

Палеонтологический журнал, 2020, № 2, стр. 65-72
Позднедевонские ботриолепидиды (Placodermi, Antiarchi) Тверской области
С. В. Молошников a, *, В. В. Линкевич b
a Музей землеведения МГУ им. М.В. Ломоносова
Москва, Россия
b Андреапольский районный краеведческий музей им. Э.Э. Шимкевича
Андреаполь, Россия
* E-mail: molsergey@rambler.ru
Поступила в редакцию 20.02.2019
После доработки 21.03.2019
Принята к публикации 09.04.2019
Аннотация
Из фаменских отложений (верхний девон) Тверской области описан вид Livnolepis heckeri (Lukševičs, 2001), отнесенный при первоописании к роду Bothriolepis. Новые материалы, собранные из типового местонахождения в окрестностях д. Билово, позволяют существенно дополнить морфологическое описание вида. У L. heckeri известны кости всей черепной крыши и почти всего туловищного панциря, а также пластинки проксимального сегмента грудного плавника. Некоторые кости представлены разными возрастными стадиями. В состав рода Livnolepis в настоящее время включены два вида: L. zadonica (H.Obrucheva, 1983) из нижнего фамена Центрального девонского поля и L. heckeri (Lukševičs, 2001) из среднего фамена юго-западной окраины Главного девонского поля. Наиболее характерными чертами строения экзоскелета ливнолеписов являются крупное преорбитальное углубление черепной крыши и слабо выраженные отпечатки переднебоковых отростков эндокрания на внутренней стороне черепной крыши.
В фаменских отложениях западной части Тверской области часто встречаются остатки антиарх из широко распространенного в девоне семейства Bothriolepididae (Молошников, Линкевич, 2017а, б; 2019). Предположительно в биловской свите обнаружены высокотелый гребенчатый Livnolepis heckeri (Lukševičs), а также Bothriolepis sp. с уплощенным панцирем без спинного гребня, в льнянской свите – B. cf. B. ciecere Lyarskaja. Среди ботриолепидид Тверской обл. преобладает Livnolepis heckeri, многочисленные остатки которого встречены около д. Билово на р. Малый Тудер. Это местонахождение было детально изучено Р.Ф. Геккером (Геккер и др., 1935). Здесь он выделил биловскую серию карбонатных пород, сложенную пестроцветными мергелями и зеленовато-серыми песчанистыми известняками и глинами, и соответствующую лебедянскому горизонту среднего фамена (Решение…, 1990).
Из биловского местонахождения Д.В. Обручев (Геккер и др., 1935, с. 74) определил остатки остео- и поролепиформных кистеперых и панцирных рыб, среди которых он установил новый вид ботриолеписа – Bothriolepis n. sp. Однако формально новый вид антиарх из Билово был описан только в 2001 г. Э.В. Лукшевичем (Lukševičs, 2001) и назван в честь Геккера как B. heckeri. Вид основан на трех экземплярах из сборов Геккера: одной почти целой и одной неполной передней среднеспинной кости, а также части боковой, краевой затылочной и заднекраевой в сочленении. При описании вида Лукшевич указал, что материал происходит предположительно из биловских слоев – Bilovo Beds (?) (Lukševičs, 2001, с. 577). Новые материалы, собранные, начиная с 2011 г., сотрудниками Андреапольского районного краеведческого музея им. Э.Э. Шимкевича (КМА) из типового местонахождения, позволили существенно дополнить описание этого вида и уточнить его систематическое положение. По ряду морфологических особенностей костей черепной крыши и туловищного панциря этот вид был отнесен к роду Livnolepis Moloshnikov (Молошников, 2016; Молошников, Линкевич, 2017а, б). В нижнефаменских отложениях Тверской обл. (Нелидовская и Редкинская скважины) ранее уже были встречены остатки ливнолеписов (Moloshnikov, 2008). Преимущественно остатки L. heckeri собирались из осыпи, что не позволяет уточнить их стратиграфическое положение. В настоящее время у L. heckeri известны кости всей черепной крыши и почти всего туловищного панциря, а также пластинки проксимального сегмента грудного плавника. Некоторые кости представлены разными возрастными стадиями. Голотип и два паратипа L. heckeri хранятся в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН) (колл. № 835), остальные изученные материалы в КМА и Музее землеведения МГУ им. М.В. Ломоносова.
Ниже L. heckeri описан по новым материалам, собранным в 2000-е гг. из типового местонахождения. При описании использовалась терминология, принятая в отечественной литературе. Измерения выполнялись по схемам, предложенным Э. Стенше (Stensiö, 1948).
Авторы глубоко благодарны проф. Л.И. Новицкой и О.Б. Афанасьевой (ПИН) за замечания и рекомендации по рукописи статьи.
ОТРЯД BOTHRIOLEPIFORMES
СЕМЕЙСТВО BOTHRIOLEPIDIDAE COPE, 1886
ПОДСЕМЕЙСТВО LIVNOLEPIDINAE MOLOSHNIKOV, 2012
Род Livnolepis Moloshnikov, 2008
Livnolepis heckeri (Lukševičs, 2001)
Табл. VIII , фиг. 1–8 ; табл. IX , фиг. 1–9 (см. вклейку)
Рис. 1.
Кости черепной крыши Livnolepis heckeri (Lukševičs): а – неполные левые боковая, краевая затылочная и заднекраевая кости в сочленении, экз. ПИН, № 835/40, снаружи; б, в – правая боковая кость, экз. № КМА/НВ/5184: б – снаружи, в – изнутри; г – передняя непарная кость, экз. № КМА/ НВ/5178, изнутри; д – затылочная кость, экз. № КМА/НВ/5173, снаружи; е – затылочная и заднепинеальная кости в сочленении, экз. № КМА/4770-1, снаружи. Обозначения: cir – полукруглая ямочная борозда, ifc1 – верхняя инфраорбитальная борозда, ifc.b – задняя косая головная ямочная борозда, L – боковая кость, nm – гладкая затылочная полоса, npp – заднепинеальная вырезка на nuchale, Nu – затылочная кость, orb – орбитальная (орбито-носовая) вырезка, PMg – заднекраевая кость, PNu – краевая затылочная кость, PP – заднепинеальная кость, prh – преорбитальное углубление, pr.po – переднебоковые углы отико-окципитального углубления, vcp – вертикальная сенсорная борозда. Масштабная линейка – 1 см.
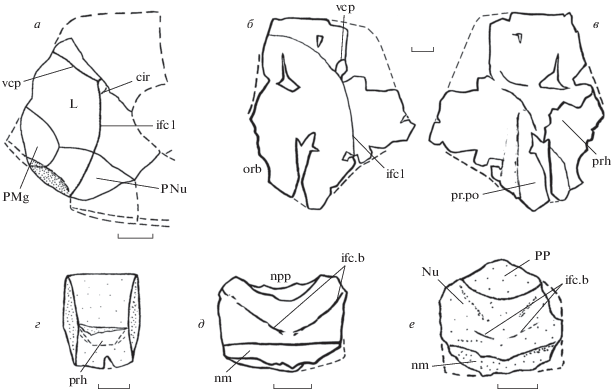
Рис. 2.
Кости туловищного панциря Livnolepis heckeri (Lukševičs): а–в – передняя среднеспинная, экз. № КМА/4049, наружная сторона: а – дорсально, б – латерально, в – спереди; г, д – задняя среднеспинная, экз. № КМА/НВ/5172, наружная сторона: г – дорсально, д – латерально; е, ж – неполная задняя среднеспинная, экз. № КМА/4770-8: е – снаружи, ж – изнутри; з – правая передняя спинно-боковая, экз. № КМА/НВ/5183, снаружи. Обозначения: cf.MxL – площадка, налегающая на mixilaterale, cr.d – срединный спинной гребень, cr.tp – заднее поперечное туловищное ребро, dlg2 – задняя косая туловищная борозда, dlm – спинная пластина, dlr – дорсолатеральное ребро, lcg – борозда главной боковой линии, llm – боковая пластина, oa.PMD – площадка, перекрывающаяся posterior medio-dorsale, pa – задний угол, pnoa – заднезатылочный угол, pr.ob – processus obstans, pt2 – задняя вентральная воронка на posterior medio-dorsale, tb – губчатое поле на posterior medio-dorsale. Масштабная линейка – 1 см.
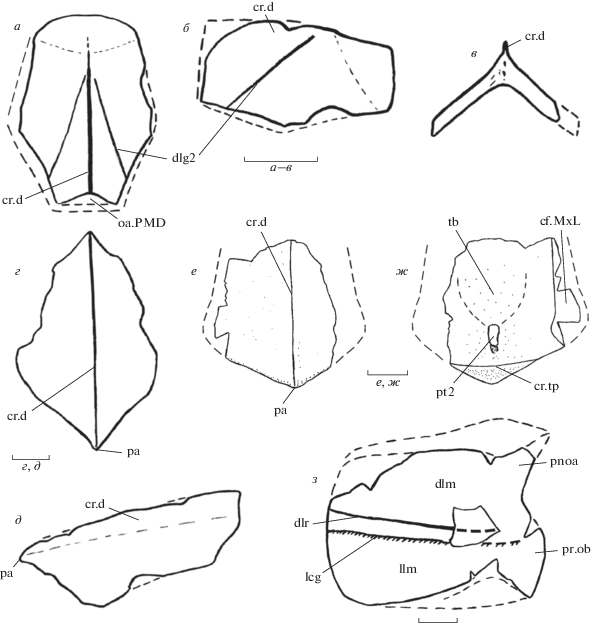
Рис. 3.
Наиболее характерные черты строения экзоскелета ливнолеписов, отличающие их от ботриолеписов: а, б – передние непарные кости, внутренняя сторона: а – Bothriolepis canadensis (Whiteaves), узкое щелевидное преорбитальное углубление (по Stensiö, 1948); б – Livnolepis zadonica (H.Obrucheva), крупное преорбитальное углубление (экз. ПИН, № 3725/1132); в, г – боковые кости, внутренняя сторона: в – Bothriolepis maxima Gross, хорошо выраженные глубокие передне-боковые углы отико-окципитального углубления (экз. ПИН б/н); г – Livnolepis zadonica (H.Obrucheva), слабо выраженные неглубокие передне-боковые углы отико-окципитального углубления (экз. ПИН, № 3525/1005). Обозначения: prh – преорбитальное углубление, pr.po – передне-боковые углы отико-окципитального углубления. Показано без масштаба.

Bothriolepis heckeri: Lukševičs, 2001, с. 577, рис. 60 H–J, 62 .
Livnolepis heckeri: Молошников, Линкевич, 2017а, рис. 3–8 ; 2017б, рис. 1, фиг. 2, 4–7 , рис. 2, фиг. 1, 2, рис. 3, фиг. 1, 3–5 ; 2019a, рис. 1–5 ; 2019б, рис. 1, 2а, 3.
Голотип – ПИН, № 835/42, передняя среднеспинная кость (anterior medio-dorsale); Тверская обл., р. Малый Тудер, окрестности д. Билово; верхний девон, фамен, биловская (?) свита.
Описание. Ботриолепидиды со сравнительно крупным и высоким панцирем, черепная крыша которого достигала в длину 12–13 см. Спинная стенка туловищного панциря могла вырастать в длину до 25 см, несет относительно высокий срединный спинной гребень (cr.d). Его максимальная высота, наблюдаемая на уровне шва между передней и задней среднеспинными костями, достигала 1 см и более.
Краниальный скелет. Боковая кость (laterale, L; рис. 1, а–в; табл. VIII , фиг. 1, 2) черепной крыши широкая, с широким и почти прямым ростральный краем. Отношение длины кости к ее ширине около 1.24. Боковые и задне-боковые углы кости выражены отчетливо. Как у взрослых (экз. № КМА/НВ/5184), так и у молодых особей (экз. № КМА/НВ/5182) верхняя инфраорбитальная борозда (ifc1) на наружной поверхности проходит посередине длины от бокового до орбитального и на расстоянии трети–половины длины от рострального до орбитального краев, т.е., боковой отдел laterale широкий. Вертикальная сенсорная борозда (vcp) длинная, хорошо выражена на костях как молодых, так и взрослых особей. Задняя косая головная ямочная линия (ifc.b) на боковых костях взрослых особей выражена слабо или отсутствует (рис. 1, б; табл. VIII , фиг. 1б); у молодых особей она отчетливая, заходит за среднюю линию орбитальной вырезки, но не достигает ее переднего края (табл. VIII , фиг. 2). На костях молодых особей также хорошо выражена полукруглая ямочная борозда (табл. VIII , фиг. 2: cir). На внутренней поверхности правой боковой кости (экз. № КМА/НВ/5184) переднебоковой угол отико-окципитального углубления (pr.po) неглубокий, выражен слабо; несколько шире в своем основании, чем соответствующий угол у L. zadonica. Преорбитальное углубление (prh) на боковой кости L. heckeri, по-видимому, такой же формы, что и у L. zadonica (Moloshnikov, 2004, 2008): развиты один боковой угол, а также боковой и передний края. Орбитальная (орбито-носовая: orb) вырезка неглубокая.
Передняя непарная кость (praemediale; рис. 1, г; табл. VIII , фиг. 3) удлиненная, с отношением длины к наибольшей ширине (вдоль переднего края) около 1.4. Передний край в 1.2 раза длиннее заднего. Передний край прямой, задний – слабовыпуклый. Боковые края слабовыпуклые. На экз. № КМА/НВ/5178 назальная пластина не сохранилась, но по участку ее прикрепления к фациальной пластине можно предположить, что преорбитальное углубление черепной крыши (prh) было крупным, как у L. zadonica. Судя по его форме на передней непарной и боковой кости (см. выше), L. heckeri, как и L. zadonica, обладал преорбитальным углублением трапециевидного типа.
Заднепинеальная кость (postpineale, PP; рис. 1, е; табл. VIII , фиг. 7 ) широкая, с выпуклым передним краем, отношение длины к ширине равно 0.6.
Затылочная кость (nuchale, Nu; рис. 1, д, е; табл. VIII , фиг. 4–7 ) широкая и сводчатая. Отношение длины кости к ширине около 0.7; угол сводчатости у заднего края равен 95°–100°. Кость достигает наибольшей ширины на уровне боковых и заднебоковых углов. Спереди nuchale несет неглубокую и широкую заднепинеальную вырезку (npp). Переднебоковые края, граничащие с laterale, прямые, короткие, в 2–3 раза короче заднебоковых. Заднебоковые края прямые или слабовогнутые. Вдоль заднего края на наружной поверхности проходит довольно широкая гладкая затылочная полоса (nm), переходящая на paranuchalia. На затылочных костях молодых особей хорошо развита задняя косая головная ямочная линия (if.b). Отверстия эндолимфатических протоков на наружной поверхности (d.end2) маленькие, расположены близко друг к другу (экз. № КМА/3666). На внутренней поверхности nuchale поперечное затылочное ребро высокое, развито по ширине всей кости, переходит на paranuchalia. Участок позади поперечного затылочного ребра длинный, с хорошо выраженным, невысоким медиальным окципитальным гребнем, сбоку от него могут косо проходить два невысоких симметричных дополнительных гребня.
Краевая затылочная кость (paranuchale, PNu; рис. 1, а) широкая. Верхняя инфраорбитальная борозда проходит на расстоянии трети–половины ширины кости от бокового края, т.е., как и на laterale, боковой отдел широкий.
Заднекраевая (postmarginale, PMg; рис. 1, а). При первоописании вида Лукшевич (Lukševičs, 2001, с. 578, рис. 62A ) указал, что эта кость удлиненная, с медиальными краями более длинными, чем боковые. Однако единственный экземпляр этой кости (ПИН, № 835/40) имеет обломанную заднебоковую часть, поэтому определить форму postmarginale довольно трудно. Возможно, что эта кость была все-таки широкой как у L. zadonica (см., напр., Moloshnikov, 2008, рис. 36, j–m ; табл. 8 , фиг. 5 ).
Посткраниальный скелет. Передняя среднеспинная кость (anterior medio-dorsale; рис. 2, а–в; табл. VIII , фиг. 8 ; табл. IX , фиг. 1) удлиненная и сводчатая на всем протяжении. Угол сводчатости у переднего края кости около 90°, у заднего – около 80°. У молодых особей anterior medio-dorsale c передним отделом относительно более широким, чем задний, имеет отношение длины к ширине около 1.3 (экз. № КМА/4049). Кости крупных особей могли иметь более широкий передний отдел и край (Lukševičs, 2001, рис. 62E ). Подобные возрастные изменения в пропорциях anterior medio-dorsale могут встречаться и у других ботриолепидид, например, у Bothriolepis sibirica Obruchev из франа Южной Сибири (Молошников, 2009, с. 81). Длина переднего отдела передней среднеспинной кости L. heckeri вдвое больше длины заднего отдела. Передний край кости слабо выпуклый, его длина в 1.4 раза меньше ширины кости на уровне боковых углов. Тергальный угол расположен на расстоянии трети длины кости от переднего края. Начиная от тергального угла, развит высокий дорсальный гребень (cr.d), достигающий максимальной высоты на месте шва между anterior и posterior medio-dorsalia. Задняя косая туловищная борозда (dlg2) на всех присутствующих в коллекции передних среднеспинных костях L. heckeri пересекает ее заднебоковой край, то есть она переходит на смешанно-боковую кость. В отличие от этого, для части экземпляров L. zadonica характерен переход задней косой туловищной борозды на заднюю среднеспинную кость (Moloshnikov, 2004, 2008). Швы передней среднеспинной кости с передней спинно-боковой и смешанно-боковой обычные для ботриолепидид: anterior medio-dorsale налегает на anterior dorso-laterale и покрывается mixilaterale на всем протяжении их контакта. Для некоторых экземпляров L. zadonica характерен нетипичный для ботриолепидид тип этих швов (Обручева, 1983; Moloshnikov, 2008).
Задняя среднеспинная кость (posterior medio-dorsale; рис. 2, г–ж; табл. IX , фиг. 2, 3) удлиненная, сводчатая на всем протяжении; с узким, длинным передним отделом; отношение длины кости к ее ширине около 1.4. Угол сводчатости у переднего края кости равен 120°. Переднебоковые углы сглажены, выражены слабо. Задний угол кости (pa) может быть оттянут назад, образуя задний отросток (экз. № КМА/НВ/5172; рис. 2, г; табл. IX , фиг. 2б) или выражен слабо (экз. № КМА/4770-8; рис. 2, е; табл. IX , фиг. 3). На внутренней поверхности posterior medio-dorsale задняя вентральная воронка (pt2) удлиненная, глубокая. Спереди от нее развито крупное губчатое поле (tb). Заднее поперечное ребро (cr.tp) невысокое, сглаженное; участок, расположенный позади от него, короткий (экз. № КМА/4770-8).
Передняя спинно-боковая кость (anterior dorso-laterale; рис. 2, з; табл. IX , фиг. 4 ) широкая, имеет несколько более узкую боковую пластину. Отношение ширины спинной пластины к ширине боковой около 1.08. Угол схождения этих пластин равен 145°. Дорсолатеральное ребро (dlr) сглажено, выражено слабо (экз. № КМА/НВ/5183). Кость несет узкий и удлиненный заднезатылочный угол (pnoa). Борозда главной боковой линии (lcg) выражена отчетливо. Передний выступ боковой пластины (processus obstans, pr.ob) слабо выдается вперед.
Смешанно-боковая кость (mixilaterale; табл. IX , фиг. 5 ) короткая и широкая. Так же как и у anterior dorso-laterale, ширина спинной пластины незначительно превышает ширину боковой. У молодых особей L. heckeri эти пластины сходятся под углом 120°–130°. Дорсолатеральное ребро (dlr) сглажено. Задний край mixilaterale слабо скошен, почти вертикальный. Задняя косая туловищная борозда (dlg2) и борозда главной боковой линии (lcg) выражены отчетливо.
Передняя вентро-латеральная кость (anterior ventro-laterale; табл. IX , фиг. 6, 7 ) с относительно узкой брюшной пластиной (vlm), отношение длины этой пластины к ее ширине около 1.5. Угол схождения боковой и брюшной пластин равен 125°. Вентро-латеральное ребро не выражено, сглажено. Субцефальный отдел anterior ventro-laterale широкий и короткий, занимает чуть меньше трети общей длины кости (экз. № КМА/НВ/5177). На наружной поверхности брюшной пластины anterior ventro-laterale развита брюшная поперечная сенсорная борозда (vg). На внутренней стороне кости постбранхиальное ребро (cr.ptbr) узкое, хорошо выражено только в боковой части кости, косо проходит от бокового края, до середины ширины брюшной пластины не доходит. Переднее внутреннее поперечное ребро (cit1) выражено слабо, имеет вид сглаженного валика.
Задняя вентро-латеральная кость (posterior ventro-laterale; табл. IX , фиг. 9 ) имеет сравнительно широкую боковую пластину с широким передним краем, и относительно узкую вентральную пластину. Длина боковой пластины почти в 2.3 раза превышает ее ширину. Угол схождения боковой и брюшной пластин равен 120°–130°. Вентро-латеральное ребро (vlr) выражено отчетливо только в задней части кости, в передней оно сглажено.
Проксимальный сегмент грудного плавника удлиненный (табл. IX , фиг. 6, 8 ), его длина превышает его ширину в 3.5 раза. Латеральный край сегмента выпуклый. Медиально-дорсальный и латеральный канты могут нести небольшие заостренные зубчики (экз. № КМА/4770-3). Медиально-вентральный кант сглаженный. Первая кость центрально-дорсального ряда (centrale-dorsale 1) широкая, отношение длины к ширине около 1.8.
Орнамент наружной поверхности костей взрослых особей L. heckeri бугорчатый. Бугорки крупные с округлой вершиной, на некоторых костях могут быть уплощенными. Обычно бугорки хорошо обособленны друг от друга, но могут и плотно примыкать друг к другу, например, на некоторых туловищных костях. Наружная поверхность костей молодых особей (напр., laterale, экз. № КМА/НВ/5182; anterior medio-dorsale, экз. № КМА/4049; mixilaterale, экз. № КМА/НВ/5175) несет ячеистый орнамент, который местами переходит в бугорчато-ребристый и бугорчатый. Вентральная стенка туловищного панциря и кости грудного плавника покрыты преимущественно ячеистым орнаментом; в узлах ячеек развиты бугорки и валики, образованные слившимися бугорками.
Размеры в мм. Praemediale, экз. КМА/НВ/5178: длина – 30, длина переднего края – 21.1, длина заднего края – 17.5; laterale, экз. КМА/НВ/5184: длина – 87.1, ширина – 70.5; postpineale, экз. КМА/4770-1: длина – 11, ширина – 18.5; nuchalia, экз. КМА/4770-1: длина – 24, ширина (восстановленная на уровне боковых углов) – ок. 31; экз. КМА/НВ/5173: длина – ок. 25.5, ширина (восстановленная на уровне боковых углов) – ок. 34; anterior medio-dorsale, экз. КМА/4049: длина – 28, ширина (на уровне боковых углов) – 22; posterior medio-dorsale, экз. КМА/НВ/5172: длина – 57.5, ширина (на уровне боковых углов) – 40.5; anterior dorso-laterale, экз. КМА/НВ/5183: длина – 56.5, ширина (с восстановленным pnoa) – ок. 44.5, ширина спинной пластины (с восстановленным pnoa) – 26, ширина боковой пластины – 24; mixilaterale, экз. КМА/НВ/5175: длина – ок. 21, ширина спинной пластины – ок. 15, ширина боковой пластины – ок. 13; anterior ventro-laterale, экз. КМА/НВ/5177: длина кости – ок. 100, ширина – ок. 65; posterior ventro-lateralia, экз. КМА/НВ/5181: длина боковой пластины – ок. 90, ширина боковой пластины – ок. 38; экз. КМА/НВ/5180: ширина вентральной пластины – 42.5; проксимальный сегмент грудного плавника, экз. КМА/НВ/5177: длина – 110, ширина – 30; centrale-dorsale 1, экз. КМА/НВ/5177: длина – 57, ширина – 31.5.
Сравнение. Судя по сводчатым затылочным, передним и задним среднеспинным костям, L. heckeri, как и L. zadonica, обладал сравнительно высоким для ботриолепидид панцирем, однако от последнего он отличается менее высоким спинным гребнем (cr.d) и пропорциями туловищного панциря. У L. heckeri, в отличие от L. zadonica, по-видимому, были более широкие боковые и более узкие спинно-боковые стенки туловищного панциря. Молодые особи L. heckeri имеют ячеистый орнамент наружной поверхности костей, на костях молодых L. zadonica развит гребенчатый и бугорчато-гребенчатый орнамент.
Материал. Из типового местонахождения: lateralia – 3; paranuchale – 1; фрагмент черепной крыши – 2; nuchalia – 5; anterior medio-dorsalia – 6; posterior medio-dorsalia – 4; фрагмент отдельного спинного гребня – 1; anterior dorso-lateralia – 2; mixilaterale – 1; anterior ventro-lateralia – 6; posterior ventro-lateralia – 5; проксимальный сегмент панциря грудного плавника – 8; отдельные кости из проксимального сегмента грудного плавника – 8; многочисленные фрагменты неопределимых посткраниальных костей.
ЗАКЛЮЧЕНИЕ
В состав рода Livnolepis в настоящее время включают два вида: L. zadonica (H. Obrucheva) из задонского горизонта (нижний фамен) Центрального девонского поля (ЦДП) и L. heckeri (Lukševičs) из среднего фамена юго-западной окраины Главного девонского поля. Наиболее характерными чертами строения экзоскелета ливнолеписов являются крупное преорбитальное углубление черепной крыши, а также слабо выраженные отпечатки переднебоковых отростков эндокрания на внутренней стороне черепной крыши (рис. 3). Ливнолеписы – довольно крупные ботриолепидиды, общая длина крыши черепа и туловищного панциря которых могла достигать 40 см. Туловищный панцирь обоих видов несет высокий срединный спинной гребень, который выполнял функцию стабилизатора тела антиарха при плавании, а, скорее всего, еще и киля (Молошников, 2010). В семействе Bothriolepididae подобные спинные гребни появились у франских представителей. Они известны, например, у австралийских видов Bothriolepis gippslandiensis и B. cullodonensis (Stensiö, 1948; Long, 1983; Long, Werdelin, 1986), а также у B. markovskii с западного склона Южного Урала (Молошников, 2010). Среди фаменских ботриолепидид высокий спинной гребень развит у ливнолеписов и B. cristata из Шотландии (Stensiö, 1948; Miles, 1968).
Низкие и широкие срединные спинные гребни туловищного панциря отмечены у шотландских видов B. wilsoni и B. obesa (Miles, 1968). Их спинные гребни, как отмечает Р. Майлс (Miles, 1968, с. 83), имеют тип строения, отличный от такового у B. cristata. По наличию низкого и широкого спинного гребня при построении кладограммы видов рода Bothriolepis Лукшевич (Lukševičs, 2001, рис. 83 ) сближает B. wilsoni с L. zadonica (ранее B. zadonica). Действительно, у голотипа L. zadonica (AMD, ПИН, № 1660/3) сохранился низкий и широкий спинной гребень. Однако при рассмотрении этого экземпляра видно, что гребень на нем обломан, а место его крепления стерто. Как можно судить по многим экземплярам среднеспинных костей из других местонахождений, у L. zadonica был развит высокий и узкий спинной гребень (см., напр., Moloshnikov, 2004, рис. 8 C, D ; 2008, табл. 9 , фиг. 3, 5 ), напоминающий гребень B. cristata, а также B. gippslandiensis и B. markovskii. Гребень подобного типа, но менее высокий, имеет и описанный L. heckeri. Поэтому филогенетическое сближение B. wilsoni с задонским видом по указанному признаку является сомнительным, тем более что появление срединного спинного гребня в туловищном панцире могло происходить независимо и параллельно в разных группах и видах ботриолепидид (Miles, 1968, с. 63; Молошников, 2010, с. 82).
Изученные новые материалы из фаменских отложений Тверской обл. дополняют данные о морфологии ботриолепидид, их систеиатическом составе и распространении представителей рода Livnolepis, что позволяет использовать их остатки для уточнения корреляции верхнедевонских отложений Восточно-Европейской платформы. Остатки крупных и высокотелых ботриолепидид, схожих с ливнолеписами, известны из франа ЦДП (Moloshnikov, 2008; неопубл. данные одного из авторов – М.С.В.) и нижнего фамена Центральной Польши (Szrek, 2004), а также из верхнедевонских отложений Южной Сибири (Молошников, 2010; Moloshnikov, 2012).
Список литературы
Геккер Р.Ф., Обручев Д.В., Филиппова М.Ф. Отложения Главного девонского поля. V–VII // Тр. Ленингр. геол.-гидро-геодезич. треста. 1935. Вып. 9. 82 с.
Молошников С.В. Новые данные о позднедевонских ботриолепидидах (Pisces, Antiarchi) Республики Тыва // Палеонтол. журн. 2009. № 5. С. 77–86.
Молошников С.В. О находках высокотелых ботриолепидид (Pisces, Placodermi, Bothriolepididae) в верхнем девоне Южного Урала и Кузбасса // Палеонтол. журн. 2010. № 5. С. 79–83.
Молошников С.В. К систематике эуантиарх (Vertebrata, Placodermi) // Наука в вузовском музее: Матер. Всеросс. научн. конф. (15–17 ноября 2016 г., Москва). Часть 2. М.: Музей землеведения МГУ, 2016. С. 26–29.
Молошников С.В., Линкевич В.В. Новые данные по позднедевонской ихтиофауне Тверской области (Билово, р. Малый Тудер) // Интегративная палеонтология: перспективы развития для геологических целей. Матер. 63 сессии Палеонтол. об-ва при РАН (3–7 апреля 2017 г., Санкт-Петербург). СПб., 2017а. С. 196–198.
Молошников С.В., Линкевич В.В. Систематическое положение фаменских антиарх (Vertebrata, Placodermi) Тверской области // Изв. высш. учебн. завед. Геол. и разведка. 2017б. № 2. С. 8–14.
Молошников С.В., Линкевич В.В. Морфология и систематическое положение некоторых фаменских ботриолепидид (Placodermi, Antiarchi) Тверской области // Морфологическая эволюция и стратиграфические проблемы. Матер. 65 сессии Палеонтол. об-ва при РАН (1-5 апреля 2019 г., Санкт-Петербург). СПб.: ВСЕГЕИ, 2019a. С. 249–251.
Молошников С.В., Линкевич В.В. К изменчивости ботриолепидид (Placodermi, Antiarchi): необычный экземпляр Livnolepis heckeri (Luksevics) из фамена Тверской области // Изв. высш. учебн. завед. Геол. и разведка. 2019б. № 4. С. 5–10.
Обручева Е.Д. Новый вид ботриолепид из отложений задонского горизонта Центрального девонского поля // Проблемы современной палеоихтиологии (Матер. конф., посвященной Д.В. Обручеву) / Ред. Новицкая Л.И. М.: Наука, 1983. С. 36–42.
Решение межведомственного регионального стратиграфического совещания по среднему и верхнему палеозою Русской платформы, с региональными стратиграфическими схемами. Л. 1988. Девонская система. Л., 1990. 58 с.
Long J.A. New bothriolepid fish from the late Devonian of Victoria, Australia // Palaeontology. 1983. V. 26. Pt 2. P. 295–320.
Long J.A., Werdelin L. A new Late Devonian bothriolepid (Placodermi, Antiarcha) from Victoria, with descriptions of other species from the state // Alcheringa. 1986. V. 10. № 4. P. 355–399.
Lukševičs E. Bothriolepid antiarchs (Vertebrata, Placodermi) from the Devonian of the north-western part of the East European platform // Geodiversitas. 2001. V. 23. № 4. P. 489–609.
Miles R.S. The Old Red Sandstone antiarchs of Scotland: family Bothriolepididae // Palaeontogr. Soc. Monogr. 1968. № 130. P. 1–130.
Moloshnikov S. Crested antiarch Bothriolepis zadonica H.D. Obrucheva from the Lower Famennian of Central European Russia // Acta Palaeontol. Pol. 2004. V. 49. № 1. P. 135–146.
Moloshnikov S.V. Devonian antiarchs (Pisces, Antiarchi) from Central and Southern European Russia // Paleontol. J. 2008. V. 42. № 7. P. 691–773.
Moloshnikov S.V. Middle–Late Devonian Placoderms (Pisces: Antiarchi) from Central and Northern Asia // Paleontol. J. 2012. V. 46. № 10. P. 1097–1196.
Stensiö E. On the Placodermi of the Upper Devonian of East Greenland II. Antiarchi: subfamily Bothriolepinae // Palaeozool. Groenl. 1948. Bd 2. 622 p.
Szrek P. The first articulated antiarch (Vertebrata, Placodermi) from the Upper Devonian of the Holy Cross Mountains (central Poland) // Acta Geol. Pol. 2004. V. 54. № 3. P. 401–406.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал