

Палеонтологический журнал, 2020, № 2, стр. 87-93
Ископаемая меламфаевая рыба рода Scopelogadus (Teleostei: Melamphaidae) из неогеновых отложений острова Сахалин, Россия
М. В. Назаркин a, *, А. Н. Котляр b, **
a Зоологический институт РАН
Санкт-Петербург, Россия
b Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
* E-mail: m_nazarkin@mail.ru
** E-mail: kotlyar@ocean.ru
Поступила в редакцию 30.04.2019
После доработки 06.05.2019
Принята к публикации 13.05.2019
Аннотация
Скелет берикоморфной рыбы из семейства Melamphaidae обнаружен в отложениях среднего-верхнего миоцена курасийской свиты острова Сахалин, Россия. Такие особенности морфологии, как короткое высокое тело с крупной головой, наличие одного спинного плавника с коротким основанием, 19 основных хвостовых лучей, небольшое число позвонков, однорядные челюстные зубы в сочетании с очень крупной циклоидной чешуей, позволяют относить это ископаемое к современному роду Scopelogadus. Это вторая известная в мире находка ископаемого скелета представителя как рода, так и семейства. Место этой находки расположено примерно на 17 градусов широты севернее ареала современных видов рода. Присутствие скопелогадусов в комплексе свиты кураси позволяет предполагать, что глубина накопления ее отложений составляла не менее 200 м.
Семейство меламфаевых (Osteichthyes, Beryciformes: Melamphaidae) объединяет 73 современных вида, сгруппированных в пять родов (Ebeling, 1962; Ebeling, Weed, 1963; Moore, 1993; Котляр, 1996, 2002, 2005, 2009, 2012, 2016; Nelson et al., 2016). Меламфаевые – исключительно морские пелагические глубоководные рыбы. Зрелые рыбы обычно встречаются на глубинах от 200 до 2000 м, и редко оказываются в более мелководных условиях (Ebeling, 1986; Котляр, 1996; Moore, Paxton, 1999). К последним приурочены, в основном, личинки и мальки. Меламфаевые обитают во всех океанах, кроме Северного Ледовитого. Отсутствуют они также в Средиземном море (Котляр, 1996; Moore, 2002). О биологии этих рыб известно немного. Меламфаевые питаются, главным образом, различными мелкими ракообразными (Bartow, 2010). А. Эбелинг и В. Вид (Ebeling, Weed, 1963, 1973) считали, что меламфаевые не совершают ясно выраженных суточных вертикальных пищевых миграций. Последующие исследования (Clarke, Wagner, 1976; Keene et al., 1987) показали, что такие миграции у них имеются.
Стандартная длина тела этих рыб (SL) достигает 187 мм у Poromitra crassiceps (Günther) (Moore, 2002). Морфологически они характеризуются довольно компактным вальковатым телом без светящихся органов; крупной головой с высоким тупым рылом и довольно большим ртом; широкими сенсорными головными каналами, покрытыми сверху тонкой кожей; редуцированной до двух передних пор боковой линией; единственным спинным плавником с коротким основанием; положением брюшных плавников под или впереди основания грудного плавника; невысоким (не более 31) числом позвонков; относительно крупной циклоидной чешуей (Котляр, 1991, 1996; Moore, Paxton, 1999; Moore, 2002).
Ископаемые находки представителей семейства довольно редки, и ограничиваются почти исключительно отолитами. Наиболее древние отолиты, принадлежащие Melamphaidae, описаны из палеоцена (танет) Баварии, и условно отнесены к роду Melamphaes Günther, 1864. Поскольку своим строением они напоминают отолиты современных Melamphaes, Scopeloberyx Zugmayer, 1911 и Sio Moss, 1962, данный палеоценовый вид рассматривается в качестве базального для остальных таксонов семейства (Schwarzhans, 2012). Отолиты Melamphaes spp. известны из нижнего и среднего эоцена Франции, нижнего и верхнего миоцена Новой Зеландии, верхнего миоцена и плиоцена Италии, плиоцена Франции, а также плиоцена и плейстоцена Калифорнии (Fitch, 1968; Grenfell, 1984; Nolf, 1985; Nolf, Cappetta, 1988; Patterson, 1993; Lin et al., 2016, 2017). Отолиты рода Scopeloberyx Zugmayer, 1911 известны из верхнего миоцена Новой Зеландии (Nolf, 1985), а отолиты рода Scopelogadus Vaillant, 1888 – из верхнего миоцена Италии и плейстоцена Калифорнии (Fitch, 1968; Nolf, 1985; Lin et al., 2017). Отолиты Poromitra Goode et Bean, 1883 обнаружены в нижнем миоцене Франции (Nolf, Brzobohaty, 2002). Отолиты из плейстоценовых отложений Калифорнии принадлежат уже современным видам меламфаид (Fitch, 1968; Fierstine et al., 2012).
Единственным ископаемым меламфаидом, описанным по скелетным остаткам к настоящему времени, является Scopelogadus capistranensis Ebeling из миоценовых отложений Калифорнии, США (Ebeling, 1962; Fierstine et al., 2012). Этот вид известен по двум небольшим экземплярам не более 40 мм стандартной длины (SL). В настоящей работе мы сообщаем о второй находке ископаемого скелета меламфаида, происходящего из миоценовых отложений курасийской свиты о-ва Сахалин.
Свита кураси, она же курасийская, распространена в юго-западной части Сахалина, где имеет мощность до 2300 м (Гладенков и др., 2002). Породы свиты сформировались в период среднего – начала позднего миоцена (Гладенков и др., 2002). Свита сложена тонкослоистыми алевролитами и аргиллитами темно-серого и коричневого цветов, выгорающими до почти белого с поверхности; нередко встречаются прослои крепких окремненных алевролитов и крупные эллиптические окремненные конкреции. В породах свиты кураси обнаружены многочисленные фоссильные остатки как беспозвоночных, так и позвоночных животных. Среди последних – кости морских млекопитающих, перья птиц и скелеты рыб. Таксономический состав миоценовой ихтиофауны курасийской свиты указывает на формирование этих неогеновых отложений в относительно глубоководных условиях, подходящих для мезопелагических рыб. Остатки этих последних составляют в свите кураси не менее 70% всех рыб (Назаркин, 2018). В собранной коллекции из нескольких сотен ископаемых из этой свиты скелет меламфаида – единственный. Степень сохранности этого скелета позволяет классифицировать ископаемое на родовом уровне, но не дает возможности достоверно определить его видовой статус. Поэтому далее мы приводим его описание в открытой номенклатуре.
ОПИСАНИЕ МАТЕРИАЛА ПО SCOPELOGADUS SP.
Скелет Scopelogadus sp., около 64.6 мм SL, происходит из берегового обрыва в 5 км к югу от пос. Пензенское, Томаринский р-н Сахалинской обл.; курасийская свита, средний–верхний миоцен; депонирован в палеоихтиологической коллекции Зоологического ин-та РАН, С.-Петербург, под номером ЗИН 451п.
Тело короткое, компактное и относительно высокое; наибольшая высота приходится, по-видимому, на уровень жаберной крышки. Голова большая; ее точная длина не известна, вероятно, она составляет около 47% SL (рис. 1). Рот большой, длина нижней челюсти не менее половины длины головы. Высота хвостового стебля около 7 раз в SL.
Рис. 1.
Scopelogadus sp., экз. ЗИН 451п, отпечаток (а) и контурный рисунок скелета (б); Сахалинская обл., Томаринский р-н, береговой обрыв в 5 км к югу от пос. Пензенское; средний–верхний миоцен, курасийская свита. Шкала 5 мм.

Границы и точную форму большинства краниальных костей не удается установить (рис. 2). Имеется несколько высоких субпрямоугольных infraorbitalia, тесно соединенных между собой; их точное число неизвестно. В передней части головы имеется крупное nasale с сильной передней вырезкой. Frontale широкое, субпрямоугольное; на его поверхности заметен слабо изогнутый узкий гребень, связанный с более короткими и неотчетливыми гребешками. В задней части нейрокрания различимо плотно прилегающее к первому позвонку exoocipitale. Parasphenoideum плоское и, по-видимому, широкое, однако его границы лишь угадываются. Другие кости черепа не различимы на имеющемся скелете.
Рис. 2.
Scopelogadus sp., экз. ЗИН 451п, контурный рисунок головы. Обозначения: aar – anguloarticulare; chy – ceratohyale; clt – cleithrum; cor – coracoideum; d – dentale; exo – exoccipitale; fr – frontale; hh – hypohyalia; hm – hyomandibulare; io – infraorbitalia; na – nasale; op – operculum; par – parasphenoideum; pc. r. – лучи грудного плавника; pcl – postcleithrum; pel – pelvis; pl. r. – лучи брюшного плавника; pmx – praemaxillare; pop – praeoperculum; ptt – posttemporale; q – quadratum; r – ребра; r.br. – radii branchiostegi; sc – scapula; scl – supracleithrum. Шкала 5 мм.
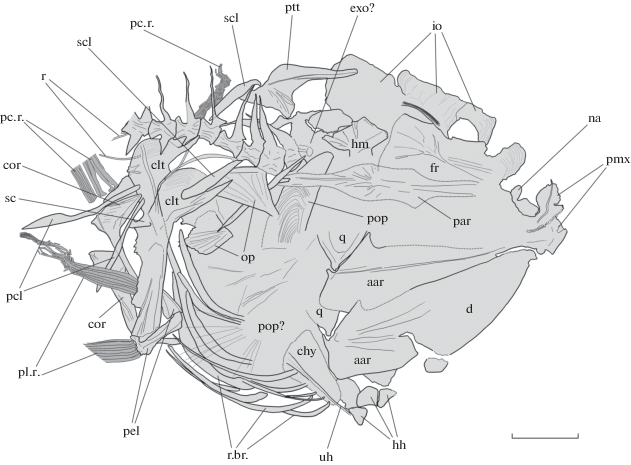
Верхняя часть hyomandibulare видна позади лобной кости; как и у современных сородичей (Котляр, 1991), она слабо подразделена на переднюю и заднюю суставные фасетки, и имеет отчетливый опрекулярный отросток. Кости крышечной серии сохранились лишь фрагментарно, однако тонкая радиальная струйчатость, присущая этим костям у современных видов рода, хорошо заметна. Quadratum вееровидное. В задне-нижней части головы угадывается широкое ceratohyale и три конусовидных hypohyalia. Различимо не менее 15 саблевидных жаберных лучей, передние из которых заметно тоньше задних. Вероятно, как и у современных сородичей, миоценовый вид обладал восемью лучами жаберной перепонки с каждой стороны.
В верхней челюсти сохранились передние участки prаemaxillaria, у которых имеется хорошо выраженный восходящий отросток. В передней части альвеолярного отростка сохранился ряд из 29 мелких конических зубов. Нижняя челюсть длинная и высокая, вероятно, немного вкладывается под верхнюю челюсть. Короноидный отросток anguloarticulare хорошо выражен, сочленовная фасетка узкая и глубокая. В передней части dentale имеется ряд из, по меньшей мере, 19 зубов сходного с верхнечелюстными зубами строения.
Позвонки слабо удлиненные, два-три передних несколько короче, все покрыты рельефом в виде тонких продольных и вертикальных гребней. В хвостовом отделе 14 позвонков, в туловищном, вероятно, 11. Длинные и широкие треугольные парапофизы имеются на 1-м–6-м сзади прекаудальных позвонках. Ребра слабо изогнуты, относительно широкие, наблюдаются с 4-го по предпоследний туловищный позвонок. Невральные и гемальные отростки высокие, обычно тонкие, но утолщены на передних четырех позвонках.
Suprаneuralia на сохранились. Спинной и анальный плавники смещены из их естественного положения, однако очевидно, что при жизни рыбы начало анального плавника было расположено несколько позади начала спинного. В спинном плавнике определенно 13 лучей и 12 проксимальных птеригиофоров. Первый луч, вероятно, колючий; членистые лучи слабо ветвятся, начиная с третьего. Высота наибольших лучей примерно равна длине основания плавника, однако последнее, очевидно, несколько увеличилось в процессе фоссилизации. В анальном плавнике один тонкий колючий и девять членистых, частью ветвистых лучей. Имеется девять проксимальных птеригиофоров анального плавника. Как и в спинном плавнике, это удлиненные, дистально расширенные, палочковидные элементы, постепенно укорачивающиеся назад. Позади 9-го птеригиофора расположен один четырехугольный костный элемент, у которого нет собственного луча (рис. 1). Возможно, этот элемент также входит в серию проксимальных птеригиофоров анального плавника.
Грудной плавник с узким основанием, содержит не менее 13 длинных тонких лучей. Его основание расположено на уровне 5-го прекаудального позвонка, а концы его лучей достигают уровня 5-го хвостового позвонка, и значительно заходят назад за начало анального плавника. Cleithrum высокое, изогнутое, с широкой дорсальной лопастью (рис. 2). Coracoideum длинное и широкое, его задний отросток (Котляр, 1991) умеренно развит; scapula значительно меньше, с крупным овальным отверстием. Плавниковые radialia и дорсальное postcleithrum не сохранились. Вентральное postcleithrum длинное и узкое, но отчетливо расширяется дистально. Posttemporale очень крупное, с широкой медиальной площадкой в задней части. Supracleithrum палочковидное.
Брюшной плавник узкий, значительно короче грудного, состоит из тонкого короткого колючего и 7 членистых лучей. Его основание расположено несколько впереди основания грудного. Длина плавника примерно соответствует длине пяти туловищных позвонков. Тазовая кость плоская, широкая, со слабым медиальным отростком в задней части.
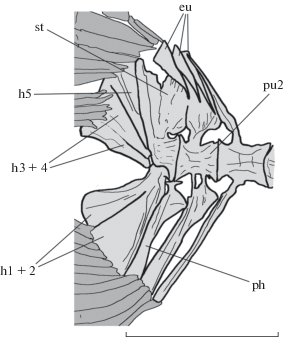
Хвостовой плавник вильчатый, его длина заметно меньше длины грудного плавника. Имеется 10 + 9 основных и по два дополнительных укороченных луча сверху и снизу. Строение хвостового скелета соответствует таковому современных видов рода (Бородулина, 1989; Котляр, 1991). Имеется единое комплексное терминальное тело позвонка, с которым срастается высокое и длинное stegurale (рис. 3). К задней части терминального тела примыкают автономное parhypurale и гипуральные пластинки: hypurale 5 и частично слившиеся между собой hypurale 1 + 2 и hypurale 3 + 4. Uroneurale не различимо. Epuralia клиновидные, переднее длиннее, а заднее шире остальных. Невральная дуга второго преурального позвонка короткая и широкая; автономна ли его гемальная дуга, не удается установить.
Рис. 3.
Scopelogadus sp., экз. ЗИН 451п, контурный рисунок хвостового скелета. Обозначения: eu – epuralia; h – hypuralia; ph – parhypurale; pu2 – второй преуральный позвонок; st – stegurale. Шкала 5 мм.
Тело покрыто очень крупной овальной циклоидной чешуей. На переднем поле отдельной чешуи развиты 3–5 крупных фестонов, в то время как заднее поле либо гладкое, либо подразделено на три крупные лопасти, медиальная из которых короче и уже латеральных. Фокус расположен в центре чешуйной пластинки. В целом, строение чешуи ископаемого сходно с таковым современных видов (рис. 4). Длина отдельных чешуй, различимых на ископаемом, колеблется в пределах 9.2–12.4 мм и достигает 19.2% SL.
Рис. 4.
Боковые чешуи Scopelogadus sp., экз. ЗИН 451п (а) и современного S. bispinosus (Gilbert) (б). Передний край слева. Шкала 5 мм.

Размеры в мм. Длина скелета (SL, в расправленном состоянии) 64.6; длина головы около 30.6; высота хвостового стебля 8.9; длина грудного плавника 17.5, брюшного плавника 9.6, хвостового плавника 15.9; высота спинного плавника 11.8, анального плавника 11.0; длина основания спинного плавника 11.6, анального плавника 7.5.
* * *
Как видно из приведенного выше описания, ископаемая рыба из свиты кураси характеризуется вальковатым телом с очень крупной головой, длинными челюстями, мелкими челюстными зубами, спинным и анальным плавниками с короткими основаниями, слабыми колючими лучами в начале вертикальных плавников, одним колючим и семью членистыми лучами в брюшном плавнике, 19-ю основными лучами хвостового плавника, положением основания брюшного плавника впереди основания грудного, и очень небольшим числом позвонков. Перечисленные признаки позволяют однозначно классифицировать это ископаемое как представителя семейства Melamphaidae. Характерное количество позвонков и число лучей в плавниках, а также наличие крупной циклоидной чешуи свидетельствуют о принадлежности миоценовой рыбы к роду Scopelogadus.
К роду Scopelogadus относят четыре современных (Котляр, 2019) и один вымерший (Ebeling, 1962) вид (табл. 1). Виды этого рода отличаются от других представителей семейства чрезвычайно крупной чешуей: число рядов чешуй в продольном ряду от виска до начала хвостового плавника не более 15. Вероятно, в относительном выражении скопелогадусы обладают самой крупной среди костистых рыб чешуей циклоидного типа.
Таблица 1.
Основные счетные признаки видов Scopelogadus. Данные по современным видам из: Бородулина, 1989; Котляр, 2019; Котляр, 2020, в печати
| †Scopelogadus sp. | †S. capistranensis | S. beanii | S. mizolepis | S. bispinosus | S. unispinis | |
|---|---|---|---|---|---|---|
| V.a. | ~11 | 10 | 10 | 10 | 10–11 | 10–11 |
| V.c. | 14 | 14 | 15–17 | 14–16 | 14–15 | 12–13 |
| V.T. | ~25 | 24 | 25–27 | 24–26 | 24–26 | 23 |
| D1 | I? | ? | I–II | II | II | I |
| D2 | 12 | 12–13 | 10–12 | 10–12 | 10–12 | 10–11 |
| A1 | I | ? | I | I | I | I |
| A2 | 9 | 9–10 | 7–9 | 7–9 | 7–9 | 7–8 |
| P | >=13 | ~12–15 | 13–16 | 14–15 | 12–15 | 14–15 |
| V | I, 7 | ~8 | I, 7 | I, 7 | I, 7 | I, 7 |
Как современные, так и ископаемые виды рода чрезвычайно сходны между собой пропорциями тела, числом позвонков и лучей в плавниках (табл. 1). Имеющиеся данные по скопелогадусу из свиты кураси не позволяют обособить его от сородичей на видовом уровне, поскольку все его основные счетные признаки и известные пропорции перекрываются с таковыми у других видов. Ископаемые Scopelogadus spp., таким образом, показывают, что уже к среднему миоцену этот род сформировался в своих основных особенностях и, кроме того, был широко расселен в северной части Тихого океана.
Ископаемый Scopelogadus sp. из миоцена Сахалина интересен тем, что место его находки (47° с.ш.) расположено гораздо севернее ареала современных видов, а также местонахождения миоценового вида в Калифорнии (27° с.ш.). Ближайший современный сородич, S. mizolepis (Günther), в северо-западной части Тихого океана не встречается севернее 30° с.ш., т.е. не достигает Южной Японии (Котляр, 2020, в печати). Присутствие скопелогадуса в комплексе ископаемых свиты кураси позволяет предположить, что накопление осадков свиты происходило на глубине не менее 200 м.
Список литературы
Бородулина О.Д. Идентификация остатков мезопелагических рыб из желудков хищников. Сообщение VII. Особенности осевого скелета массовых рыб семейства Melamphaeidae // Вопр. ихтиол. 1989. Т. 29. Вып. 1. С. 165–167.
Гладенков Ю.Б., Баженова О.К., Гречин В.И. и др. Кайнозой Сахалина и его нефтегазоносность. М.: Геос, 2002. 223 с.
Котляр А.Н. Остеология рыб подотряда Stephanoberycoidei. Сообщение 2. Melamphaidae // Вопр. ихтиол. 1991. Т. 31. Вып. 1. С. 24–39.
Котляр А.Н. Бериксообразные рыбы Мирового океана. М.: Изд-во ВНИРО, 1996. 368 с.
Котляр А.Н. Sio nordenskjoldii (Melamphaidae): морфологическая характеристика и распространение // Вопр. ихтиол. 2002. Т. 42. Вып. 1. С. 32–40.
Котляр А.Н. Ревизия рода Scopeloberyx (Melamphaidae). Часть 3. Виды группы S. opisthopterus // Вопр. ихтиол. 2005. Т. 45. Вып. 1. С. 21–32.
Котляр А.Н. Ревизия рода Poromitra (Melamphaidae). Часть 5. Виды групп P. capito и P. crassa // Вопр. ихтиол. 2009. Т. 49. Вып. 6. С. 732–744.
Котляр А.Н. Ревизия рода Melamphaes (Melamphaidae). Часть 4. Многотычинковые виды: M. ebelingi, M. occlusus sp. nova, M. nikolayi sp. nova // Вопр. ихтиол. 2012. Т. 52. Вып. 3. С. 279–287.
Котляр А.Н. Ревизия рода Melamphaes (Melamphaidae). 7. Малотычинковые виды: M. danae и M. pumilis // Вопр. ихтиол. 2016. Т. 56. Вып. 4. С. 379–390.
Котляр А.Н. Ревизия рода Scopelogadus (Melamphaidae). 1. S. beanii // Вопр. ихтиол. 2019. Т. 59. Вып. 5. С. 507–522.
Котляр А.Н. Ревизия рода Scopelogadus (Melamphaidae). 2. S. mizolepis // Вопр. ихтиол. 2020. Т. 60. № 1. C. 3–15.
Назаркин М.В. Особенности миоценовой ихтиофауны Дальнего Востока России // Биологические проблемы Севера. Матер. междунар. научн. конф. памяти В.Л. Контримавичуса. 18–22 сентября 2018. Магадан: Изд-во ИБПС ДВО РАН, 2018. С. 445–447.
Bartow K.A. Taxonomy and ecology of the deep-pelagic fish family Melamphaidae, with emphasis on interactions with a mid-ocean ridge system. PhD Dissertation. Boca Raton: Florida Atlantic Univ., 2010. 185 p.
Clarke T.A., Wagner P.J. Vertical distribution and other aspects of the ecology of certain mesopelagic fishes taken near Hawaii // Fish. Bull. 1976. V. 74. № 3. P. 635–647.
Ebeling A.W. Melamphaidae I. Systematics and zoogeography of the species in bathypelagic fish genus Melamphaes Günther // Dana Rep. 1962. № 58. P. 1–164.
Ebeling A.W. Family Melamphaidae // Smiths’ Sea Fishes / Eds. Smith M.M., Heemstra P.C. Johannesburg: Macmillan South Africa, 1986. P. 427–431.
Ebeling A.W., Weed W.H. Melamphaidae III. Systematics and distribution of the species in the bathypelagic fish genus Scopelogadus Vaillant // Dana Rep. 1963. № 60. P. 1–58.
Ebeling A.W., Weed W.H. Order Xenoberyces (Stephanoberyciformes) // Fishes of the Western North Atlantic. New Haven: Yale Univ., 1973. P. 397–478 (Mem. Sears Found. Mar. Res. № 1. Pt 6).
Fierstine H.L., Huddleston R.W., Takeuchi G.T. Catalog of the Neogene bony fishes of California. A systematic inventory of all published accounts // Occas. Pap. Calif. Acad. Sci. 2012. № 159. 206 p.
Fitch J.E. Otoliths and other fish remains from the Timms Point Silt (early Pleistocene) at San Pedro, California // Contrib. in Sci. LA Cty Mus. 1968. № 146. P. 1–29.
Grenfell H.R. Early Miocene teleost otoliths from Parengarenga Harbour, New Zealand // New Zeal. J. Geol. Geophys. 1984. V. 27. P. 51–96.
Keene M.J., Gibbs R.H., Jr., Krueger W.H. Family Melamphaidae, Bigscales // Biology of midwater fishes of the Bermuda Ocean Acre / Eds. Gibbs R.H., Jr., Krueger W.H. Washington D.C., 1987. P. 169–187 (Smithson. Contrib. Zool. № 452).
Lin C.-H., Brzobohatý R., Nolf D., Girone A. Tortonian teleost otoliths from northern Italy: taxonomic synthesis and stratigraphic significance // Eur. J. Taxonomy. 2017. V. 322. P. 1–44.
Lin C.-H., Nolf D., Steurbaut E., Girone A. Fish otoliths from the Lutetian of the Aquitaine Basin (SW France), a breakthrough in the knowledge of the European Eocene ichthyofauna // J. Syst. Palaeontol. 2016. https://doi.org/10.1080/14772019.2016.1246112
Moore J.A. Phylogeny of the Trachichthyiformes (Teleostei: Percomorpha) // Bull. Mar. Sci. 1993. V. 52. P. 114–136.
Moore J.A. Melamphaidae // FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of the WCA. Vol. 2. Bony fishes part 1 (Acipenseridae to Grammatidae) / Ed. K.E. Carpenter. Rome: FAO, 2002. P. 1162–1163.
Moore J.A., Paxton J.R. Melamphaidae. Bigscales (ridgeheads) // FAO species identification guide for fishery purposes. The living marine resources of the WCP. Vol. 4. Bony fishes part 2 (Mugilidae to Carangidae) / Eds. Carpenter K.E., Niem V.H. Rome: FAO, 1999. P. 2201–2202.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken, NJ: Wiley, 2016. 707 p.
Nolf D. Otolithi Piscium // Handbook of paleoichthyology. V. 10 / Ed. H.P. Schultze. Stuttgart: G. Fischer, 1985. 145 p.
Nolf D., Brzobohatý R. Fish otoliths from the Saubrigues paleocanyon (Chattian to Langhian), Aquitane, France // Rev. Micropaléontol. 2002. V. 45. P. 261–296.
Nolf D., Cappetta H. Otolithes de poisson Pliocène du Sud-Est de la France // Bull. Inst. Sci. Natur. Belg. Sci. Terre. 1988. V. 58. P. 209–271.
Patterson C. Osteichthyes. Teleostei // The Fossil Record 2 / Ed. M.J. Benton. L.: Chapman & Hall, 1993. P. 621–656.
Schwarzhans W. Fish otoliths from the Paleocene of Bavaria (Kressenberg) and Austria (Kroisbach and Oiching-Graben) // Palaeoichthyologica. 2012. V. 12. P. 1–88.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал