceros в Либакосе и Gazellospira в Аликесе могут говорить об одновременном существовании этих антилоп в позднем виллафранке и в этом регионе. Фауны Либакос и Аликес соотвествуют фаунистической стадии Фарнета в Италии и их возраст, по-видимому, близок к 1.5 млн л. н. (e.g., Koufos, Kostopoulos, 2016).
Рис. 6.
Хронологическое положение основных местонахождений винторогих антилоп Восточно-Cредиземноморской провинции.
Непосредственные предки винторогих антилоп родов Gazellospira и Pontoceros пока не установлены, хотя многие исследователи считали, что их следует искать в Азии. Дальнейшие исследования и новые находки позволят уточнить фаунистические связи поздневиллафранкских фаун Крыма и Кавказа, выявить возможные родственные связи антилоп этого региона с формами из Азии и Африки, и будут способствовать более точной реконструкции связей между фаунистическими событиями и миграций ранних гоминин в районе Восточного Средиземноморья.
Список литературы
Алексеева Л.И. Териофауна раннего антропогена Восточной Европы. М.: Наука, 1977. 216 с.
Байгушева В.С. Ископаемая териофауна Ливенцовского карьера (Северо-Восточное Приазовье) // Тр. Зоол. ин-та АН СССР. 1971. Т. 69. Вып. 49. С. 5–28.
Байгушева В.С. Новые данные о таманском фаунистическом комплексе из раскопа у села Семибалки (Приазовье) // Историко-археологические исследования в Азове и на Нижнем Дону в 1998 г. Вып. 16. Азов, 2000. С. 27–57.
Bерещагин Н.К., Алексеева Л.И., Давид А., Байгушева В.С. Род Pontoceros // Плейстоцен Тирасполя. Кишинев: Штиинца, 1971. С. 167–170.
Вислобокова И.А., Агаджанян А.К. Новые данные о крупных млекопитающих плейстоценовой фауны Трлица (Черногория, Центральные Балканы) // Палеонтол. журн. 2015. № 6. С. 86–103.
Дмитриева Е.Л. Ископаемые винторогие антилопы Западной Монголии и некотопых районов Средней Азии // Фауна и биостратиграфия мезозоя и кайнозоя Монголии. М.: Наука, 1975. С. 76–83.
Дмитриева Е.Л. Антилопы неогена Монголии и сопредельных территорий. М.: Наука, 1977. 120 с.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Докл. АН. 2019. Т. 485. № 3. С. 381–385.
Титов В.В. Крупные млекопитающие позднего плиоцена Северо-Восточного Приазовья. Ростов-на-Дону: Изд-во ЮНЦ РАН, 2008. 264 с.
Arribas A., Baeza E., Bermúdez D. et al. Nuevos registros paleontológicos de grandes mamíferos en la Cuenca de Guadix-Baza (Granada): aportaciones del Proyecto Fonelas al conocimiento sobre las faunas continentales del Plio-Pleistoceno europeo // Bol. Geol. Minerol. 2004. V. 115. № 3. P. 567–581.
Athanassiou A. Contribution to the study of the fossil mammals of Thessaly (Greece). Tesis Doctoral. Atenas: Univ. Atenas, 1996. 353 p.
Athanassiou A. Gazellospira torticornis (Aymard, 1854) from the late Pliocene locality of Sésklo (Thessaly, Greece) // Quaternaire. 2005. V. 2. P. 137–44.
Azanza B., Morales J. Los artiodáctilos de Huélago, Huescar-1 y Cúllar de Baza-1 (cuenca de Guadix-Baza, Granada) // Trabajos sobre Neógeno-Cuaternario.1989. V. 11. P. 289–314.
Bellucci L., Sardella R. The last Antilopini bovids from the Early Pleistocene of Italy // Quatern. Intern. 2015. V. 357. P. 245–252.
Bibi F., Bukhsianidze M., Gentry A.W. et al. The fossil record and evolution of Bovidae: State of the field // Palaeontol. Electron. 2009. V. 12. Iss. 3. 10A. 10 p. http://palaeo-electronica.org/2009-3/169/index.html
Bouvrain G. Le gisement de vertébrés pliocènes de Çalta, Ankara, Turquie. Bovidés // Geodiversitas. 1998. V. 20. P. 467–485.
Bukhsianidze M. The fossil Bovidae of Dmanisi. Doct. Thesis. Ferrara: Univ. Ferrara, 2005. 192 p.
Bukhsianidze M., Chagelishvili R., Hertler C. et al. New findings about the Akhalkalaki site (Southern Georgia, Caucasus; 0.98–0.78 Ma) // Abstr. Book VIth Intern. Conf. on Mammoths and their Relatives. Thessaloniki: Aristotle Univ., 2014. P. 38 (Sci. Ann. School Geol. V. 102).
Bukhsianidze M., Koiava K. Synopsis of the terrestrial vertebrate faunas from the Middle Kura Basin (Eastern Georgia and Western Azerbaijan, South Caucasus) // Acta Palaeontol. Pol. 2018. V. 63. № 3. P. 441–461.
Crégut-Bonnoure E. Apport des Caprinae et Antilopinae (Mammalia, Bovidae) à la biostratigraphie du Pliocène terminal et du Pléistocène d’Europe // Quaternaire. 2007. V. 18. № 1. P. 73–97.
Demirel F.A., Mayda S. A new Early Pleistocene mammalian fauna from Burdur Basin, SW Turkey // Russian J. Theriol. 2014. V. 13. № 2. P. 55–63.
Dorlhac J. Notice géologique sur le cratère du Coupet, et sur son gisement de gemmes et d’ossements fossils // Ann. Soc. Agricult. Sci. Art. Commerc. Puy. 1854. V. 19. P. 597–617.
Duvernois M.P., Guérin C. Les Bovidae (Mammalia) du Villafranchien supérieur d’Europe occidentale // Géobios. 1989. V. 22. P. 339–379.
Garrido G. The most recent and complete set of Gazellospira torticornis (Bovidae, Artiodactyla, Mammalia) fossils known for the late Upper Pliocene of Western Europe (Fonelas P-1, Gaudix Basin, Granada) // Vertebrados del Plioceno superior terminal en el suroeste de Europa: Fonelas P-1 y el Proyecto Fonelas / Ed. Arribas A. Madrid: Inst. Geol. Minero España, 2008. P. 413–460 (Cuadern. Museo Geominero. № 10).
Heintz E. La presence de Gazellospira torticornus dans le gisement Villafranchien de Saint-Vallier (Dome). Essai de repartition géographique et stratigraphique // Bull. Soc. Géol. France. 1966. V. 7. № 8. P. 25–30.
Heintz E. Les Cervides Villafranchiens de France et d’Espagne // Mém. Mus. Nat. Hist. Natur. Ser. C. 1970. T. 22. P. 1–202.
Kahlke R.D., García N., Kostopoulos D.S. et al. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe // Quatern. Sci. Rev. 2011. V. 30. P. 1368–1395.
Konidaris G.K., Tourloukis V., Kostopoulos D.S. et al. Two new vertebrate localities from the Early Pleistocene of Mygdonia Basin (Macedonia, Greece): Preliminary results // C. R. Palevol. 2015. V. 14. P. 353–362.
Kostopoulos D.S. The Plio-Pleistocene artiodactyls from Macedonia, Greece: 2. The fossillirerous locality of Volaks (Volakas Basin, Drama, NE Greece) // Paleontol. Evol. 1997a. V. 30–31. P. 83–92.
Kostopoulos D.S. The Plio-Pleistocene artiodactyls (Vertebrata, Mammalia) of Macedonia 1. The fossiliferous site “Apollonia-1”, Mygdonia basin of Greece // Geodiversitas. 1997b. V. 19. № 4. P. 845–875.
Kostopoulos D.S. Greek bovids through time // Ann. Géol. Pays Hellén. 2006. V. 41. № 1. P. 141–152.
Kostopoulos D.S. Taxonomic re-assessment and phylogenetic relationships of Miocene homonymously spiral-horned antelopes // Acta Palaeontol. Pol. 2014. V. 59. № 1. P. 9–29.
Kostopoulos D.S., Koufos G.D. The Plio-Pleistocene artiodactyls of Macedonia (Northern Greece) and their biostratigraphic significance, preliminary report // C. R. Acad. Sci. Sér. 2. 1994. V. 318. P. 1267–1272.
Kostopoulos D.S., Sen S. Late Pliocene (Villafranchian) mammals from Sirakol-Tepe, Ankara, Turkey // Mitt. Bayer. Staatsslg. Paläontol. Hist. Geol. 1999. Bd 39. P. 165–202.
Kostopoulos D.S., Vassiliadou K., Koufos G.D. The beginning of the Pleistocene in the Balkan area according to the Mammal record: palaeozoogeographical approach // Ann. Géol. Pays Hellén. 2002. V. 39. P. 253–278.
Koufos G. The Villafranchian mammalian faunas and biochronology of Greece // Boll. Soc. Paleontol. Ital. 2001. V. 40. № 2. P. 217–223.
Koufos G.D., Kostopoulos D.S. The Pliocene large mammal record in Greece: Implication for early human dispersal into Europe // Palaeoanthropology of the Balkans and Anatolia: Human evolution and its context / Eds. Harvati K., Roksandis M. Dortrecht: Springer, 2016. P. 269–280.
Krijgsman W., Tesakov A., Yanina T. et al. Quaternary time scales for the Pontocaspian domain: Interbasinal connectivity and faunal evolution // Earth-Sci. Rev. 2019. V. 188. P. 1–40.
Lordkipanidze D., Jashashvili T., Vekua F. et al. A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo // Science. 2013. V. 342. P. 326–330.
Lyras G.A., van der Geer A.A.E. The Late Pliocene vertebrate fauna of Vatera (Lesvos Island, Greece) // Cranium. 2007. V. 24. № 2. P. 11–24.
Martínez-Navarro B., Belmaker M., Bar-Yosef O. The bovid assemblage (Bovidae, Mammalia) from the Early Pleistocene site of Ubeidiya, Israel: Biochronological and paleoecological implications for the fossil and lithic bearing strata // Quatern. Intern. 2012. V. 267. P. 78–97.
Nomade S., Pastre J.F., Guillou H. et al. 40Ar/39Ar constraints on some French landmark Late Pliocene to Early Pleistocene large mammalian paleofaunas: Paleoenvironmental and paleoecological implications // Quatern. Geochron. 2014. V. 21. P. 2–15.
Pilgrim G.E., Schaub S. Die schraubenhörnige Antilope des europäischen Oberpliocaens und ihre systematische Stellung // Abh. Schweiz. Paläontol. Ges. 1939. Bd 62. S. 1–30.
Radulesko C. Artiodactyles du Pliocène et du Pléistocène inférieur de Roumanie // Quaternaire. 2005. V. 2. P. 191–200.
Rădulescu C., Samson P., Petculescu A., Ştiucă E. Pliocene large mammals of Romania // Coloq. Paleontol. 2003. V. 53. № 1. P. 549–558.
Rădulescu C., Samson P., Ştiucă E. Cadre biostratigraphique du paléolithique inferieur en Roumanie // Quaternaire. 1998. V. 9. № 4. P. 283–290.
Rodrigo M.A. Los bóvidos Villafranquiensis de la Puebla de Valverde y Villarroya: systemática, filogenia y paleobiología. Tesis Doct. Zaragoza: Univ. Zaragoza, 2011. 433 p.
Sickenberg O. Etude du “Villafranchien” en Anatolie (Turquie) // Bull. Bur. Rech. Géol. Min. 1974. V. 78. № 1. P. 241–245.
Spassov N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe // Rev. Paléobiol. 2003. V. 22. P. 197–229.
Spassov N. Southeastern Europe as a route for the earliest dispersal of Homo towards Europe: Ecological conditions and the timing of the first human occupation of Europe // Paleoanthropology of the Balkans and Anatolia: Human evolution and its context / Eds. Harvati K., Roksandic M. Dordrecht: Springer, 2016. P. 281–290.
Spassov N., Crégut-Bonnoure E. Premières données sur les Bovidae Villafranchiens de Bulgarie // C. R. Acad. Sci. 1999. V. 328. P. 493–498.
Steensma K.-J. Plio-Pleistozane Grosssaugetiere (Mammalia) aus dem Becken von Kastorial Grevena, sudlich von Neapolis – NW-Griechenland. Doct. Thesis. Clausthal: Tech. Univ., 1988. 315 p.
Tesakov A.S., Dodonov A.E., Titov V.V., Trubikhin V.M. Plio-Pleistocene geological record and small mammal faunas, eastern shore of the Azov Sea, Southern European Russia // Quatern. Intern. 2007. V. 160. P. 57–69.
Vekua A. New spiral-horned antelope in Dmanisi fauna // Bull. Georg. Nat. Acad. Sci. 2012. V. 6. № 3. P. 139–144.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал