

Палеонтологический журнал, 2020, № 2, стр. 78-86
Новый морфотип зубов рыб отряда Eurynotoidiformes (Actinopterygii) из верхнепермских отложений европейской России
А. С. Бакаев *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: alexandr.bakaev.1992@mail.ru
Поступила в редакцию 09.01.2019
После доработки 01.02.2019
Принята к публикации 16.08.2019
Аннотация
В терминально-северодвинских отложениях Восточной Европы (местонахождение Марьюшкина Слуда-С, Вологодская обл.) обнаружен новый морфотип поликуспидных зубов лучеперых рыб отряда Eurynotoidiformes, предположительно ассоциируемый с видом Isadia opokiensis A. Minich et Andrushkevich, 2017, описанным по изолированным чешуям из стратиграфически и географически близкого местонахождения Мутовино. Зубы морфотипа I. opokiensis робустные, характеризуются сочетанием мощного контрфорса и широкой коронки с большим (до восьми) количеством зубцов лопатовидной формы, несущих высокие акродиновые колпачки и образующих слабоизогнутую рабочую поверхность. Вершинки зубов имеют хорошо выраженные площадки прижизненного стирания на лабиальной стороне, что указывает на преобладание соскребающего типа питания водорослями-обрастателями. Конструкция челюстных зубов I. opokiensis является морфологически переходной между зубами I. suchonensis и I. arefievi, что позволяет объединить все три морфотипа в единую эволюционную линию. Предполагается, что сходная (расширенная) форма зубцов на челюстных зубах другого вида Isadia – I. aristoviensis – возникла конвергентно вследствие модификации более примитивного морфотипа, характерного для уржумских эвринотоидиформ.
ВВЕДЕНИЕ
Древнейшие лучеперые рыбы, освоившие облигатную альгофагию, относятся к отряду Eurynotoidiformes, широко распространенному в средней-поздней перми на территории Восточной Европы (Миних, Миних, 1990; Esin, 1997). Группа является эндемиком данного региона. Она динамично эволюционировала до конца палеозоя и вымерла на пермо–триасовом рубеже. На растительноядность большинства эвринотоидиформных рыб указывает специализированное (поликуспидное) строение их челюстных зубов, которое находит множество аналогий среди современных рыб-фитофагов из семейств Characidae, Cichlidae, Acanthuridae, Siganidae, Scatophagidae и Mugilidae (Bellwood, 2003; Bellwood et al., 2014).
Для наиболее ранних – уфимско-казанских – представителей отряда [Eurynotoides nanus (Eichwald, 1861), E. costatus (Eichwald, 1861), Adzvalepis puchontoi Yankevich, 1998], описанных по единичным неполным находкам, строение зубной системы остается неизвестным. Формы с поликуспидными зубами начинают фиксироваться в верхнеуржумских отложениях, причем по ряду признаков (в том числе особенностям строения зубной системы) могут быть отнесены к различным видам и родам (Миних, Миних, 1990). Полноскелетные находки эвринотоидиформ очень немногочисленны и не дают полного представления о многих структурах челюстного аппарата, строение которого как комплексного образования для группы в целом остается слабо изученным. При этом, сложная морфология зубов уржумско-вятских эвринотоидиформ видоспецифична и позволяет прослеживать эволюционные тренды внутри группы, а, при подборе современных аналогов, реконструировать трофическую специализацию конкретных представителей отряда. Для решения этих задач может быть использован, в том числе, и изолированный материал.
МАТЕРИАЛ
При изучении комплекса ихтиолитов, полученных с помощью промывки породы из местонахождения Марьюшкина Слуда-С (Вологодская обл., бассейн р. Сухоны; рис. 1), обнаружено 9 зубов эвринотоидид ранее неизвестного морфотипа (экз. ПИН, №№ 5652/200–209; сборы В.В. Буланова). Местонахождение приурочено к верхней половине кичугской пачки полдарсской свиты северодвинского яруса (Голубев, 2000, рис. 31 ; Arefiev et al., 2015, рис. 12 ). Костеносная порода отобрана из основания аллювиальной линзы, где костям рыб сопутствуют изолированные остатки тетрапод. Ранее М.П. Арефьев и др. (Arefiev et al., 2015) определили из этого местонахождения виды рыб Sludalepis spinose и Plotnikovichthys gorodokensis. В новых сборах обнаружены остатки Toyemia tverdochlebovi, T. cf. blumentalis, Strelnia certa, Mutovinia stella, Geryonichthys longus, Isadia aristoviensis, I. opokiensis, Acropholis sp., Boreolepis tataricus, Varialepis vitalii (определения автора). Сопутствующие таксоны тетрапод могут быть отнесены к ильинскому субкомплексу соколковского фаунистического комплекса и включают представителей Kotlassiidae (Microphon arcanus; определение В.В. Буланова), Dvinosauridae (Dvinosaurus sp.), Pareiasauria gen. indet., Chroniosuchidae (Chroniosaurus levis), Gorgonopia fam. indet. и Procynosuchidae (Sludica bulanovi) (Ивахненко и др., 1997; Ивахненко, 2012).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Высокая степень специализации поликуспидных зубов позволяет отнести их к роду Isadia (отряд Eurynotoidiformes). При этом, морфологических данных, дающих возможность уточнить их систематическое положение до видового уровня, недостаточно, поскольку известные ныне виды этого рода основаны на морфологически неполном или несравнимом материале (Миних, 1986, Миних, Миних, 1990; Миних и др., 2015; Миних, Андрушкевич, 2017). Однако морфо-функциональный анализ материала и его сравнение с имеющимися данными по другим видам позволяет выделить новый морфотип зубных элементов, относящийся к этому роду, и выдвинуть предположение о его систематической принадлежности.
Верхи северодвинского яруса хорошо охарактеризованы ихтиофауной, в том числе остатками трех эвринотоидиформ, относимых к роду Isadia: I. suchonensis A. Minich, 1986, I. aristoviensis A. Minich, 1990 и I. opokiensis A. Minich et Andrushkevich, 2017.
Морфология зубов двух первых видов описана и обсуждалась ранее (Миних, Миних, 1990; Esin, 1997; Буланов, Миних, 2017; Pindakiewicz et al., in press).
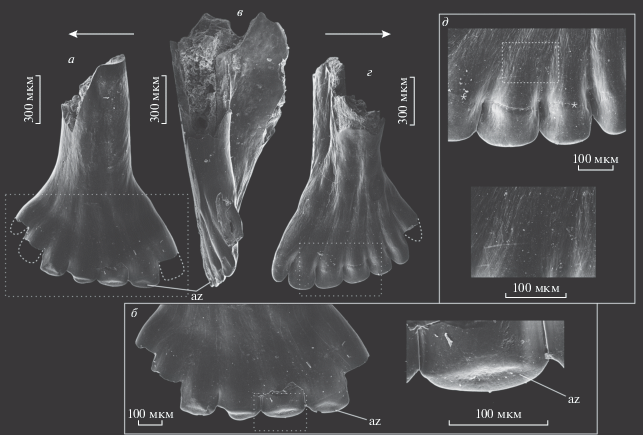
Зубы нового морфотипа робустные, их основание формирует с лингвальной стороны мощный широкий контрфорс, захватывающий перибазальную часть коронки и здесь распадающийся на пологие валики, выходящие к апикально расположенным зубцам (рис. 2). Коронка широкая, примерно вдвое шире основания, полого наклонена внутрь ротовой полости (угол наклона 10°–20°). Рабочий край коронки образован шестью–семью зубцами лопатовидной формы с крупными (во всю высоту зубца), уплощенными, округленными по краям акродиновыми колпачками. Краевые зубцы коронки более узкие, но также с округлыми уплощенными окончаниями. Зубцы расположены очень плотно, боковые края оснований зубцов почти смыкаются. Режущие канты по бокам зубцов не развиты. Профиль образуемого зубцами рабочего края коронки дугообразный, с выраженным скосом рострально. С лабиальной стороны на всех зубцах (кроме одного или двух наиболее дистальных) имеются хорошо выраженные площадки прижизненного стирания.
Рис. 2.
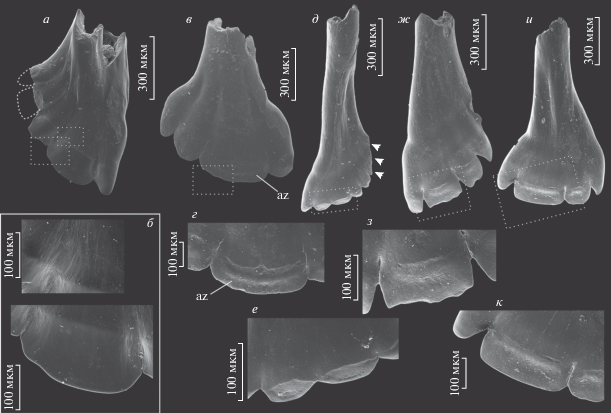
Зубы Isadia opokiensis Minich et Andrushkevich, 2017: а, б – экз. ПИН, № 5652/207, зуб верхней челюсти: а – с лабиальной стороны, б – увеличенные участки коронки; в–д – зубы верхней челюсти: в – экз. ПИН, № 5652/208 с лингвальной стороны; г – экз. ПИН, № 5652/209 с лингвальной стороны, д – тот же экземпляр, увеличенная часть коронки; Вологодская обл., Великоустюгский р-н, местонахождение Марьюшкина Слуда-С; северодвинский ярус, путятинский горизонт, полдарсская свита, кичугская пачка. Стрелки указывают в ростральном направлении; звездочки маркируют границу акродинового колпачка и ганоина. Обозначение: az – площадки истирания.
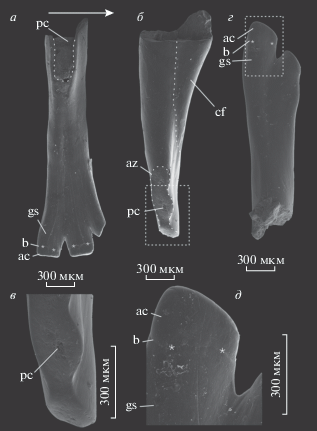
Зубы I. aristoviensis, благодаря характерной форме, легко диагностируются и известны из многих местонахождений верхнесеверодвинско–терминальновятского интервала (Миних и др., 2015; Pindakiewicz et al., in press). В большом количестве они обнаружены и в местонахождении Марьюшкина Слуда-С. В отличие от зубов нового морфотипа, контрфорс на зубах I. aristoviensis (рис. 3) развит слабее (угол между лингвальным краем и длинной осью зуба составляет около 15°–20°), а значительно более узкие коронки (апикальная часть не более, чем в 1.5 раза шире шейки) несут меньшее количество зубцов. Верхнечелюстные зубы у данного вида, предположительно, имеют три–четыре зубца, в то время как у зубов нижней челюсти коронки бикуспидные (Pindakiewicz et al., in press). Форма зубцов варьирует от прямоугольной (при стирании акродиновых колпачков) до трапециевидной со скругленными краями (при не выраженном износе), однако зубцы отчетливо разделены и не смыкаются боковыми сторонами, сохраняя у оснований небольшой, но хорошо выраженный режущий кант. Акродиновый колпачок составляет не более половины высоты зубца. Рабочая кромка коронки, формируемая совокупностью расположенных на ней зубцов, умеренно наклонена лингвально (примерно 15°–25°) и круто изогнута, вследствие значительно более слабого развития и низкого положения самого переднего (рострального) зубца. Часто зубы I. aristoviensis несут очевидные следы прижизненного стирания в виде плоских площадок на концах зубцов, ориентированных под прямым углом к продольной оси зубца. В ряде случаев наблюдается стирание рабочего края коронки до самых оснований зубцов и формирование единой широкой скребущей поверхности (рис. 3, б, в; az).
Рис. 3.
Зубы Isadia aristovensis A. Minich, 1990: а–в – зубы верхней челюсти: а – экз. ПИН, № 5652/800 с лингвальной стороны; б, в – экз. ПИН, № 5652/801: б – вид спереди, в – увеличенная часть коронки (видно отверстие пульпарной полости и расположенные вокруг дентиновые канальцы); г – экз. ПИН, № 5652/802, зуб нижней челюсти с лабиальной стороны, д – детали строения зубцов того же экземпляра при большем увеличении; Архангельская обл., Карпогорский р-н, местонахождение Раша; вятский ярус, верхневятский подъярус. Стрелки указывают в ростральном направлении; звездочками отмечена граница ганоинового слоя и акродинового колпачка. Обозначения: ac – акродиновый колпачок; az – площадки истирания; b – наружная граница акродинового колпачка и ганоина; cf – контрфорс; gs – ганоиновое поле; pc – пульпарная полость.
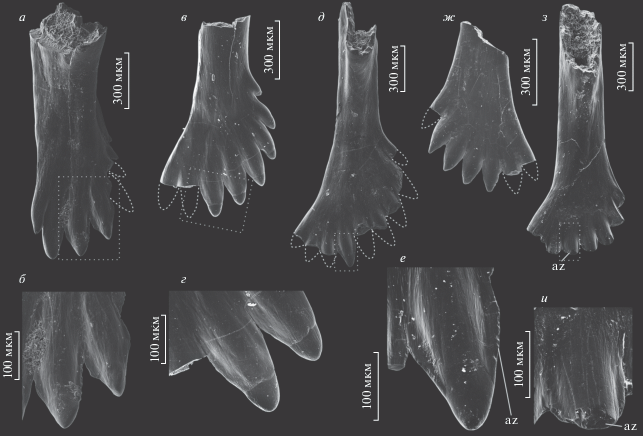
Зубы I. suchonensis близки к новому морфотипу расширенной формой коронки, которая в 2.5 раза шире основания, и большим количеством (до девяти) расположенных на ней зубцов (рис. 4). Сами зубцы, однако, сильно отличаются удлиненно-конической (не уплощенной) формой и наличием хорошо развитых режущих кантов, начинающихся от оснований акродиновых колпачков и расширяющихся в базальном направлении. Промежутки между основаниями зубцов сужены до узких щелей. Акродиновый колпачок образует не более трети высоты зубца и имеет округло-коническое окончание. Вследствие такой формы зубцов, рабочая поверхность коронок челюстных зубов I. suchonensis сильно рассечена. Кроме того, коронка в боковой проекции имеет резко асимметричный профиль из-за более базального положения зубцов, расположенных на ее ростральном крае. Таким образом, рабочий край формирует веерообразно изогнутый профиль с резким понижением рострально. Контрфорс развит слабо и едва достигает основания расширения коронки (угол между лингвальным и лабиальным краями основания составляет около 10°–15°), а толщина стенки зуба тоньше, чем у I. aristoviensis и описанных выше зубов нового морфотипа.
Рис. 4.
Зубы Lapkosubia sp. и Isadia suchonensis A. Minich, 1986: а – Lapkosubia sp., экз. ПИН, № 5157/596, зуб верхней челюсти с лингвальной стороны; б – тот же экземпляр, увеличенная часть коронки; Республика Татарстан, Камско-Устьинский р-н, Монастырский овраг-D; уржумский ярус, третья свита; в–и – Isadia suchonensis: в – экз. ПИН, № 5652/600, зуб верхней челюсти с лабиальной стороны, г – тот же экземпляр, детали строения зубцов при большем увеличении; Нижний Новгород, местонахождение Лагерный Овраг-3; вятский ярус, верхневятский подъярус, жуковский горизонт, обнорская свита; д – экз. ПИН, № 5652/803, зуб верхней челюсти с лингвальной стороны, е – тот же экземпляр, увеличенная часть коронки; ж – экз. ПИН, № 5652/804, зуб верхней челюсти с лабиальной стороны; з – экз. ПИН, № 5652/805, зуб верхней челюсти с лингвальной стороны, и – тот же экземпляр, увеличенная часть коронки; Архангельская обл., Карпогорский р-н, местонахождение Раша; вятский ярус, верхневятский подъярус. Обозначение: az – площадки истирания.
Исходя из перечисленных отличий, новый морфотип зубов отличается от морфотипов как I. aristoviensis, так и I. suchonensis. Наиболее близки к новому морфотипу по строению зубы I. arefievi. Данный вид был недавно описан А.В. Миних и др. (2015) из верхов верхневятского подъяруса бассейна р. Малая Северная Двина (местонахождение Элеонора, Архангельская обл.) по изолированным элементам. Зубы этого вида (рис. 6), как и у нового морфотипа, имеют мощный высокий контрфорс, достигающий середины коронарного расширения зуба. Коронки значительно (в 2.5 раза) шире оснований и имеют изогнутый профиль рабочей поверхности, форма которого вариабельна и, предположительно, зависела от положения зуба в зубном ряду. Прямоугольная плоская форма самих зубцов (часто со скругленными краями) также сходна с таковыми у нового морфотипа, однако один из зубцов, как правило, значительно шире остальных. Краевые зубцы коронки расширены слабо и аналогичны зубцам в том же положении на зубах нового морфотипа.
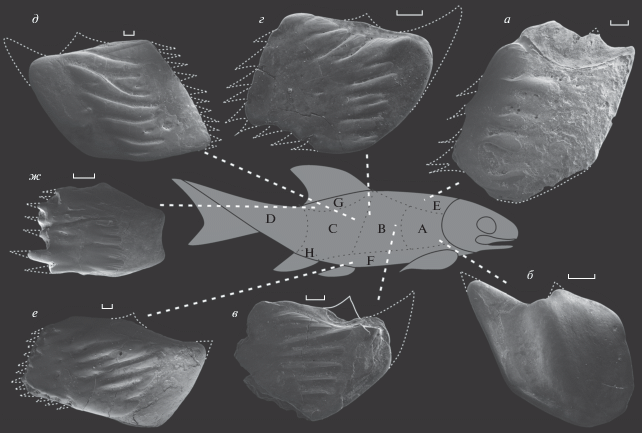
Строение зубов Isadia opokiensis точно не известно, потому что данный таксон описан по изолированным чешуям (Миних, Андрушкевич, 2017), но зубы нового морфотипа могут быть ассоциированы именно с этим видом, и далее будут называться “морфотип Isadia opokiensis”, поскольку найдены в местонахождении Марьюшкина Слуда-С совместно с относящимися к нему чешуями (экз. ПИН, № 5652/210-219; рис. 5). Для этих чешуй характерны увеличенная толщина, мощный киль на внутренней стороне, а также широкие и выпуклые ганоиновые гребни свободного поля, ориентированные диагонально, от антеро-дорсального края чешуи к ее каудо-вентральному углу. Передние окончания гребней орнаментированы тонкой струйчатостью. Эти чешуи отличаются от чешуй I. aristoviensis и I. suchonensis формой и ориентацией гребней свободного поля.
Рис. 5.
Чешуи c различных участков тела Isadia opokiensis Minich et Andrushkevich, 2017 (буквами на силуэте отмечены области распространения различных морфотипов чешуй; по: Есин, 1990): а – экз. ПИН, № 5652/200, чешуя из зоны Е; б – экз. ПИН, № 5652/201, чешуя из зоны А; в – экз. ПИН, № 5652/202, чешуя участка B, вид с внутренней стороны; г – экз. ПИН, № 5652/203, чешуя участка С; д – экз. ПИН, № 5652/204, чешуя участка С; е – экз. ПИН, № 5652/205, чешуя из зоны F; ж – экз. ПИН, № 5652/206, чешуя участка D; Вологодская обл., Великоустюгский р-н, местонахождение Марьюшкина Слуда-С; северодвинский ярус, путятинский горизонт, полдарсская свита, кичугская пачка. Все изображены с наружной стороны (если нет других указаний). Зубцы на заднем и нижнем крае всех чешуй обломаны.
Типовое для I. opokiensis местонахождение Мутовино расположено в 5 км от местонахождения Марьюшкина Слуда-С (Голубев, 2000; Arefiev et al., 2015). Оба разреза хорошо коррелируются в пределах общего комплекса верхнепермских отложений, вскрытого в бассейне р. Сухоны, приурочены к верхней части кичугской пачки, и содержат сходный комплекс рыб и тетрапод.
Рис. 6.
Зубы Isadia arefievi Minich, 2015: а – экз. ПИН, № 5652/806, зуб верхней челюсти с лингвальной стороны, б – тот же экземпляр, увеличенная часть коронки; в – экз. ПИН, № 5157/807, зуб верхней челюсти с лабиальной стороны, г – тот же экземпляр, увеличенная часть коронки; Архангельская обл., Карпогорский р-н, местонахождение Раша; вятский ярус, верхневятский подъярус; д – экз. ПИН, № 5652/601, зуб верхней челюсти с лабиальной стороны, е – тот же экземпляр, увеличенная часть коронки; ж – экз. ПИН, № 5652/602, зуб верхней челюсти с лабиальной стороны, з – тот же экземпляр, увеличенная часть коронки; и – экз. ПИН, № 5652/603, зуб верхней челюсти с лабиальной стороны, к – тот же экземпляр, увеличенная часть коронки; Нижний Новгород, местонахождение Лагерный Овраг-3; вятский ярус, верхневятский подъярус, жуковский горизонт, обнорская свита. Стрелки указывают в ростральном направлении. Обозначение: az – площадки истирания.
Зубы иного типа (помимо обсуждаемых выше), которые могли бы быть соотнесены с найденными в местонахождении Марьюшкина Слуда-С чешуями I. opokiensis, отсутствуют.
ОБСУЖДЕНИЕ
Морфо-функциональный анализ зубов различных эвринотоидиформов указывает на различные способы питания относящихся к этому отряду рыб, их интерпретации приведены ранее М. Пиндакевичем и др. (Pindakiewicz et al., in press). Характерной особенностью зубов морфотипа Isadia opokiensis является сочетание широкой, рассеченной на многочисленные широкие и уплощенные зубцы коронки и мощного контрфорса, формируемого основанием. Данные признаки организации зуба в совокупности с наличием площадок стирания на лабиальной стороне зубцов указывают на соскребающий тип питания водной растительностью, по аналогии с данными о строении и функции зубов подобного типа у некоторых современных Cichlidae (Yamaoka, 1983; Purnell et al., 2012; Dieleman et al., 2015).
От I. opokiensis зубы I. arefievi отличаются меньшим (не более пяти) количеством зубцов коронки и наличием сильно расширенного зубца в ее осевой зоне. Примечательно, что, в отличие от I. opokiensis, промежутки, разделяющие зубцы, часто не прорезают рабочую часть коронки глубже основания акродиновых колпачков (рис. 6, а, б), которые, таким образом, сливаются базальными отделами и до конца не разделены. Предположительно это может свидетельствовать об образовании единого увеличенного зубца путем слияния нескольких отдельных, имевшихся у предковой формы. Если данное предположение верно, то наиболее вероятным исходным таксоном для I. arefievi является I. opokiensis, зубы которого близки к данному морфотипу по большинству рассмотренных параметров (ширина и профили рабочего края коронки, высокие мощные акродиновые колпачки, уплощенная форма зубцов и их тесное прилегание боковыми сторонами, форма краевых зубцов коронки, отсутствие режущих окантовок, наличие мощного контрфорса, толщина стенок оснований зубов).
Необходимость слияния зубцов коронки в более широкую рабочую структуру у I. arefievi диктовалась дальнейшей адаптацией зубной системы к соскребающему типу питания водорослевым перифитоном, на которую указывает наличие у обоих видов развитых площадок стирания на лингвальной стороне коронок.
Находки зубов I. opokiensis и I. aristoviensis в местонахождении Марьюшкина Слуда-С свидетельствуют о том, что специализированные биоморфы с соскребающим типом питания среди Eurynotoidiformes появились не позднее конца северодвинского века (учитывая глубину специализации зубов I. opokiensis, даже раньше этой временной отметки). Зубы I. aristoviensis значительно менее специализированы: контрфорс развит слабее, а коронки значительно уже, и несут не более четырех плоских прямоугольных зубцов, сохраняющих в местах слияния режущий кант (рис. 3). При этом наблюдаемые площадки стирания ориентированы перпендикулярно продольной оси зубцов (рис. 3, б, в), т.е., не так, как у I. opokiensis, что в совокупности указывает на значительные отличия в способе и стратегии питания обоих видов. Меньшая специализация зубной системы I. aristoviensis в направлении соскребающего типа питания позволяет рассматривать этот морфотип в качестве возможного предшественника морфотипа I. opokiensis.
Принципиальное сходство с морфотипом I. opokiensis в организации челюстных зубов обнаруживается у I. suchonensis: коронки сильно расширены и несут значительно большее (до девяти) число зубцов. Их коническая форма, наличие хорошо развитых режущих окантовок, отсутствие следов активного стирания и слабо развитый контрфорс свидетельствуют о преобладании движений челюстей стригущего типа и вероятности питания пучками нитчатых водорослей.
Зубы поликуспидного типа формируются в эволюции эвринотоидиформных рыб очень рано (не позднее позднеуржумского времени). Коническая форма зубцов характерна для наименее специализированных представителей отряда, для которых многовершинность доказана (Kichkassia furkae A. Minich, 1986). Несмотря на отличия в строении зубцов и контрфорса, в целом строение зубов морфотипа I. suchonensis более соответствует предковому состоянию для морфотипа I. opokiensis; наблюдаемые различия являются коррелятивно связанными и синхронно изменялись по мере переориентации на иной тип питания, при котором стало преобладать соскребание растительной массы с твердых субстратов. Данное предположение подтверждается субконической формой краевых (наименее функциональных и слабее модифицированных) зубцов коронок морфотипов I. opokiensis и I. arefievi, характерных для исходного морфотипа (I. suchonensis).
Происхождение морфотипа I. aristoviensis остается наименее очевидным. Формирование уплощенных зубцов коронок у этого вида, вероятно, происходило конвергентно и было связано со сходной трофической переориентацией – переходом к соскабливающему типу питания, т.е., проходило независимо от линии I. suchonensis–I. opokiensis–I. arefievi (рис. 7). Эволюционная последовательность “конический зуб”–Kichkassia–Lapkosubia–Isadia suchonensis–I. opokiensis–I. arefievi представляется наиболее вероятной, так как количество морфологических отличий в строении зубов у соседних видов данного ряда минимально, и составляет два (в первых двух случаях) или даже один (в третьем случае) признак.
Рис. 7.
Эволюционный ряд некоторых морфотипов поликуспидных зубов Eurynotoidiformes (от гипотетического неспециализированного предка): а – зуб неспециализированной лучеперой рыбы; б – Kichkassia furkae; в – Lapkosubia sp.; г – Isadia suchonensis; д – Isadia opokiensis; е – Isadia arefievi; ж – Isadia aristoviensis. Все зубы изображены с лабиальной стороны, контрфорс показан пунктиром в проекции. Серым отмечены рабочие поверхности. Прерывистыми стрелками показаны менее достоверные переходы (объяснения в тексте). Обозначения: b – наружная граница акродинового колпачка и ганоина (маркирована пунктиром); cf – контрфорс.
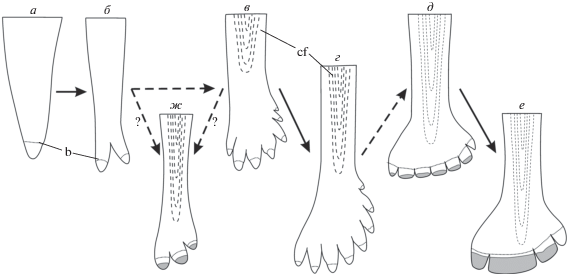
Аналогичный процесс модификации зубов у предков I. aristoviensis не привел к столь же значительному преобразованию зубной системы. Тем не менее, с момента своего появления в позднесеверодвинское время, морфотип I. aristoviensis просуществовал до самого конца перми и представлял собой одну из самых устойчивых и широко распространенных биоморф восточно-европейских бассейнов конца палеозоя. Наиболее вероятно, что предковый таксон для I. aristoviensis по строению зубной системы был близок к видам рода Lapkosubia или даже Kichkassia, коронки которых несут сходное количество зубцов (Миних, Миних, 1990); среди этих же форм следует искать предковый вид для эволюционной последовательности Isadia suchonensis–I. opokiensis–I. arefievi.
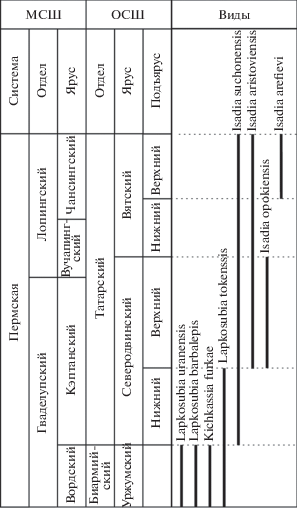
Рассмотренная на примере строения челюстных зубов эволюционная последовательность морфотипов, относимых к роду Isadia, хорошо согласуется со стратиграфическими данными (рис. 8). Остатки I. opokiensis определены только из верхов северодвинского яруса (Мутовино, Марьюшкина Слуда-С), в то время как I. arefievi характеризует самые верхи вятского яруса (местонахождения Элеонора, Раша, Жуков Овраг-1А, Лагерный овраг-3, Соковка, Федурники). Совместное появление I. opokiensis и I. aristoviensis в региональном разрезе с учетом разделяющей их значительной морфологической дистанции свидетельствуют против их прямого филогенетического родства; вместе с тем, исходные для обоих видов морфотипы зубов (роды Kichkassia, Lapkosubia) появляются уже в верхах уржумского яруса.
Список литературы
Буланов В.В., Миних А.В. Конвергентный морфогенез зубной системы тетрапод и лучеперых рыб средней–поздней перми Восточной Европы // Матер. конф. “Морфогенез в индивидуальном и историческом развитии: онтогенез и формирование биологического разнообразия”. М.: ПИН РАН, 2017. С. 9–11.
Голубев В.К. Пермские и триасовые хрониозухии и биостратиграфия верхнетатарских отложений Восточной Европы по тетраподам. М.: Наука, 2000. С. 1–174. (Тр. ПИН РАН. Т. 276.)
Есин Д.Н. Чешуйный покров Amblypterina costata (Eichwald) и таксономия палеонисцид по изолированным чешуям // Палеонтол. журн. 1990. № 2. С. 89–96.
Ивахненко М.Ф. Пермские Cynodontia (Theromorpha) Восточной Европы // Палеонтол. журн. 2012. № 2. С. 89–97.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Миних А.В. Новый род палеонисков из поздней перми Южного Приуралья. Саратов: Изд-во СарГУ, 1986. 13 с. (Деп. в ВИНИТИ 18.04.1986, № 2837- В).
Миних А.В., Андрушкевич С.О. Новый вид лучеперой рыбы из верхней перми Московской синеклизы // Изв. Саратовск. ун-та. Нов. сер. 2017. Т. 17. Сер. Науки о Земле. Вып. 2. С. 100–105.
Миних А.В., Арефьев М.П., Голубев В.К. Новый вид рыбы рода Isadia (Actinopterygii, Eurynotoidiformes) из нового местонахождения на р. Малая Северная Двина (терминальная пермь, Вологодская область) // Палеонтол. журн. 2015. № 6. С. 54–63.
Миних А.В., Миних М.Г. Ревизия некоторых палеонисков и новые рыбы из верхней перми Восточно–Европейской платформы и возможности использования их в стратиграфии // Вопросы геологии Южного Урала и Нижнего Поволжья. Сборн. науч. трудов / Ред. Алёшечкин О.И. Саратов: Изд-во Саратовск. ун-та, 1990. С. 84–104.
Миних А.В., Миних М.Г. Ихтиофауна перми Европейской России. Саратов: Изд. центр “Наука”, 2009. 244 с.
Эйхвальд Э. Палеонтология России. Древний период. Ч. II. Фауна граувакковой, горноизвестковой и медистосланцевой формации России. СПб., 1861. 521 с.
Arefiev M.P., Golubev V.K., Balabanov Yu.P. et al. Type and reference sections of the Permian–Triassic continental sequences of the East European Platform: main isotope, magnetic, and biotic events. Sukhona and Severnaya Dvina Rivers field trip (XVIII Intern. Congr. on the Carboniferous and Permian). M.: PIN RAS, 2015. 104 p.
Bellwood D.R. Origins and escalation of herbivory in fishes: a functional perspective // Paleobiol. 2003. V. 29. P. 71–83.
Bellwood D.R., Hoey A.S., Bellwood O., Goatley C.H.R. Evolution of long–toothed fishes and the changing nature of fish–benthos interactions on coral reefs // Nature Commun. 2014. № 5. P. 31–44.
Dieleman J., Van Bocxlaer B., Manntschke C. et al. Tracing functional adaptation in African cichlid fishes through morphometric analysis of fossil teeth: exploring the methods // Hydrobiologia. 2015. V. 755. № 1. P. 73–88.
Esin D.N. Peculiarities of the trophic orientation changes in Palaeoniscoid assemblages from the Upper Permian of the European part of Russia // Modern Geol. 1997. V. 21. № 1. P. 185–195.
Pindakiewicz M., Tałanda M., Sulej T. et al. New finds of teeth of the herbivorous actinopterygians from the latest Permian of East European Platform and feeding convergence among extinct and extant ray-finned fish // Acta Palaeontol. Pol. (in press).
Purnell M., Seehausen O., Galis F. Quantitative three-dimensional microtextural analyses of tooth wear as a tool for dietary discrimination in fishes // J. Roy. Soc. Interface. 2012. V. 9. P. 2225–2233.
Yamaoka K. Feeding behaviour and dental morphology of algae scraping cichlids (Pisces: Teleostei) in Lake Tanganyika // Afr. Study Monogr. 1983. V. 4. P. 77–89.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал