

Палеонтологический журнал, 2020, № 3, стр. 20-31
Раннемеловые устрицы Мангышлака: таксономический состав, стратиграфическое и палеобиогеографическое распространение
И. Н. Косенко a, b, *, Е. К. Метелкин b, c
a Нанкинский институт геологии и палеонтологии Китайской академии наук
Нанкин, Китай
b Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
c Новосибирский государственный университет
Новосибирск, Россия
* E-mail: ki-2009@mail.ru
Поступила в редакцию 22.05.2019
После доработки 18.07.2019
Принята к публикации 22.07.2019
Аннотация
На основе изучения представительной коллекции берриас-барремских устриц Мангышлака уточнены сведения об их таксономическом составе, стратиграфическом и палеобиогеографическом распространении. Описан древнейший вид рода Gyrostrea – G. bogdanovae sp. nov. из кугусемской свиты (готерив–баррем), а также Aetostreon subsinuatum (Leymerie, 1842), впервые установленный на Мангышлаке и представленный здесь тремя морфотипами (типовым, falciformis и dorsatum).
ВВЕДЕНИЕ
Раннемеловые устрицы Мангышлака представляют собой интерес по двум причинам. Во-первых, раннемеловая фауна этого региона носит смешанный характер, сочетая в себе тетические и бореальные элементы. Это связано с палеогеографическим положением территории полуострова в раннем мелу, представлявшей из себя пролив, соединяющий северные акватории Тетиса с Русским морем (Барабошкин, 2003, 2007). В связи с этим, важно понять, какими таксонами по происхождению (тетическими или бореальными) представлены сообщества устриц. Во-вторых, на границе юры и мела и в самом начале мела происходит значительная таксономическая перестройка в сообществах устриц: практически повсеместно исчезают грифеи (за исключением арктических морей Сибири (Захаров, 1966; Kosenko, 2017), в самом начале берриаса появляются первые представители Pycnodonteinae (Kosenko, 2018), а среди Exogyrinae широкое распространение получают роды Aetostreon и Ceratostreon вместо доминировавших в поздней юре родов Nanogyra и Palaeogyra. Появление и распространение типичных для мела родов устриц в разных регионах происходило, по-видимому, не одновременно, и только к валанжину меловые таксоны получили широкое распространение. Детальная привязка представленных в коллекции устриц позволяет реконструировать, как менялся таксономический состав остреоидей на территории Мангышлака в раннем мелу, и в свою очередь уточнить особенности стратиграфического и палеобиогеографического распространения этой группы двустворчатых моллюсков.
Раннемеловые устрицы Мангышлака изучались В.П. Семеновым (1896, 1899), определения устриц присутствуют в статье Т.А. Мордвилко (1953), их описания даны в работах Т.Н. Богдановой (1978, 1980; Валанжин…, 1983; Берриас…, 1988). Из берриаса были описаны Deltoideum delta (Smith, 1817) (лона Neocosmoceras и Septaliphoria semenovi), Liostrea germaini mangyshlakensis (Bogdanova, 1988), Rhynchostreon? tombeckianum (d’Orbigny, 1848), Rastellum rectangularis (Roemer, 1839) (весь берриас). Из валанжина были описаны Ceratostreon minos (Coquand, 1869) (берриас-валанжин) и Rastellum ex gr. macroptera (J. de C. Sowerby, 1825) (лона Buchia keyserlingi). Из берриаса и валанжина Богдановой были описаны виды Picnodonte (=Pycnodonte) miranda Bogdanova, 1980, а из валанжина – Ceratostreon fragilis Bogdanova, 1980. Вместе с тем, по мере накопления во ВСЕГЕИ (Санкт-Петербург) новых коллекций становилось понятно, что таксономический состав раннемеловых устриц Мангышлака и их стратиграфическое распространение нуждаются в уточнении (Т.Н. Богданова, устн. сообщ.). В статье представлены результаты ревизии таксономического состава и стратиграфического распространения устриц в нижнем мелу Мангышлака; монографически описаны виды, ранее с его территории неизвестные.
МАТЕРИАЛ
Исследована представительная коллекция раннемеловых (берриас-барремских) устриц Мангышлака, переданная авторам из ВСЕГЕИ и состоящая из сборов Н.П. Луппова, Т.А. Мордвилко, В.Г. Кликушина, Т.Н. Богдановой, проводивших палеонтолого-стратиграфические исследования на этом полуострове. Коллекция происходит из 32 обнажений (рис. 1), насчитывает более 200 раковин, большинство из которых характеризуются очень хорошей сохранностью и точной стратиграфической привязкой: для берриаса и валанжина – к аммонитовым и бухиевым лонам, для готерива и баррема – к ярусу. Кроме того, авторами изучена коллекция берриас-валанжинских устриц Мангышлака, хранящаяся в Центральном научно-исследовательском геологоразведочном музее им. акад. Ф.Н. Чернышева (ЦНИГР музее). Изученная коллекция устриц передана на хранение в центр коллективного пользования “Геохрон” ИНГГ СО РАН (колл. № 2087).
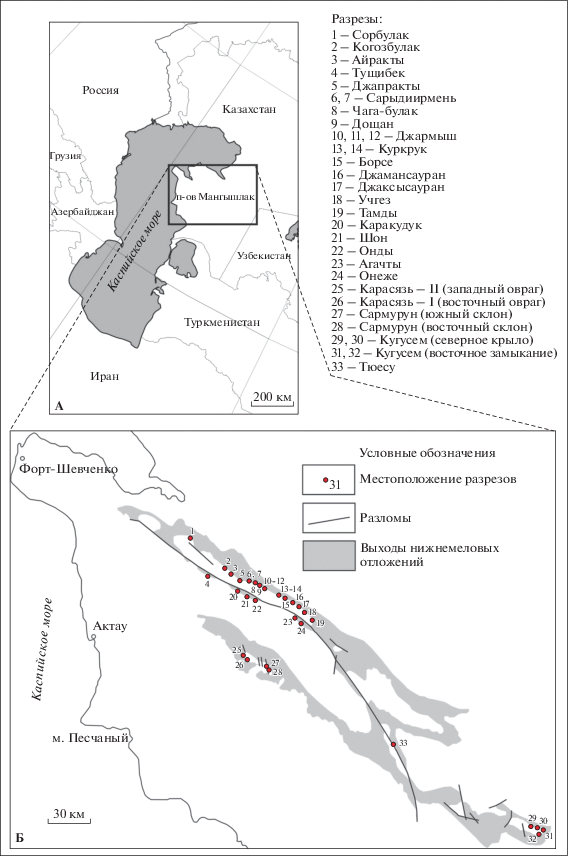
СВЕДЕНИЯ ПО СТРАТИГРАФИИ НИЖНЕГО МЕЛА МАНГЫШЛАКА
Полуостров Мангышлак расположен на восточном побережье Каспийского моря в Казахстане (рис. 1). На территории Мангышлака широко развиты нижнемеловые отложения (Стратиграфия…, 1986).
Ниже приводится краткий обзор сведений по стратиграфии нижнего мела (берриаса-баррема) Мангышлака, выполненный по материалам из нескольких ключевых работ (Валанжин…, 1983; Стратиграфия…, 1986; Берриас…, 1988).
Берриас–нижний готерив Мангышлака представлены разнообразными по литологическому составу мелководно-морскими прибрежными маломощными отложениями, фациально изменчивыми по площади. Верхний готерив и баррем представлены преимущественно континентальной кугусемской свитой, сложенной пестроцветными глинами с преобладанием красноватых и зеленоватых разностей с прослоями песчаников (Савельев, Василенко, 1963; Стратиграфия…, 1986). Стратиграфия берриаса и валанжина Мангышлака к настоящему времени детально разработана (Валанжин…, 1983; Берриас…, 1988). Берриасские отложения залегают со стратиграфическим несогласием на средне- и верхнеюрских, при этом нижний берриас, как и верхи берриаса, на Мангышлаке отсутствуют. Валанжин также залегает на берриасе со стратиграфическим несогласием. Берриасские и валанжинские отложения на Мангышлаке подразделены на местные стратиграфические подразделения – лоны (Валанжин…, 1983; Берриас…, 1988), выделенные по аммонитам и бухиям. Берриас подразделен на три лоны: Neocosmoceras и Septaliphoria semenovi, Buchia volgensis, и Riasanites и Pygurus rostratus (Берриас…, 1988). Валанжин также подразделен на три лоны: Buchia keyserlingi, Polyptichites spp. и Dichotomites sp. (Валанжин…, 1983). Готеривские морские отложения представлены на Мангышлаке небольшой пачкой (до 10 м) изменчивого на площади вещественного состава, характерной особенностью которой являются ракушняки с крупными двустворками Isognomon sp., устрицами, кораллами и брахиоподами. Эта пачка не охарактеризована ископаемыми, типичными только для готерива, поэтому граница между валанжином и готеривом проводится условно по смене комплексов ископаемых. Повсюду эта пачка согласно перекрывается кугусемской свитой. Схема соотношения стратиграфических подразделений берриаса – баррема Мангышлака показана на рис. 2.
Рис. 2.
Стратиграфические подразделения берриаса–баррема Мангышлака (по: Стратиграфия СССР, 1986; с изменениями).
В палеогеографическом отношении территория современного Мангышлака представляла часть Каспийского пролива, соединявшего Русское море с акваториями Тетис, и существовавшего на протяжении почти всего раннего мела, кроме раннего берриаса. Морские условия в проливе прекращались в позднем валанжине–раннем готериве, в конце готерива–барреме и начале апта, когда он заполнялся континентальными и лагунными осадками (Барабошкин, 2003, 2007).
ОБСУЖДЕНИЕ
В результате проведенного исследования авторам удалось уточнить таксономический состав и стратиграфическое распространение устриц в берриасе–барреме Мангышлака (табл. 1).
Таблица 1.
Распространение устриц в берриасе–барреме Мангышлака. Местные стратиграфические подразделения даны по Стратиграфия…, 1986
| Ярус | Местное стратиграфическое подразделение | Таксономический состав устриц |
|---|---|---|
| Баррем | Кугусемская свита | Aetostreon subsinuatum, Gyrostrea bogdanovae sp. nov. |
| Готерив | ||
| Ракушняк с Isognomon | “Liostrea” germaini, Ceratostreon minos, A. subsinuatum, A. subsinuatum морфотип dorsatum | |
| Валанжин | Лона Dichotomites sp. | “Pycnodonte” miranda*1, Amphidonte? sp., A. subsinuatum, C. minos |
| Лона Polyptichites spp. | “P.” miranda*, C. minos*, A. subsinuatum морфотип falciformis, A. subsinuatum | |
| Лона B. keyserlingi | “P.” miranda*, Amphidonte? sp., Rastellum (Rastellum) ex gr. macroptera*, Ceratostreon fragilis*, C. minos*, A. subsinuatum, A. subsinuatum морфотип falciformis | |
| Берриас | Лона Riasanites и P. rostratus | Rastellum (Arctostrea) rectangularis*, Rhynchostreon? tombeckianum*, “L.” germaini, C. minos |
| Лона B. volgensis | R. (A.) rectangularis*, Rh.? tombeckianum*, “L.” germaini, C. minos | |
| Лона Neocosmoceras и S. semenovi | Deltoideum delta*, R. (A.) rectangularis*, Rh.? tombeckianum*, “P.” miranda*, “L.” germaini, Amphidonte? sp., C. minos* |
Среди полученных новых данных следует отметить следующее:
1. В валанжин-барремском интервале Мангышлака широко представлены устрицы рода Aetostreon. Ранее его представители не описывались из Мангышлака, а в литературе (Валанжин…, 1983, Стратиграфия…, 1986) встречались лишь указания о присутствии A. subsinuatum (Leymerie, 1842) в валанжине и готериве.
2. Из кугусемской свиты (верхний готерив–баррем) описан новый вид устриц Gyrostrea bogdanovae sp. nov., являющийся к настоящему времени древнейшим представителем рода. Это позволило предложить новую гипотезу возникновения гирострей.
ОПИСАНИЕ ТАКСОНОВ
НАДСЕМЕЙСТВО OSTREOIDEA RAFINESQUE, 1815
СЕМЕЙСТВО FLEMINGOSTREIDAE STENZEL, 1971
ПОДСЕМЕЙСТВО CRASSOSTREINAE SCARLATO ET STAROBOGATOV, 1978
Род Gyrostrea Mirkamalov, 1963
Gyrostrea bogdanovae Kosenko et Metelkin, sp. nov.
Название вида в честь палеонтолога Т.Н. Богдановой, специалиста по стратиграфии мела и меловым двустворчатым моллюскам и аммонитам ZooBank LSID: urn:lsid:zoobank.org:act: 897951AF-D9B3-44FA-B365-A7D67D162D42.
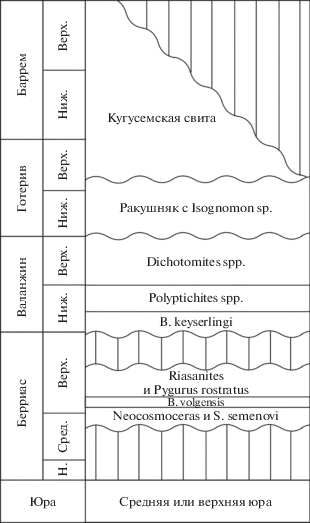
Голотип – ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/4 (рис. 3, д, ж); Казахстан, п-ов Мангышлак, Джаман-Сауран; верхний готерив–баррем, кугусемская свита.
Рис. 3.
Gyrostrea bogdanovae sp. nov.: а – экз. ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/1, левая створка, вид на внутреннюю поверхность; б – экз. ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/2, левая створка вид на внутреннюю поверхность; в, г – экз. ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/3, сросток из двух раковин: в – правая створка, вид на внутреннюю поверхность, г – левая створка, приросшая к правой, вид на внутреннюю поверхность; д, ж – голотип ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/4, левая створка с приросшими к ней ювенильными устрицами: д – вид на внешнюю поверхность; ж – вид на внутреннюю поверхность с приросшими к ней раковинами того же вида; е – экз. ЦКП “Геохрон” (ИНГГ СО РАН) № 2087/5, левая створка, приросшая к другой створке того же вида, вид на внутреннюю поверхность. Все экземпляры происходят из кугусемской свиты местонахождения Джаман-Сауран. Длина масштабных отрезков – 10 мм.
Описание (рис. 3). Раковины от небольших до средних размеров, тонкостенные, слабо неравностворчатые, вытянутые в высоту, прямоугольно-овальных очертаний. Левая створка на ранних стадиях развития сильно выпуклая, с возрастом выполаживается, снаружи покрыта концентрическими чешуями роста, редко с нерегулярными радиальными ребрами и радиальными складками. Замочная площадка гиростреоидного типа: на ранних стадиях развития спирально закрученная, позднее – прямая. Ямка для связки глубокая, четко выраженная. Валики хорошо выраженные, развиты одинаково. Макушка спирально завернута назад в плоскости створки, не выступающая. Примакушечная полость хорошо выраженная, очень глубокая. Отпечаток мускула-замыкателя почковидной формы, выражен слабо, сильно смещен к заднему краю, чуть смещен к нижнему краю. Внутренняя полость створок гладкая, хоматы отсутствуют. Правые створки в коллекции отсутствуют. На сколах раковины вблизи замочной площадки заметны многочисленные пустые камеры.
Размеры в мм и отношения:
| № экз. | В | Д | В/Д | |
|---|---|---|---|---|
| Голотип | 2087/4 | 43.1 | 36.7 | 1.17 |
| 2087/2 | 32.6 | 33.5 | 0.97 | |
| 2087/5 | 19.7 | 15.6 | 1.26 | |
| 2087/6 | 24.0 | 26.0 | 0.92 | |
| 2087/7 | 34.2 | 29.2 | 1.17 |
Сравнение. От G. turkestanensis (Bobkova, 1949), G. akrabatensis Mirkamalov, 1966, G. ventricosa Mirkamalov, 1966, G. longa (Bobkova, 1961), описываемый вид отличается, в первую очередь, меньшими размерами раковин. Наибольшая высота раковины G. bogdanovae sp. nov. составляет чуть более 4 см, в то время как остальные виды характеризуются раковинами с высотой до 8–10 см. Другой характерной особенностью G. bogdanovae sp. nov. является то, что большинство экземпляров образует сростки из нескольких раковин, что не характерно для других гирострей.
Материал. 30 отдельных левых створок, включая ювенильные створки из сростков, из кугусемской свиты, местонахождение Джаман-Сауран.
СЕМЕЙСТВО GRYPHAEIDAE VIALOV, 1936
ПОДСЕМЕЙСТВО EXOGYRINAE VIALOV, 1936
Род Aetostreon Bayle, 1878
Aetostreon subsinuatum (Leymerie, 1842)
Табл. IV , фиг. 1–3; табл. V , фиг. 1–3 (см. вклейку)
Exogyra subsinuata: Leymerie, 1842, c. 17, табл. XII , фиг. 3–7 ; Эристави, 1957, с. 44; Прозоровский и др., 1961, с. 124, табл. IX , фиг. 3.
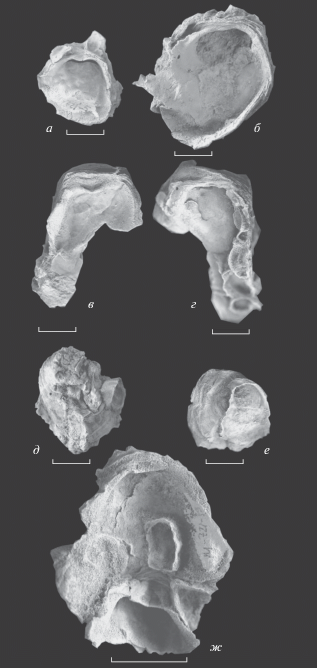
Рис. 4.
Поля корреляции измеренных признаков раковин Aetostreon subsinuatum (Leymerie): длины и высоты (а), длины и выпуклости (б) и отношений длины к высоте (Д/В) и выпуклости к высоте (Вып/В). Измерения произведены на 33 левых створках. Точки соответствуют типовому морфотипу, плюсы – морфотипу falciformis.
Exogyra subsinuata var. falciformis: Leymerie, 1842, с. 16, 17, табл. XII , фиг. 5 ; Ренгартен, 1926, с. 62, табл. IV , фиг. 5 , табл. VI , фиг. 1; Атлас…, 1949, с. 158, табл. XXVI , фиг. 5 , табл. XXVII , фиг. 1 a, b, c.
Ostrea (Exogyra) couloni: Семенов, 1899, с. 71.
Exogyra couloni: Wollemann, 1900, c. 8, табл. I , фиг. 1.
Exogyra subsinuata forma typica: Ренгартен, 1926, с. 61, табл. IV , фиг. 4, табл. V , фиг. 1.
Exogyra subsinuata var. falciformis: Ренгартен, 1926, с. 62, табл. IV , фиг. 5 , табл. VI , фиг. 1.
Exogyra subsinuata var. crimica: Эристави, 1957, с. 44, табл. II , фиг. 1.
Exogyra aff. subsinuata: Эристави, 1957, с. 44.
Exogyra subsinuata aff. var. falciformis: Эристави, 1957, с. 44.
Amphidonta subsinuata: Атлас…, 1960, с. 199, табл. XVI , фиг. 1–4; Атлас…, 2005, с. 257, табл. XLI , фиг. 5 .
Exogyra sp. nov. inden.: Богданова, 1961, с. 138, табл. III , фиг. 1–4.
Exogyra sp.: Богданова, 1961, с. 140, табл. II , фиг. 5 .
Exogyra falciformis: Прозоровский и др., 1961, с. 125, табл. X , фиг. 1, табл. XI , фиг. 1.
Aetostreon latissimum: Pugaczewska, 1975, с. 51, табл.VII–IX; Dhondt, Dieni, 1988, с. 38, табл. VIII , фиг. 1–7 , табл. IX , фиг. 1–6 , рис. 18 .
Aetostreon subsinuatum: Богданова в Берриас…, 2012, с. 250, табл. XLIV , фиг. 13 .
Лектотип (выбран авторами) – экз. Exogyra subsinuata, изображенный в работе М. Леймери (Leimerie, 1842, табл. XII , фиг. 3, левая створка); Франция, департамент Верхняя Марна; нижний мел, готерив.
Описание. Раковины средних и крупных размеров (высотой до 108 мм и длиной до 80 мм), толстостенные, сильно неравностворчатые, вытянутые в высоту, от треугольных, треугольно-овальных, овальных до серповидных очертаний. Левая створка обычно сильно, реже умеренно выпуклая, с невыступающей макушкой, прижатой к заднему краю, и килем, делящим створку на переднее и заднее поля. Площадка прирастания различного размера – от полного отсутствия до очень большого размера, иногда она занимает практически всю закилевую часть створки. Киль обычно хорошо выраженный, реже сглаженный. Некоторые левые створки имеют выраженный радиальный сулькус (параллельный килю приближенный к заднему краю створки желобок). Наружная поверхность створки покрыта концентрическими грубыми чешуями роста. Иногда наблюдается слабо выраженная радиальная ребристость. Замочная площадка экзогироидного типа, но передний валик не всегда редуцирован полностью. Примакушечная полость отсутствует. Правая створка плоская, редко слабо вогнутая, покрыта грубыми концентрическими пластинами роста. Внутренняя поверхность створки гладкая. Замочная площадка аркообразно изогнута. Задний валик не всегда редуцирован, иногда не до конца редуцирован и передний валик. Зубовидный выступ отсутствует. Отпечаток мускула-замыкателя большого размера, овальной или округлой формы, смещен к заднему краю, слегка приближен к верхнему краю створки. Хоматы отсутствуют.
Размерыв мм и отношения. Были измерены высота (В), длина (Д) и выпуклость (Вып) 33 левых створок из разных местонахождений и разных стратиграфических уровней, а затем посчитаны коэффициенты удлинения (Д/В) и выпуклости (Вып/В). Результаты измерений были обработаны с помощью программы Past 3, где были построены поля корреляции высоты и длины (рис. 4, а), выпуклости и высоты (рис. 4, б) и коэффициентов выпуклости и удлинения (рис. 4, в). На полях корреляции хорошо видно, что значения измеренных величин и коэффициентов морфотипа falciformis (обозначен знаком “+”) не выбиваются из значений типового морфотипа, что свидетельствует в пользу рассмотрения морфотипа falciformis именно в качестве морфотипа, а не самостоятельного вида.
Изменчивость. При первоописании вида Леймери выделил несколько вариететов (Leymerie, 1842, табл. XII , фиг. 3–7 ). В качестве типового вариетета Леймери рассматривал относительно изометричные раковины субтреугольных очертаний. Вариететы falciformis и aquilina характеризуются вытянутыми в высоту раковинами, falcifomis – с резко выраженным килем и наличием радиального сулькуса, aquilina – с сильновыпуклой левой створкой без резко выраженного киля. Вариетет dorsata обладает изометричной раковиной субтреугольных очертаний с выраженными радиальными складками.
Нами встречены раковины, соответствующие трем вариететам, выделенным Леймери: типовому, falciformis и dorsata. Наиболее многочисленны раковины типового вариетета. Вместе с ними в валанжине присутствуют раковины вариетета falciformis, а в готериве – единственная раковина вариетета dorsata. Вариетет dorsata, по-видимому, редко встречается, о чем свидетельствует отсутствие его упоминаний в литературе. Вариетет falciformis, напротив, встречается относительно часто, и был неоднократно описан различными авторами либо как отдельный вид, либо под другими видовыми названиями.
В настоящей статье вариететы, выделенные Леймери, рассматриваются как морфотипы вида A. subsinuatum. В пользу этого свидетельствует совместная встречаемость раковин, относимых к разным вариететам, наличие промежуточных по морфологии раковин в изученной коллекции, а также совместная встречаемость нескольких “вариететов”-морфотипов, отмеченная в других регионах (Pugaczewska, 1975; Dhondt, Dieni, 1988).
Сравнение. Вытянутые в высоту раковины морфотипа falciformis имеют сходство с видом A. imbricatum (Krauss, 1843) из валанжина Южной Африки. Этот вид детально описан М. Купером (Cooper, 1995, с. 5, фиг. 1, 5–7, 8 A–C ). A. subsinuatum морфотип falciformis отличается от A. imbricatum более резким килем и маленькой не грифоидной макушкой.
От вида A. pilmatuegrossum (Rubilar et Lazo, 2009) (с. 730, фиг. 3-14 ) из валанжина Аргентины отличается строением киля, который у аргентинского вида имеет сложное строение и делится на первичный и вторичный.
Некоторые авторы (Pugaczewska, 1975; Dhondt, Dieni, 1988) включают вид A. subsinuatum в синонимику широко понимаемого вида A. latissimum (Lamarck, 1801), являющегося типовым видом рода Aetostreon (лектотип этого вида происходит из апта Франции, хранится в Музее естественной истории Парижа: http://coldb.mnhn.fr/catalognumber/mnhn/f/a50176). Однако типовые экземпляры двух этих видов существенно отличаются очертаниями и степенью выпуклости раковины (субтреугольная, более узкая и выпуклая раковина у A. subsinuatum и субовальная, широкая раковина у A. latissimum) и степенью развития киля (у A. subsinuatum киль сильнее выражен за счет того, что переднее и заднее поля створки, разделенные килем, круче падают по сравнению с A. latissimum), и происходят из разных ярусов мела. Наблюдения авторов над аптскими устрицами A. latissimum из Туркменистана показали, что виды A. subsinuatum и A. latissimum имеют заметные отличия в морфологии раковин, позволяющие рассматривать их в качестве разных видов. Эти отличия заключаются в: 1) меньших размерах A. subsinuatum; 2) более выпуклой форме раковин A. subsinuatum; 3) более выраженном киле у A. subsinuatum, делящим переднее и заднее боковые поля под меньшим углом, чем в случае A. latissimum; 4) округлом отпечатке мускула-замыкателя у A. subsinuatum (у A. latissimum отпечаток мускула-замыкателя вдавленный, почковидной формы, с вогнутым верхним краем); 5) более архаичном строении замочной площадки у A. subsinuatum (прогрессивное строение замочной площадки у A. latissimum выражено в полной редукции заднего бугорка, а также наличии углубления под задним бугорком на левых створках). Перечисленные отличия и разные диапазоны стратиграфического распространения позволяют различать A. subsinuatum и A. latissimum, а также предположить происхождение A. latissimum от A. subsinuatum.
Замечания. Морфологически близки к A. subsinuatum экзогиры, описанные из неокома Мексики Р. Имлеем (Imlay, 1937), которые по морфологии раковин должны рассматриваться в составе рода Aetostreon: A. reedi (Imlay, 1937) (там же, с. 566, табл. 77 , фиг. 1–6 , табл. 78 , фиг. 1–7 ), A. putnami (Imlay, 1937) (там же, с. 566, табл. 81 , фиг. 1–8 ) и A. tamaulipana (Imlay, 1937) (там же, с. 567, табл. 79 , фиг. 1–5 ). Они встречаются совместно, начиная с основания мела, а различия между ними напоминают различия между морфотипами вида A. subsinuatum и заключаются в степени выраженности киля, а также в форме и очертаниях раковины. Вид A. reedi соответствует морфотипу falciformis, вид A. putnami – морфотипу aquilina, а вид A. tamaulipana – типовому морфотипу вида A. subsinuatum, хотя и обладает менее выраженным килем. Это позволяет предположить, что описанные Имлеем виды являются младшими синонимами вида A. subsinuatum. Тем не менее, доказательство этой точки зрения требует изучения коллекций из нижнего мела Мексики.
Материал. Типовой морфотип: четыре левые створки из местонахождения Сорбулак, лоны B. keyserlingi; четыре левые створки из лоны B. keyserlingi Шаира; одна левая и одна правая створка из лоны B. keyserlingi Кугусема; две левые створки из лоны Dichotomites spp. Сорбулака; одна левая створка из валанжина – готерива Карашимрау; три левых створки из готерива Чага-Булака; две левых створки и одна правая створка из готерива Шаира; одна целая раковина, две левых створки и четыре правых створки из готерива Когоз-Булака; три левых створки из кугусемской свиты Чага-Булака; одна левая и две правых створки из кугусемской свиты Таспаса; четыре левых створки и пять правых створок из кугусемской свиты Агачты. Морфотип falciformis: пять левых створок и одна целая раковина из лон B. keyserlingi и Polyptichites spp. валанжина Мангышлака (Карашимрау, Сорбулак). Морфотип dorsatum: единственная левая створка из готерива Мангышлака.
Распространение. Берриас–готерив Крыма; валанжин–готерив Мангышлака, Франции, Германии, Польши, Кавказа, Средней Азии; (?) нижний мел Мексики.
ПОЛОЖЕНИЕ GYROSTREA В СИСТЕМЕ OSTREOIDEA
Род Gyrostrea был выделен Х.Х. Миркамаловым (1963) в составе подсемейства Exogyrinae и рассматривался вместе с родом Pseudogyra как регрессивная ветвь экзогирин (Миркамалов, 1966). Позднее С. Френе (Freneix, 1982) на основании изучения морфологии и микроструктуры раковины Gyrostrea delettrei (Coquand, 1862) отнесла Gyrostrea к подсемейству Crassostreinae. Н. Мальхус (Malchus, 1990) рассматривал Gyrostrea в составе трибы Flemingostreini подсемейства Liostreinae. По нашему мнению, главными признаками для определения положения Gyrostrea в системе Ostreoidea являются: наличие хорошо выраженной примакушечной полости, отпечаток мускула-замыкателя почковидной формы, смещенный к задне-нижнему краю, а также способность к образованию сростков из нескольких раковин. Все эти признаки характерны для подсемейства Crassostreinae. Наличие радиальных складок у некоторых представителей Gyrostrea также сближает их с родом Crassostrea. Кроме того, спиральное закручивание замочной площадки на ранних стадиях онтогенеза иногда наблюдается у современных устриц Crassostrea gigas (Thunberg, 1793) (Косенко, 2016). Все вышеперечисленные признаки рода Gyrostrea свидетельствуют о необходимости рассматривать его в составе подсемейства Crassostreinae семейства Flemingostreidae.
ПРОИСХОЖДЕНИЕ РОДА GYROSTREA
Предположение о происхождении рода Gyrostrea от представителей рода Curvostrea в апте–альбе было высказано Мальхусом (Malchus, 1996). Находка Gyrostrea bogdanovae sp. nov. в кугусемской свите (верхний готерив–баррем) Мангышлака позволяет выдвинуть другую гипотезу. Среди широко распространенных берриас-готеривских устриц морфологически близок к роду Gyrostrea вид “Liostrea” germaini (Coquand, 1869). Эти устрицы присутствуют и в берриасе–готериве Мангышлака. Вид характеризуется неравностворчатыми раковинами с выпуклыми левыми створками, вытянутыми в высоту замочными площадками от субтреугольной до прямоугольной формы с выраженной ямкой для связки, наличием примакушечной полости, почковидным отпечатком мускула-замыкателя, смещенным к задне-нижнему краю, способностью к образованию сростков из нескольких раковин, а также плохо выраженной нерегулярной радиальной скульптурой. Кроме того, некоторые изученные нами экземпляры имеют спирально завернутую на ранних стадиях замочную площадку. Все эти признаки сближают устриц “L.” germaini с родом Gyrostrea и позволяют рассматривать их в качестве предкового таксона. Таксономическое положение вида “L.” germaini на наш взгляд дискусионно. Наличие у этого вида нерегулярных радиальных ребер не позволяет однозначно относить его к роду Liostrea, характеризующегося гладкой поверхностью левых створок. В то же время, не вызывает сомнений принадлежность “L.” germaini к Crassostreinae. Уточнение систематического положения этого вида требует отдельного исследования и выходит за рамки этой статьи.
ПАЛЕОБИОГЕОГРАФИЧЕСКИЙ АНАЛИЗ КОМПЛЕКСОВ УСТРИЦ МАНГЫШЛАКА
Уточненные данные по таксономическому составу и стратиграфическому распространению устриц Мангышлака позволяют сравнить комплексы устриц этого района с близлежащими регионами.
Данные о берриас-валанжинских устрицах Русской плиты крайне немногочисленны. Нами обнаружено лишь описание вида Ostrea limaciforme Gerasimov, 1955 из берриаса Московской и Рязанской областей (здесь этот вид рассматривается в составе рода Praeexogyra Charles et Maubeuge, 1952). С другой стороны, имеются обобщающие сведения по берриас–валанжинским устрицам Крыма (Берриас…, 2012), Польши (Pugaczewska, 1975), Урала и Сибири (Захаров, 1963, 1966; Захаров, Месежников, 1974; Kosenko, 2017). Эти данные позволяют сравнить таксономический состав устриц Мангышлака и приведенных выше регионов (табл. 2, 3).
Таблица 2.
Распространение устриц в берриасе Мангышлака и сопредельных регионов (по данным: Герасимов, 1955; Захаров, 1963, 1966; Захаров, Месежников, 1974; Pugaczewska, 1975; Берриас…, 2012; Kosenko, 2017)
| Таксон | Регион | |||||
|---|---|---|---|---|---|---|
| Мангышлак | Крым | Польша | Русская платформа | Урал | Сибирь | |
| Deltoideum delta | ˅ | |||||
| Rastellum rectangularis | ˅ | ˅ | ||||
| Rhynchostreon? tombeckianum | ˅ | ˅ | ||||
| Rhynchostreon? etalloni | ˅ | |||||
| “Pycnodonte” miranda | ˅ | |||||
| Pycnogryphaea weberae | ˅ | |||||
| “Liostrea” germaini | ˅ | ˅ | ||||
| Ceratostreon minos | ˅ | ˅ | ||||
| C. tuberculiferum | ˅ | ˅ | ||||
| Aetostreon subsinuatum | ˅ | |||||
| “Ostrea” sanctae crucis | ˅ | |||||
| Praeexogyra limaciforme | ˅ | |||||
| P. lyapinensis | ˅ | |||||
| P. siberica | ˅ | |||||
| Pernostrea uralensis | ˅ | |||||
| Deltoideum (Boreiodeltoideum) anabarensis | ˅ | |||||
Таблица 3.
Распространение устриц в валанжине Мангышлака и сопредельных регионов (по данным: Захаров, 1963, 1966; Pugaczewska, 1975; Берриас…, 2012; Kosenko, 2017)
| Таксон | Регион | |||
|---|---|---|---|---|
| Мангышлак | Крым | Польша | Сибирь | |
| Rastellum rectangularis | ˅ | |||
| Rastellum ex gr. macroptera | ˅ | |||
| Rhynchostreon? tombeckianum | ˅ | |||
| “Pycnodonte” miranda | ˅ | |||
| “Liostrea” germaini | ˅ | ˅ | ||
| Ceratostreon minos | ˅ | ˅ | ˅ | |
| Aetostreon subsinuatum | ˅ | ˅ | ˅ | |
| “Ostrea” sanctae crucis | ˅ | |||
| Gryphaeostrea cf. lateralis | ˅ | |||
| Gryphaeostrea? arduennensis | ˅ | |||
| “Lopha” cf. cotteaui | ˅ | |||
| “Lopha” cf. eos | ˅ | |||
| Pernostrea cucurbita | ˅ | |||
| Deltoideum (Boreiodeltoideum) anabarensis | ˅ | |||
| Gryphaea borealis | ˅ | |||
Сравнение таксономического состава берриасских устриц Мангышлака и сопредельных регионов показывает, что наибольшее сходство наблюдается между Мангышлаком и Крымом, присутствуют четыре общих вида: R. (A.) rectangularis, Rh.? tombeckianum, “L.” germaini, C. minos, широко распространенных в Тетической надобласти. Стоит отметить присутствие в берриасе Мангышлака вида D. delta. Этот вид типичен для верхней юры суббореальных бассейнов (оксфорд-кимеридж Англии, севера Франции и Нормандии, волжский ярус Русской платформы), экотонного бореально-тетического Польского бассейна (кимериджский–волжский ярусы), а присутствие D. delta в берриасе Мангышлака является наиболее поздней находкой вида. Стоит также отметить присутствие A. subsinuatum в берриасе Крыма, как наиболее раннее точно задокументированное появление вида, получившего в валанжине широкое распространение. Кроме того, обращает на себя внимание присутствие представителей Pycnodonteinae в Крыму и на Мангышлаке. В Крыму подсемейство представлено многочисленными находками вида Pycnogryphaea weberae, а на Мангышлаке – видом “Pycnodonte” miranda, точная родовая принадлежность которого на данный момент не определена, но принадлежность к Pycnodonteinae не вызывает сомнений. Находки пикнодонтин в берриасе Мангышлака и Крыма являются к настоящему времени наиболее ранними находками представителей этого подсемейства.
Валанжинский комплекс устриц Мангышлака отличается от берриасского, главным образом, появлением в нем рода Aetostreon. В валанжине сходство наблюдается между Мангышлаком, Крымом и Польшей. Отмечаются общие виды: C. minos, A. subsinuatum. В Крыму и на Мангышлаке также присутствуют виды рода Rastellum – в Крыму продолжает существование R. (A.) rectangularis, на Мангышлаке этот вид замещается близким видом R. (R.) ex gr. macroptera.
Анализ распространения устриц показывает, что устричная фауна Мангышлака в берриасе и валанжине являлась типично тетической. Бореальное влияние эта фауна на себе не испытывала. Ее основу составляют широко распространенные от Польши до Северной Африки раннемеловые виды – “L.” germaini, C. minos, A. subsinuatum. Типичные для Бореальной надобласти устрицы на Мангышлаке отсутствуют. Исключение составляет только вид D. delta, характерный для верхней юры суббореальных бассейнов и встречающийся в берриасе Мангышлака. Однако и этот вид исчезает уже к верхам берриаса.
Список литературы
Атлас руководящих форм ископаемых фаун СССР. Том 10. Нижний отдел меловой системы. М.: Гос. изд-во геол. лит-ры, 1949. 328 с.
Атлас нижнемеловой фауны Северного Кавказа и Крыма. М.: Гос. науч.-тех. изд-во нефт. и горно-топл. лит-ры, 1960. 702 с.
Атлас раннемеловой фауны Грузии // Тр. Геол. ин-та АН Грузии. Нов. Сер. 2005. Вып. 120. 1–788 с.
Барабошкин Е.Ю. Раннемеловые проливы Русской плиты // Бюлл. МОИП, отд. геол. 2003. Т. 78. Вып. 4. С. 35–48.
Барабошкин Е.Ю. Раннемеловые проливы Северного полушария // Проливы Северного полушария в мелу и палеогене. М.: Изд-во геол. ф-та МГУ, 2007. С. 11–59.
Богданова Т.Н. Пелициподы валанжина Копет-Дага и их стратиграфическое распределение // Тр. ВСЕГЕИ. Нов. сер. 1961. Т. 46. Вып. 2. С. 126–151.
Богданова Т.Н. Берриасские и валанжинские устрицы Мангышлака // Второй Всесоюзный симп. по морфологии, системе, филогении и экогенезу двустворчатых моллюсков. Тез. докл. Тирасполь: Изд-во ТГПИ, 1978. С. 8–10.
Богданова Т.Н. Новые раннемеловые устрицы Мангышлака // Новые виды древних растений и беспозвоночных СССР. Вып. 5. М.: Наука, 1980. С. 26–28.
Берриас Горного Крыма. СПб.: “ЛЕМА”, 2012. 472 с.
Берриас Мангышлака. Л.: Наука, 1988. 204 с.
Валанжин Мангышлака. М.: Наука, 1983. 120 с.
Герасимов П.А. Руководящие ископаемые мезозоя центральных областей европейской части СССР. Часть 1. М.: Госгеолтехиздат, 1955. 379 с.
Захаров В.А. О позднеюрских и раннемеловых устрицах в арктических морях Сибири // Палеонтол. журн. 1963. № 4. С. 44–52.
Захаров В.А. Позднеюрские и раннемеловые двустворчатые моллюски севера Сибири (отряд Anisomyaria) и условия их существования. М.: Наука, 1966. 190 с.
Захаров В.А., Месежников М.С. Волжский ярус Приполярного Урала. Новосибирск: Наука, 1974. 216 с.
Косенко И.Н. О позднеюрских–раннемеловых устрицах рода Deltoideum Rollier (Bivalvia, Ostreoidea) Сибири // Палеонтол. журн. 2016. № 4. С. 21–30.
Миркамалов Х.Х. Классификация экзогир // Бюлл. МОИП, отд. геол. 1963. Т. 38. Вып. 5. С. 152–153.
Миркамалов Х.Х. Экзогиры, их систематика и значение для стратиграфии меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: ФАН УзССР, 1966. 133 с.
Мордвилко Т.А. Основные горизонты с фауной пелиципод в разрезах нижнего мела Мангышлака // Тр. ВНИГРИ. Нов. Сер. 1953. Вып. 73. С. 337–352.
Прозоровский В.А., Коротков В.А., Мамонтова Е.В. и др. Неоком Западной Туркмении // Тр. Всес. науч.-иссл. геол. ин-та. Нов. Сер. 1961. Т. 51. С. 1–234.
Ренгартен В.П. Фауна меловых отложений Ассинско-Камбилеевского района на Кавказе // Тр. Геол. ком. Н. сер. 1926. Вып. 147. С. 1–132.
Савельев А.А., Василенко В.П. Фаунистическое обоснование стратиграфии нижнемеловых отложений Мангышлака // Тр. ВНИГРИ. 1963. Вып. 218. С. 248–300.
Семенов В.П. Фауна юрских образований Мангышлака и Туаркыра // Тр. СПб. об-ва естествоиспыт. 1896. Т. 24. С. 1–79.
Семенов В.П. Фауна меловых образований Мангышлака и некоторых пунктов Закаспийского края // Тр. СПб. об-ва естествоиспыт. 1899. Т. 28. С. 1–178.
Стратиграфия СССР. Меловая система. Полутом 1. М.: Недра, 1986. 340 с.
Эристави М.С. Сопоставление меловых отложений Грузии и Крыма. М.: Изд-во АН СССР, 1957. 83 с.
Cooper M.R. Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Part 1 // Durban Mus. Novit. 1995. V. 20. P. 1–48.
Dhondt A.V., Dieni I. Early Cretaceous bivalves of Eastern Sardinia // Mem. Scienze Geol. 1988. V. 40. P. 1–97.
Freneix S. Disparité microstructurale du test entre Gryphaeostrea et Gyrostrea (huitres du Crétacé – Miocène). Novelle classification du genres // Ann. Paléontol. (Invertébr.). 1982. V. 66. № 3. P. 67–134.
Imlay R.W. Lower Neocomian fossils from the Miquihuana Region, Mexico // J. Paleontol. 1937. V. 11. № 4. P. 552–574.
Kosenko I.N. Late Jurassic–Early Cretaceous oysters from Siberia: A systematic review // Acta Palaeontol. Pol. 2017. V. 62. № 4. P. 759–778.
Kosenko I.N. The origin of the Pycnodonteinae and relationship between gryphaeas and true pycnodontes // Acta Palaeontol. Pol. 2018. V. 63. № 4. P. 769–778.
Leymerie M.A. Mémoire sur le terrain Crétacé de l’Aube. Partie 2 // Mém. Soc. géol. France. 1842. T. 5, pt. 1. P. 1–34.
Malchus N. Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik) // Berlin. Geowiss. Abh. Reihe A. 1990. Bd 125. S. 1–231.
Malchus N. Palaeobiogeography of Cretaceous oysters (Bivalvia) in the Western Tethys // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1996. H. 77. P. 165–181.
Pugaczewska H. Neocomian oysters from Central Poland // Acta Palaeontol. Pol. 1975. V. 20. № 1. P. 47–72.
Rubilar A.E., Lazo D.G. Description of Aetostreon pilmatuegrossum sp. nov. from the Lower Cretaceous of Argentina (Nequén Basin), and significance of the conservative left valve morphology in oysters of the genus Aetostreon Bayle // Cret. Res. 2009. V. 30. P. 727–748.
Wollemann A. Die Bivalven und Gastropoden des deutschen und holländischen Neocoms // Abh. König. Preuss. Geol. Landesanst. N. F. 1900. H. 31. S. 1–180.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал