

Палеонтологический журнал, 2020, № 2, стр. 35-39
Новые диктионевридовые (Insecta: Dictyoneurida Handlirsch = Palaeodictyoptera Goldenberg) из пермских отложений Воркутского бассейна
Н. Д. Синиченкова *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: nina_sin@mail.ru
Поступила в редакцию 05.03.2019
После доработки 11.03.2019
Принята к публикации 11.03.2019
Аннотация
Из пермских насекомоносных отложений Воркутского угольного бассейна описано два новых вида Homaloneura brauckmanni sp. nov. и Baeoneura desperata sp. nov., принадлежащих семейству Spilapteridae. Для Vorkutoneura variabilis Sinitshenkova, 1977 из того же семейства впервые описаны переднее имагинальное и заднее нимфальное крылья. Комплекс диктионевридовых противоречит определению позднепермского возраста отложений, по насекомым возраст изученных отложений скорее соответствует ранней перми.
В новых материалах из насекомоносных отложений перми Воркутского бассейна обнаружены, помимо других насекомых, представители отряда Dictyoneurida Handlirsch, 1906 (=Palaeodictyoptera Goldenberg, 1854). Это один из самых древних и разнообразных отрядов крылатых насекомых, включающий в себя около 200 видов, более 40 родов из 21 семейства. Диктионевридовые широко распространены в карбоне и перми (Carpenter, 1992; Sinitshenkova, 2002). Эти насекомые, размером от средних до очень крупных, имели две пары почти гомономных крыльев с широким основанием, которые никогда не складывались, а держались распростертыми в стороны. Помимо многочисленных продольных жилок, некоторые представители отряда имели густую сеть поперечных жилок, или архедиктий. Для диктионевридовых характерен длинный сосущий хоботок. Развивались они на суше по типу археметаболии, а их преимагинальные стадии обитали, как и взрослые, в кронах деревьев (Синиченкова, 1980).
Представители отряда известны с конца раннего карбона, но достигли процветания в среднем и особенно верхнем карбоне. Распространены они в основном в Северном полушарии, но наибольшее их разнообразие известно в Еврамерии. Единичные находки диктионевридовых отмечены в Аргентине, Австралии на Тасмании, Восточной Сибири и Монголии (Carpenter, 1992; Sinitshenkova, 2002). В перми диктионевридовые становятся редкими и концу этого периода исчезают. Ни одно семейство отряда не пересекает пермо–триасовую границу.
Отряд Dictyoneurida делится на два подотряда – Dictyoneurina Handlirsch, 1906 и Frankenholziina Guthörl, 1962. Первый включает шесть надсемейств с 27 семействами, а второй менее разнообразный – входящие в его состав два надсемейства включают 10 семейств (Sinitshenkova, 2002; Prokop et al., 2017). Описанные ниже диктионевридовые из Воркутского бассейна отнесены к семейству Spilapteridae Brongniart, 1893, которое входит в надсемейство Spilapteroidea Brogniart, 1893 подотряда Dictyoneurina.
Из Воркутского бассейна диктионевридовые были ранее известны по трем видам из семейства Spilapteridae: Dunbaria borealis Sharov et Sinitshenkova, 1977, D. quinquefasciata Sharov et Sinitshenkova, 1977 и Vorkutoneura variabilis Sinitshenkova, 1977 (Шаров, Синиченкова, 1977).
Новые виды из этого же семейства принадлежат родам Homaloneura Brongniart, 1885 и Baeoneura Sinitshenkova, 1977. Для Vorkutoneura variabilis, описанного первоначально по строению заднего крыла имаго, найдены и впервые описываются переднее имагинальное и заднее нимфальное крылья.
Представители Homaloneura и Baeoneura до сих пор были известны из более древних отложений, датируемых поздним карбоном: первый – из Европы (Kukalova, 1969; Carpenter, Richardson, 1971; Prokop et al., 2016, 2017), второй – из Сибири (Шаров, Синиченкова, 1977). Новый вид Homaloneura brauckmanni sp. nov. найден в аячьягинской подсвите лекворкутской свиты, а Baeoneura desperata sp. nov. в интинской свите. Описанные в статье новые виды являются наиболее северными и наиболее поздними из известных представителей Homaloneura и Baeoneura.
Сравнительно разнообразная фауна диктионевридовых в Воркутском бассейне, а также находка мисхоптеридовых, близких к карбоновым видам (Синиченкова, 2019), противоречит определению возраста насекомоносных толщ как позднепермского. Выводы по изученным группам насекомых (Расницын и др., 2005; Лозовский и др., 2009) лучше согласуются с прежними возрастными оценками насекомоносных слоев, когда вся лекворкутская и нижняя часть интинской свиты относились к низам уфимского яруса (Македонов, Погоревич, 1966). Такой вывод противоречит схеме Н.С. Муравьева (1988), по которой воркутский ярус, включающий интинскую и большую часть лекворкутской свиты, относится к верхней перми, лишь низы лекворкутской свиты остаются в нижнепермском кунгурском ярусе. К кунгуру всю воркутскую серию относили и другие авторы (Пухонто, 1998).
Работа частично поддержана Программой Президиума РАН № 30 “Эволюция органического мира и планетарных процессов” и грантом РФФИ № 18-04-00322.
ОТРЯД DICTYONEURIDA (=PALAEODICTYOPTERA)
ПОДОТРЯД DICTYONEURINA
НАДСЕМЕЙСТВО SPILAPTEROIDEA BROGNIART, 1893
СЕМЕЙСТВО SPILAPTERIDAE BRONGNIART, 1893
Род Homaloneura Brongniart, 1885
Homaloneura brauckmanni Sinitshenkova sp. nov.
Табл. V , фиг. 1 (см. вклейку)
Рис. 1.
Homaloneura brauckmanni sp. nov., голотип ПИН, № 1631/397, переднее имагинальное крыло; Воркутский бассейн, нижняя пермь. Длина масштабной линейки 10 мм.
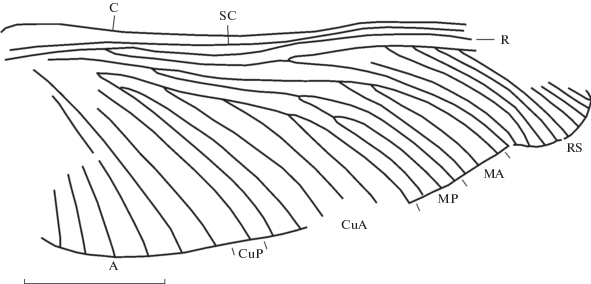
Название вида в честь палеоэнтомолога К. Браукманна.
Голотип – ПИН, № 1631/397, прямой и обратный отпечатки почти целого переднего имагинального крыла; Республика Коми, Хальмеръюское месторождение, скв. ХК-966, гл. 355.4 м (пакет N); нижняя пермь, лекворкутская свита, рудницкая подсвита.
Описание (рис. 1). Переднее имагинальное крыло. Передний край крыла немного прогнут посередине; костальное и субкостальное поля почти равны по ширине, в основании крыла костальное поле значительно расширяется. RS отходит от R в базальной четверти крыла, гребенчатый назад, десятиветвистый, его первое ветвление на уровне середины крыла. М ветвится в базальной трети крыла, МА и МР трехветвистые, МА гребенчатая назад, начинает ветвиться дистальнее первого развилка RS, МР гребенчатая вперед, ветвится немного базальнее первого развилка МА. Передняя ветвь CuA гребенчатая назад, четырехветвистая, ее задняя ветвь дихотомирует близ своего основания; CuP простая. Анальные жилки простые, не менее шести. Поперечные жилки на отпечатке немногочисленные, плохо заметны из-за особенностей сохранности.
Размеры в мм: длина фрагмента переднего имагинального крыла (голотип) 40, его полная длина около 42, его наибольшая ширина 15.
Сравнение. По размерам H. brauckmanni sp. nov. сравним лишь с H. bonnieri из Верхнего карбона Франции, длина переднего крыла которого составляет 43 мм. Остальные известные виды почти вдвое мельче. От H. bonnieri новый вид отличается меньшим числом поперечных жилок, меньшим числом ветвей MP, CuA, простой CuA и простыми анальными жилками. У H. bonnieri четыре ветви MP и восемь ветвей CuA, анальные ветвятся.
Замечание. Описываемый вид отнесен к роду Homaloneura на основании типичной формы крыла с суженной вершиной и расширенной анальной частью, многоветвистыми RS, M и CuA, наличия многочисленных анальных жилок.
Материал. Кроме голотипа, паратип ПИН, № 1631/398 – фрагмент имагинального крыла; Воркутское месторождение, скв. К-1348, гл. 1140.20 м; нижняя пермь, лекворкутская свита, аячьягинская подсвита.
Род Baeoneura Sinitshenkova, 1977
Baeoneura desperata Sinitshenkova, sp. nov.
Табл. V , фиг. 2, 3
Рис. 2.
Baeoneura desperata sp. nov., голотип ПИН, № 1631/402, фрагмент переднего крыла; Воркутский бассейн, нижняя пермь. Длина масштабной линейки 5 мм.
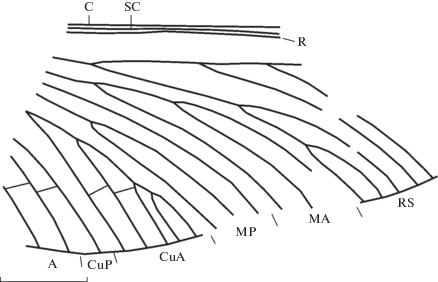
Рис. 3.
Vorkutoneura variabilis Sinitshenkova, 1977, экз. ПИН, № 1631/391, переднее имагинальное крыло; Воркутский бассейн, нижняя пермь. Длина масштабной линейки 10 мм.
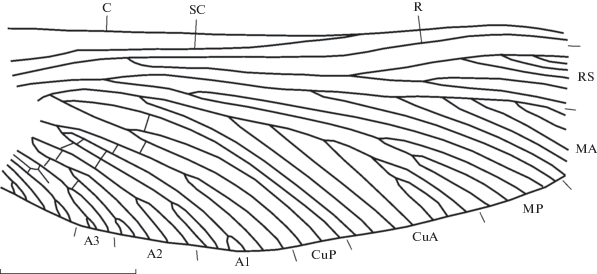
Название вида от desperatus лат. – безнадежный.
Голотип – ПИН, № 1631/402, прямой и обратный отпечатки фрагмента переднего имагинального крыла; Республика Коми, Воркутское месторождение, скв. К-1304, гл. 626.0 м; нижняя пермь, воркутская серия, интинская свита.
Описание (рис. 2). Переднее имагинальное крыло. C, SC и R сближены, но не сливаются. Радиальное поле очень широкое; передняя ветвь RS гребенчатая назад, несет не менее трех ветвей, задняя ветвь гребенчатая вперед, трехветвистая. Передняя ветвь RS начинает ветвиться немного базальнее первого развилка задней ветви. MA и MP трехветвистые, первый развилок MA значительно дистальнее первого развилка MP, задняя ветвь МА простая, передняя делится один раз. Передняя ветвь MP простая, задняя ветвится заметно базальнее деления передней ветви МА. CuA пятиветвистая, ее задняя ветвь простая, передняя ветвится почти дихотомически трижды. CuP и анальные простые.
Размеры в мм: длина фрагмента переднего крыла 26, полная длина крыла около 36.
Сравнение. От единственного ранее известного вида B. obscura Sinitsh., 1977 из верхнего карбона Тунгусского бассейна новый вид отличается более крупными размерами, более широким радиальным полем, большим числом ветвей MA и CuA.
Замечание. Описываемый вид отнесен к роду Baeoneura на основании широкого радиального поля, гребенчатых ветвей RS, многоветвистой CuA и простой CuP.
Материал. Голотип.
Род Vorkutoneura Sinitshenkova, 1977
Vorkutoneura variabilis Sinitshenkova, 1977
Табл. V , фиг. 4–6
Vorkutoneura variabilis: Шаров, Синиченкова, 1977, с. 60, табл. VI , фиг. 7 , рис. 5 .
Голотип – ПИН, № 1631/2; Нижне-Сырьягинское месторождение (скв. НСК-83, гл. 162.8 м); нижняя пермь.
Описание (рис. 3). Переднее имагинальное крыло. Передний край крыла прогнут, костальное поле в базальной части крыла вдвое шире субкостального, в середине крыла ширина этих полей одинакова. C, SC и R сильно сближены лишь на вершине крыла. Радиальное поле в дистальной половине крыла вдвое шире костального и субкостального. RS отходит от R в базальной четверти крыла. Первый развилок RS располагается дистальнее середины крыла, его передняя ветвь гребенчатая назад четырехветвистая, задняя – гребенчатая вперед двуветвистая. М ветвится немного базальнее основания RS; MA гребенчатая назад с шестью ветвями, MP четырехветвистая, ее ветви отходят вперед, первый развилок МР значительно базальнее первого развилка MA, задняя ветвь МР ветвится дихотомически. CuA семиветвистая, все ее ветви отходят назад, CuP трехветвистая, ее ветви отходят вперед, причем первая ветвь отходит значительно базальнее первого развилка CuA. Анальные жилки ветвятся дихотомически. Две задние ветви A1 с развилками; А2 впадает в край крыла пятью ветвями, А3 – тремя, А4 и А6 с короткими вилками, А5 простая. Поперечные жилки почти не заметны на отпечатках.
Заднее нимфальное крыло. Анальная часть крыла расширена. C, SC и R образуют единое утолщение на переднем крае крыла. Основание M и Cu, а также их передние ветви сильно утолщены. Характер ветвления продольных жилок сходен с таковым заднего имагинального крыла.
Размеры в мм: длина фрагмента переднего имагинального крыла 42, его полная длина около 44, его наибольшая ширина 16.7; длина заднего нимфального крыла 8, его наибольшая ширина 3.8.
Замечания. Первоначально V. variabilis был описан по задним имагинальным крыльям также из Воркутского бассейна – голотип ПИН, № 1631/2 и паратип ПИН, № 1631/33 из Нижне-Сырьягинского месторождения (скв. НСК-83, гл. 162.8 м) и другой паратип ПИН, № 1631/ 18 из Верхне-Сырьягинского месторождения (скв. ВСК-133, гл. 285.19–286.0 м) (Шаров, Синиченкова, 1977). По длине заднее и переднее крылья сходны, длина заднего крыла в оригинальном описании указывается как 43 мм. Значительные различия заметны в ширине крыльев, если заднее крыло с расширенной анальной областью составляет 22 мм, то наибольшая ширина переднего достигает лишь 16.7 мм. На заднем и переднем крыльях сходный характер жилкования в системах RS и M. В отличие от простой CuP на переднем крыле, на заднем крыле эта жилка трехветвистая, что не удивительно при более широкой анальной части крыла.
Материал. Помимо типовых экземпляров, из Республики Коми: ПИН, № 1631/391 – прямой и обратный отпечатки переднего имагинального крыла; Воркутское месторождение, р. Хей-Яга, обн. 3, сл. 9; нижняя пермь, лекворкутская свита, аячьягинская подсвита; экз. ПИН, №№ 1631/392, 393, 394 – фрагменты заднего имагинального крыла, там же, обн. 3, сл. 14 (392); экз. ПИН, № 1631/395 – прямой отпечаток заднего нимфального крыла, там же; экз. ПИН, № 1631/396 – основание переднего имагинального крыла, Хальмеръюское месторождение, скв. ХК-780, гл. 298.0 м; нижняя пермь, интинская свита.
Список литературы
Лозовский В.Р., Миних М.Г., Грунт Т.А. и др. Уфимский ярус Восточно-Европейской шкалы: статус, валидность, корреляционный потенциал // Стратигр. Геол. корреляция. 2009. Т. 17. № 6. С. 46–58.
Македонов А.В., Погоревич В.В. Северная часть Предуральского прогиба и хребет Пай-Хой // Стратиграфия СССР. Пермская система. М.: Недра, 1966. С. 244–259.
Муравьев Н.С. Воркутский ярус пермской системы // Пермская система: вопросы стратиграфии и развития органического мира. Казань: Изд-во Казанск. ун-та, 1988. С. 88–107.
Пухонто С.К. Стратиграфия и флористическая характеристика пермских отложений угольных месторождений Печорского бассейна. М.: Научн. мир, 1998. 312 с.
Расницын А. П., Сукачева И. Д., Аристов Д. С. Пермские насекомые воркутской серии Печорского бассейна и их стратиграфическое значение // Палеонтол. журн. 2005. № 4. С. 63–75.
Синиченкова Н.Д. Отряд Dictyoneurida // Историческое развитие класса насекомых. М.: Наука, 1980. С. 44–47 (Тр. Палеонтол. ин-та АН СССР. Т. 175).
Синиченкова Н.Д. Новые виды мисхоптеридовых из пермских отложений Воркутского бассейна (Insecta: Mischopterida = Megasecoptera) // Палеонтол. журн. 2019. № 1. С. 53–55.
Шаров А.Г., Синиченкова Н.Д. Новые Palaeodictyoptera с территории СССР // Палеонтол. журн. 1977. № 1. С. 48–63.
Carpenter F.M. Treatise on Invertebrate Palaeontology. Pt R. Arthropoda 4. Vol. 3. Superclass Hexapoda. Lawrence, Boulder: Geol. Soc. America, Univ. Kansas Press, 1992. 655 p.
Carpenter F.M., Richardson E. Jr. Additional insects in Pennsylvanian concretions from Illinois // Psyche. 1971. V. 78. P. 267–295.
Kukalova J. Revisional study of the order Palaeodictyoptera in the Upper Carboniferous shales of Commentry, France. Pt 1 // Psyche. 1969. V. 76. P. 163–215.
Prokop J., Nel A., Engel M. S. et al. New Carboniferous fossils of Spilapteridae enlighten postembryonic wing development in Palaeodictyoptera // Syst. Entomol. 2016. V. 41. P. 178–190.
Prokop J., Pecharová M., Jarzembowski E. A., Ross A. J. New palaeodictyopterans from the Late Carboniferous of the UK (Insecta: Palaeodictyoptera) // Earth and Environ. Sci. Trans. Roy. Soc. Edinburgh. 2017. V. 107. P. 99–107.
Sinitshenkova N.D. Superorder Dictyoneuridea Handlirsch, 1906 // History of Insects / Eds. Rasnitsyn A.P., Quicke D.L.J. Dordrecht, Boston, L.: Kluwer Acad. Publ., 2002. P. 115–124.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал