

Палеонтологический журнал, 2020, № 3, стр. 32-42
Новые данные о байос-батских Parkinsoniidae (Ammonoidea, Perisphinctoidea)
В. В. Митта a, c, *, Ю. А. Бакарюкина a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
Москва, Россия
c Череповецкий государственный университет
Москва, Россия
* E-mail: mitta@paleo.ru
** E-mail: bakaryukina2010@yandex.ru
Поступила в редакцию 18.06.2019
После доработки 24.06.2019
Принята к публикации 24.06.2019
Аннотация
Проанализированы некоторые особенности строения лопастной линии взрослых представителей сем. Parkinsoniidae. Установлено, что у родов Caumontisphinctes, Pseudocosmoceras, Sokurella и большинства Rarecostites вентральная лопасть (V) глубже первой боковой (U). У микроконхов поздних Rarecostites, а также у Parkinsonia и Oraniceras, напротив, лопасть V мельче лопасти U; у макроконхов они могут быть примерно равными по глубине. С учетом данных по морфогенезу раковины и скульптуры и хроностратиграфической последовательности таксонов предложена схема филогенеза семейства в объеме подсемейств Parkinsoniinae Buckman (роды Caumontisphinctes, Rarecostites, Parkinsonia и Oraniceras) и Pseudocosmoceratinae Sasonov (роды Pseudocosmoceras и Sokurella). Подтверждается правильность отнесения Djanaliparkinsonia к подсем. Garantianinae (Stephanoceratidae), на основе сходства строения лопастной линии.
ВВЕДЕНИЕ
Аммониты семейства Parkinsoniidae распространены в позднем байосе и раннем бате по всей северной периферии океана Тетис от Центральной Азии до Западной Европы и имеют важное значение для подразделения и корреляции вмещающих отложений. Семейство было установлено С. Бакменом (Buckman, 1920). Изначально в его состав было вполне определенно включено восемь родов (Parkinsonia, Strenoceras, Garantiana, Baculatoceras, Subparkinsonia, Bigotites, Pseudobigotella, Caumontisphinctes) и, кроме этого, условно – еще три рода (Choffatia, Leptosphinctes, Vermisphinctes).
За прошедшее столетие объем семейства неоднократно менялся, как за счет установления принадлежности части перечисленных родов к семействам Stephanoceratidae и Perisphinctidae, так и за счет отнесения к Parkinsoniidae вновь установленных родов. В итоге из первоначального перечня, помимо номинального рода Parkinsonia, в составе Parkinsoniidae к настоящему времени остался только род Caumontisphinctes. Неоднократно ставился под сомнение и ранг Parkinsoniidae – некоторые исследователи предпочитали рассматривать его как подсемейство, в составе сем. Stephanoceratidae или сем. Perisphinctidae. Различались также мнения о происхождении паркинсониид – являются они непосредственными дериватами Stephanoceratoidea, или же должны относиться к Perisphinctoidea, как таксон, происходящий от Perisphinctidae (впрочем, последние тоже ведут свое начало от стефаноцератоидей). Указанные различия в понимании ранга таксона, его объема и происхождения обсуждались или приведены в многочисленных специальных публикациях и сводных работах (Arkell, 1957; Schindewolf, 1965; Donovan et al., 1981; Безносов, Кутузова, 1982; Schlegelmilch, 1985 и др.). В составе Parkinsoniidae было выделено также подсемейство Pseudocosmoceratinae с типовым родом Pseudocosmoceras Murashkin (Сазонов, 1960).
Мы понимаем Parkinsoniidae как семейство в объеме двух подсемейств: Parkinsoniinae и Pseudocosmoceratinae. В составе Parkinsoniinae Buckman, 1920 нами рассматриваются роды Caumontisphinctes Buckman, 1920 [M, m] (включая Infraparkinsonia Westermann, 1956, как младший субъективный синоним), Rarecostites Besnosov et Kutuzova, 1982 [M, m], Parkinsonia Bayle, 1878 [M, m] (включая Gonolkites Buckman, 1925 и Durotrigensia Buckman, 1928, как младшие субъективные синонимы) и Oraniceras Flamand, 1911 [M, m]. Сюда же относится очень плохо изученный, известный лишь из Грузии род Okribites Kakhadze, 1937. В составе Pseudocosmoceratinae Sasonov, 1960 нами рассматриваются роды Pseudocosmoceras Murashkin, 1930 [M, m] (включая Medvediceras Nikolaeva, 1967, как младший субъективный синоним) и Sokurella Mitta, 2004 [M, m].
Несмотря на долгую историю изучения, филогенетические взаимоотношения паркинсонииид остаются недостаточно ясными. Филетическая линия Caumontisphinctes → Raracostites → Parkinsonia → Oraniceras номинального подсемейства, несмотря на некоторые лакуны, представляется вполне логичной и подтверждается хроностратиграфической последовательностью таксонов. Филогенез Pseudocosmoceratinae, ареал которых ограничен акваторией позднебайосского–раннебатского Южнорусского морского бассейна (южная часть Русской платформы от Нижнего Поволжья на востоке до Донбасса на западе и Северный Кавказ) изучен еще явно неполно. В статье предпринята попытка реконструкции филогенеза Parkinsoniidae с учетом особенностей строения лопастных линий взрослых раковин. Данные по изменению формы раковины и скульптуры паркинсониид в процессе их исторического развития присутствуют в большинстве процитированных работ и ниже рассматриваются лишь в необходимом объеме.
Цифровые фотографии аммонитов выполнены С.В. Багировым (Палеонтологический ин-т им. А.А. Борисяка РАН, ПИН), которому авторы выражают глубокую благодарность. Зарисовки лопастных линий выполнены одним из авторов, Ю.А. Бакарюкиной. Аммониты, чьи изображения и лопастные линии приведены в работе, хранятся в ПИН (Москва), Государственном геологическом музее им. В.И. Вернадского РАН (ГГМ, Москва) и Центральном научно-исследовательском геолого-разведочном музее им. акад. Ф.Н. Чернышева (ЦНИГРМ, С.-Петербург).
РЕЗУЛЬТАТЫ ИЗУЧЕНИЯ ЛОПАСТНЫХ ЛИНИЙ
Детальные исследования онтогенеза лопастной линии различных таксонов Parkinsoniidae, проведенные предшествующими исследователями, не привели к консенсусу о систематическом составе и филогении паркинсониид. Мнение одного из крупнейших исследователей лопастных линий мезозойских аммонитов О. Шиндевольфа о принадлежности Parkinsoniidae к Stephanoceratoidea (Schindewolf, 1923, 1965) в последние несколько десятилетий поддерживалось только Р. Шлегельмильхом (Schlegelmilch, 1985). Немало сил отдал изучению онтогенезов и отдельных лопастных линий юрских аммонитов, в том числе паркинсониид, Н.В. Безносов, считавший несомненной их принадлежность к перисфинктоидеям (Безносов, Кутузова, 1982; Безносов, Митта, 1993, 1995).
Наши попытки изучить полный онтогенез лопастной линии позднебайосских–раннебатских аммонитов не увенчались успехом из-за сохранности материала, представленного преимущественно не-кальцитовыми ядрами, иногда с остатками раковины. Поэтому авторами проведено сравнительное изучение отдельных лопастных линий различных видов Parkinsoniidae и некоторых других одновозрастных таксонов, для возможного прояснения вопросов их филогении. Такие исследования уже проводились в истории изучения паркинсониид, когда зарисовки отдельных лопастных линий (Wetzel, 1911; Nicolesco, 1928) использовались для характеристики видов, что позволило в итоге проследить эволюционную линию паркинсониин (“Parkinsonia” s.l. у указанных авторов) от Caumontisphinctes до Oraniceras.
На рис. 1 представлены зарисовки лопастных линий северокавказских Rarecostites – взрослых микроконхов, молодого макроконха сходного с ними размера, и взрослого, судя по размерам, макроконха. На всех зарисовках видно, что глубина первой боковой лопасти (U) заметно меньше, чем вентральной лопасти (V). Это особенно выражено у микроконхов и молодого макроконха, и меньше у взрослого макроконха. Такое же соотношение наблюдается у следующего вида этого рода, R. donezianus (Borissjak) – вентральная лопасть заметно глубже у микроконха и почти равна по глубине первой боковой лопасти на взрослых стадиях макроконха (рис. 2).
Рис. 1.
Лопастные линии Rarecostites: а – R. sherstyukovi Mitta, взрослый микроконх, паратип ПИН, № 5546/67, при В = 13 мм; б – R. subarietis (Wetzel), взрослый микроконх, экз. ПИН, № 5546/36 (Митта и др., 2018, табл. II , фиг. 4), при В = 15 мм; в – R. kyafarensis Mitta, молодой макроконх, экз. ПИН № 5546/68 (изображен на табл. VI , фиг. 1; см. вклейку), при В = 16.5 мм; г – R. mutabilis (Nicolesco), взрослый макроконх, экз. ПИН, № 5546/145 (Митта и др., 2018, табл. II , фиг. 1), при В = 44 мм.
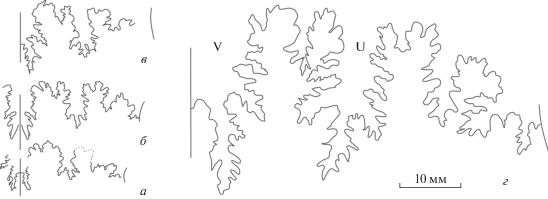
Рис. 2.
Лопастные линии Rarecostites donezianus (Borissjak): а – микроконха (по: Борисяк, 1908, табл. IX , фиг. 6b); б – макроконха (по: Борисяк, 1908, табл. IX , фиг. 6d; аммонит изображен там же, табл. VII , фиг. 1).

Литературные данные не совсем совпали с нашими наблюдениями. Так, В. Ветцель (Wetzel, 1911, рис. 31–35 ) изобразил лопастные линии установленных им видов Parkinsonia subarietis и P. acris (относимых нами к роду Rarecostites, при этом второе видовое название является младшим субъективным синонимом первого; см. Митта, 2017). На этих зарисовках соотношение глубины лопастей V и U варьирует – то одна из них глубже, то они равны. К сожалению, из работы Ветцеля остается неясным, с каких экземпляров (макроконхов или микроконхов) были зарисованы лопастные линии. Но, пересмотрев несколько десятков раковин северокавказских рарекоститов, мы все же нашли экземпляр микроконха, у которого лопасти V и U равны по глубине на взрослой раковине (рис. 3).
Рис. 3.
Лопастная линия Rarecostites sherstyukovi Mitta, микроконх, экз. ПИН, № 5546/77 (изображен в: Митта, 2017, рис. 5, а–в), при В = 10 мм.

К. Николеско также изобразил лопастные линии “Parkinsonia subarietis” и “P. оrbignyana” (Nicolesco, 1928, рис. 9, 10), у которых лопасть U заметно глубже лопасти V. Аммониты, с которых зарисованы эти линии, изображены в работе Николеско и являются микроконхами; по нашему мнению, они относятся к Rarecostites rarecostatus (Buckman).
Предковым для Rarecostites (и всех остальных паркинсониид) считается род Caumontisphinctes. Нам удалось зарисовать лопастную линию лишь у одного микроконха этого рода, у которого глубина первой боковой лопасти оказалась заметно меньше глубины вентральной (рис. 4). Зарисовки лопастных линий нескольких макроконхов и микроконхов этого рода привел Г. Дитль (Dietl, 1980, рис. 3, 5), и у всех этих аммонитов лопасть V также глубже лопасти U.
Рис. 4.
Лопастная линия Caumontisphinctes aff. gruibingensis Dietl, микроконх, экз. ПИН, № 5546/190 (изображен на табл. VI , фиг. 8), при В = 7 мм.
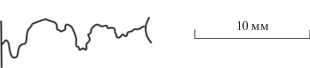
Рис. 5.
Лопастные линии Parkinsonia sp.: а – микроконх, экз. ПИН, № 5546/230 (изображен на табл. VII , фиг. 2; см. вклейку), при В = 14.5 мм; б – молодой макроконх, экз. ПИН, № 5546/240 (изображен на табл. VII , фиг. 1), при В = 19 мм.
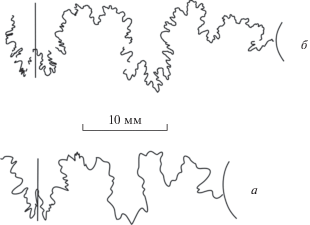
На рис. 5 приведены зарисовки лопастных линий микроконха и макроконха рода Parkinsonia, не достигших еще зрелости. Хорошо видно, что глубина вентральной лопасти этих аммонитов заметно меньше глубины первой боковой. Раковины паркинсоний в нашей коллекции в большинстве своем оказались непригодными для изучения лопастной линии из-за своей сохранности, кроме нескольких экземпляров. Однако лопастные линии этого рода хорошо охарактеризованы в уже упомянутой выше работе Николеско (Nicolesco, 1928). Этим исследователем наглядно показано, что на ранних стадиях развития лопасть V глубже лопасти U, а по мере взросления это соотношение оказывается обратным.
Следующий представитель подсемейства Parkinsoniinae – род Oraniceras. Несмотря на достаточно хорошую выборку этого рода в наших коллекциях, удалось зарисовать лопастные линии микроконхов только фрагментарно, для одного вида (рис. 6). Но и по этим зарисовкам видно, что первая боковая лопасть у этих аммонитов заметно глубже, чем вентральная. Лопастные линии макроконхов Oraniceras зарисованы для нескольких видов (рис. 7), в этом случае лопасти U и V практически равны по глубине.
Рис. 6.
Лопастные линии микроконхов Oraniceras scythicum Mitta: а – экз. ПИН, № 5546/141 (изображен в: Митта и др., 2017, табл. I , фиг. 1), при В = 13 мм; б – экз. ПИН, № 5546/4 (изображен в: Митта, 2015, табл. III , фиг. 3), при В = 14 мм; в – экз. ПИН, № 5546/232 (изображен на табл. VI , фиг. 2), при В = = 22 мм; г – экз. ПИН, № 5546/3 (изображен в: Митта, 2015, табл. III , фиг. 2), при В = 22 мм; д – экз. ПИН № 5546/6 (изображен в: Митта, 2015, табл. III , фиг. 5), при В = 24 мм.

Рис. 7.
Лопастные линии макроконхов Oraniceras: а – O. mojarowskii (Masarowich), экз. ГГМ, № 09441 (изображен в: Мурашкин, 1930, табл. VII , фиг. 1–3; Гуляев, 2019, табл. I , фиг. 2), при В = 70.5 мм; б – O. besnosovi Mitta et Seltzer, экз. ГГМ, № 09706 [CR-2779] (изображен в: Митта и др., 2004, табл. 2 , фиг. 2), при В = 63.5 мм; в – O. gyrumbilicum (Quenstedt), экз. ПИН, № 5546/257 (изображен на табл. VII , фиг. 3), при В = 55.5 мм.
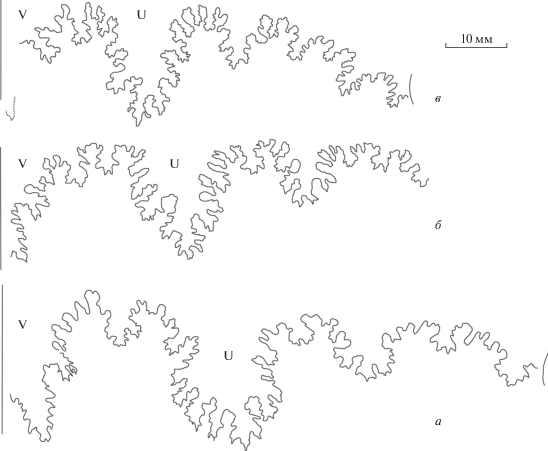
Зарисовки лопастных линий представителей рода Pseudocosmoceras хорошо представлены в работах А.А. Борисяка (1908, рис. 5, a–c), и П.К. Мурашкина (1930, рис. 6–8, 10, 12). Мы воспроизводим здесь часть этих рисунков, а также приводим наши собственные зарисовки лопастной линии различных таксонов этого подсемейства (рис. 8). Хорошо видно, что глубина первой боковой лопасти Pseudocosmoceras обычно вдвое меньше глубины вентральной – как у микроконхов, так и у макроконхов. У макроконха рода Sokurella, с мелкоразмерной раковиной, эта разница менее заметна (рис. 8, д).
Рис. 8.
Лопастные линии Pseudocosmoceratinae: а, б – Pseudocosmoceras michalskii (Borissjak): а – ювенильная раковина (по: Борисяк, 1908, табл. IX , фиг. 5a; аммонит изображен там же, табл. III , фиг. 17 ), б – макроконх (по: Борисяк, 1908, табл. IX , фиг. 5c); в – P. inclarum Murashkin, экз. ГГМ, № 09447 [VI-62/9] (изображен в: Мурашкин, 1930, табл. 9 , фиг. 18–20 ; Митта, 2004, табл. III , фиг. 8), при В = 9 мм; г – P. masarowici Murashkin, макроконх, экз. ГГМ, № 09445 [VI-62/7] (изображен в: Мурашкин, 1930, табл. 9 , фиг. 13, 14, 17 ; Митта, 2004, табл. IV , фиг. 1), при В = 25.5 мм; д – Sokurella galaczi Mitta, макроконх, экз. ГГМ, № 09673 [CR-2698а] (изображен в: Митта и др., 2004, табл. 1 , фиг. 3; здесь, табл. VI , фиг. 3), при В = 16 мм.
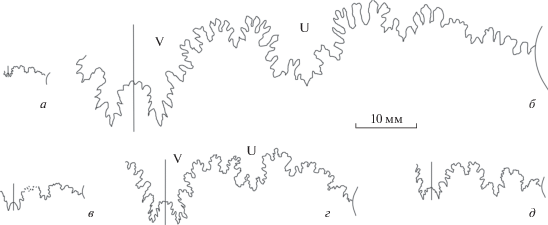
Рис. 9.
Лопастные линии Garantianinae: а–г – Djanaliparkinsonia alanica Mitta: а – взрослый микроконх, экз. ПИН, № 5546/224 (изображен на табл. VI , фиг. 6) при В = 19 мм; б – молодой макроконх, экз. ПИН, № 5546/225 (изображен на табл. VI , фиг. 5) при В = 17 мм; в – взрослый макроконх, экз. ПИН, № 5546/223 (изображен на табл. VI , фиг. 4) при В = 43 мм; г – взрослый макроконх, экз. ПИН, № 5546/197 (изображен в: Митта, 2018, табл. V, VI ), при В = 37 мм; д – Garantiana subgaranti Wetzel, почти взрослый макроконх, экз. ПИН, № 5546/140 (изображен на табл. VI , фиг. 7) при В = 16 мм.
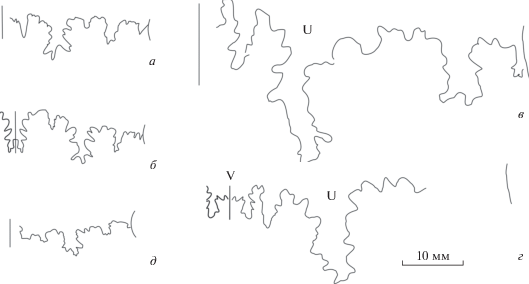
Рис. 10.
Лопастная линия Leptosphinctes sp., взрослый макроконх, экз. ПИН, № 5546/247 (изображен на табл. VI , фиг. 9), при В = 22.5 мм.
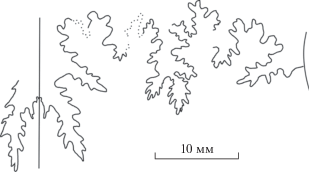
Результаты изучения были бы недостаточно корректными без сравнения лопастных линий паркинсониид с таковыми близкородственных стефаноцератид и перисфинктид из сходных по возрасту отложений. Поэтому были сделаны зарисовки лопастных линий некоторых других аммонитин позднего байоса. У представителей рода Djanaliparkinsonia, изначально описанного как подрод рода Parkinsonia (Кутузова, 1975) в семействе Parkinsoniidae, лопастная линия имеет иные очертания. Сравнительно узкая первая боковая лопасть значительно глубже вентральной, особенно у макроконхов; кроме того, лопастные линии у этих аммонитов гораздо менее рассеченные, чем у паркинсониид при сходных размерах (рис. 9, a–г). В то же время контур лопастной линии джаналипаркинсоний очень сходен с таковым у рода Garantiana (подсем. Garantianinae семейства Stephanoceratidae) (рис. 9, д).
Гораздо ближе к паркинсониидам лопастная линия Leptosphinctes (Perisphinctidae, подсем. Leptosphinctinae). Отчетливо трехраздельная, как и у паркинсониид, первая боковая лопасть у этого рода значительно мельче вентральной (рис. 10).
ОБСУЖДЕНИЕ
Сходство лопастной линии Parkinsoniidae и Perisphinctidae свидетельствует о правильности объединения их в одном надсемействе Perisphinctoidea (Arkell, 1950; Westermann, 1956; и др.) и близком родстве первых паркинсониид с Leptosphinctes (Pavia, 1973; Безносов, Кутузова, 1982). Большие отличия лопастной линии Djanaliparkinsonia от таковой паркинсониид и несомненное сходство с лопастной линией Garantiana указывают на правильность выведения рода Djanaliparkinsonia из состава семейства Parkinsoniidae и отнесения к Stephanoceratoidea (Stephanoceratidae, Garantianinae) (Митта, 2018).
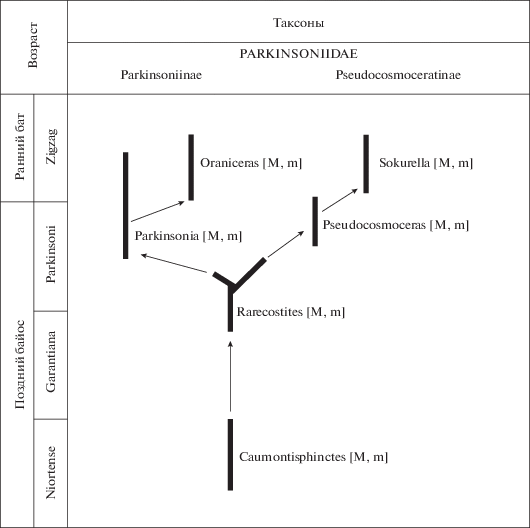
Суммируя полученные нами результаты с данными предшествующих исследователей, можно считать установленным, что соотношение глубины вентральной и первой боковой лопастей меняется как в онтогенезе отдельных таксонов, так и в филогенезе Parkinsoniidae, и является важным признаком для реконструкции эволюции этого семейства. На ранних стадиях развития раковины у всех паркинсониид вентральная лопасть глубже первой боковой. Это соотношение сохраняется и для лопастной линии взрослых раковин древнейших (расцвет в хрон Niortense) представителей семейства, рода Caumontisphinctes – как для микроконхов, так и для макроконхов. У более молодых Rarecostites (расцвет в фазу Subarietis) в строении лопастной линии начинаются эволюционные изменения. У большинства микроконхов Rarecostites лопасть V все еще глубже лопасти U, но у микроконха последнего (западноевропейского) представителя рода, R. rarecostatus (Buckman) первая боковая лопасть на взрослой раковине становится заметно глубже вентральной. Отметим сразу, что такое изменение сохраняется у микроконхов Parkinsoniinae и далее, у родов Parkinsonia и Oraniceras; у макроконхов этих таксонов различия в глубине лопастей V и U несущественны, что можно объяснить большей природной консервативностью самок.
В эндемичной линии Pseudocosmoceratinae (Pseudocosmoceras → Sokurella) соотношение глубины лопастей остается архаичным: лопасть V всегда глубже лопасти U, независимо от принадлежности к микроконхам или макроконхам. Очевидно, что это дериваты рода Rarecostites, исходя в том числе из стратиграфического распространения, совпадающего с таковым Parkinsonia и Oraniceras (верхи хронозоны Parkinsoni – низы хронозоны Zigzag).
Типичные представители Rarecostites rarecostatus, завершающего филолинию микроконхов этого рода в Западной Европе, как показывают наши последние исследования, в Южнорусском бассейне отсутствуют. Место этого вида, по-видимому, здесь занимает R. donezianus, впервые описанный из Донбасса (Борисяк, 1908) и только недавно обнаруженный на Северном Кавказе (Митта и др., 2017). Тип этого вида не был до сих пор обозначен (обсуждение см.: Митта, 2017), поэтому мы обозначаем здесь как лектотип экземпляр, изображенный Борисяком (1908, табл. IV , фиг. 3), и приводим фотографии – его и одного из паралектотипов (рис. 11, а–г). Для R. donezianus характерны длинные первичные ребра, с бугорками в точке ветвления очень близко к наружной стороне; на вентральной стороне располагаются еще два ряда бугорков, отделенных срединной бороздой.
Рис. 11.
Оригиналы к работе Борисяка (1908): а–г – Rarecostites donezianus (Borissjak), а–в – лектотип ЦНИГРМ, № 77/314: а – с устья, б – сбоку, в – с вентральной стороны; г – паралектотип ЦНИГРМ, № 78/314, сбоку; д, е – Pseudocosmoceras michalskii (Borissjak), лектотип ЦНИГРМ, № 71/314: д – с вентральной стороны, е – сбоку.
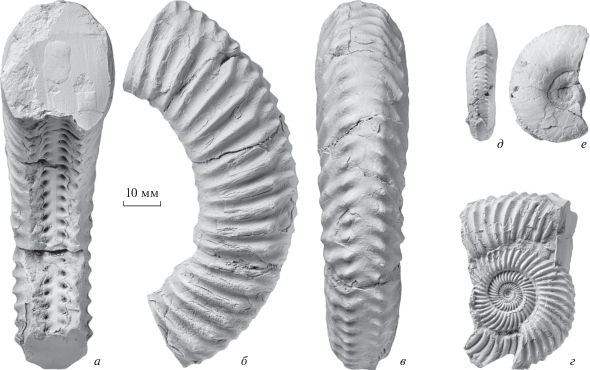
В верхней части подзоны Subarietis на р. Кяфар (Карачаево-Черкесия) изредка встречаются аммониты, отличающиеся от типичных R. subarietis (Wetzel) длинными и гуще расположенными первичными ребрами с бугорками в точке ветвления и гребневидно приподнятыми на вентральной стороне кончиками ветвей. Эта аберрация, скорее всего, и привела в итоге к появлению R. donezianus, найденных на Кяфаре выше по разрезу, в слоях с Parkinsonia djanelidzei.
Первые Pseudocosmoceras отличаются выраженным трендом к еще большему удлинению первичных ребер со слиянием латерального и вентрального ряда бугорков и трансформации их в мелкие зубчики, при этом на ядрах первичные ребра сглаживаются (ослабленная скульптура является диагностическим признаком для всего подсемейства Pseudocosmoceratinae). Типовой экземпляр для типового вида рода, P. michalskii (Borissjak), также не был до сих пор предложен. Мы обозначаем здесь как лектотип экземпляр, изображенный Борисяком (1908, табл. III , фиг. 18 ), и приводим его фотографии (рис. 11, д, е).
Род Sokurella известен лишь по типовому виду, S. galaczi Mitta. В недавней работе Д.Б. Гуляева (2019) сокуреллы рассматриваются в ранге подрода рода Oraniceras в качестве микроконхов последнего, включенного Гуляевым в подсемейство Pseudocosmoceratinae. К сожалению, этот исследователь не обратил внимания, что голотип Sokurella galaczi (Митта, 2004, табл. III , фиг. 1) представляет собой взрослую раковину с типичной для макроконхов паркинсониид жилой камерой со слабо выраженной скульптурой и простым (и хорошо сохранившимся) устьевым краем. Микроконхи сокурелл вдвое меньше в диаметре и сохраняют хорошо выраженные вторичные ребра и на жилой камере (там же, фиг. 2, экз. в нижней части конкреции). Рельефность скульптуры микроконхов на взрослой жилой камере (по сравнению с “лысеющими” с возрастом макроконхами) характерна и для других паркинсониид, да и среднеюрских перисфинктоидей в целом. Это подтверждается изображениями взрослых микроконхов Oraniceras (Hahn, 1970, табл. 3 , фиг. 3–5, 11, 12; Dietl, 1977, табл. 5 , фиг. 3; Callomon et al., 1987, табл. 2 , фиг. 2; Dietze, Dietl, 2006, табл. 9 , фиг. 1; Митта, 2015, табл. III , фиг. 2; табл. IV , фиг. 3). Впрочем, установленные Гуляевым новые виды “Sokurella” pletniovkae и “S.” elshankae также сохраняют хорошо выраженную на жилой камере скульптуру. По всей вероятности, “S.” pletniovkae представляет собой микроконх вида Oraniceras mojarowskii (Masarowich), а названия Oraniceras calvum Gulyaev и O. issae Gulyaev являются младшими субъективными синонимами O. mojarowskii, установленного А.Н. Мазаровичем (1923) по макроконху. Сохранность типового материала по новым таксонам в статье Гуляева оставляет желать лучшего, и экземпляры, указанные там как Sokurella elshankae Gulyaev, мы можем определить лишь как Parkinsonia ? sp.
Гуляев рассматривает род Oraniceras в составе Pseudocosmoceratinae, предполагая его происхождение от Pseudocosmoceras. Мы полагаем, что взрослые макроконхи первых представителей Oraniceras – O. scythicum на Сев. Кавказе (Митта, 2015) и O. “n. sp. aff. fretense” в Юж. Германии (Dietze, Dietl, 2006) – еще очень близки к макроконхам, ранее выделявшимся как Gonolkites поздних Parkinsonia. Хорошо выраженная скульптура Oraniceras на ранних стадиях развития и приведенные выше данные по строению лопастной линии свидетельствуют в пользу отнесения этого рода к Parkinsoniinae. В качестве переходного звена от Parkinsonia к Oraniceras может рассматриваться форма, описанная из пограничных отложений байоса и бата Юж. Германии как Parkinsonia cf. vermicularis (Buckman) (Dietl et al., 1983, рис. 2, а).
Филогенетическая схема семейства Parkinsoniidae, составленная в соответствии с представлениями авторов, приведена на рис. 12.
ЗАКЛЮЧЕНИЕ
Филогенез семейства Parkinsoniidae был в общих чертах выяснен в последние десятилетия и без изучения особенностей лопастной линии – на основе уточнения стратиграфической последовательности таксонов разного ранга и выявления эволюционных трендов в развитии внешней морфологии родов. Тем не менее, сравнительное изучение даже отдельных элементов лопастной линии взрослых (и не совсем взрослых) раковин оказалось довольно полезным для решения спорных вопросов систематики. При этом выявились и любопытные расхождения в эволюционных процессах по разным группам признаков.
Система паркинсониид выстроена, прежде всего, на основе различий в развитии скульптуры. У представителей подсемейства Parkinsoniinae скульптура рельефная и сохраняется обычно в том или ином виде практически до конца фрагмокона, а у Pseudocosmoceratinae ребра слабо выражены, начиная с ювенильных оборотов. Но если брать за основу систематики признак “лопасть V глубже лопасти U”, то линия Caumontisphinctes → → Rarecostites → Pseudocosmoceras → Sokurella является основным стволом развития семейства, со стабильным сохранением “архаичного” строения лопастной линии, а линия Rarecostites rarecostatus → Parkinsonia → Oraniceras лишь боковой ветвью. Заметим, что правило “лопасть V глубже лопасти U” распространяется на многих, если не на всех, байос–батских перисфинктид, от которых, собственно, и происходят паркинсонииды. Важным наблюдением мы считаем некоторую консервативность макроконхов поздних Parkinsoniinae, у которых изменение контура лопастной линии происходит медленнее, чем у микроконхов. Возможно, что это еще одно подтверждение правильности отнесения макроконхов к самкам.
В микроэволюции паркинсониид остается еще достаточно много невыясненных вопросов. В первую очередь, это проблема номинального рода семейства – является ли происхождение Parkinsonia монофилетическим, или же часть видов, относящихся в настоящее время к паркинсониям, произошла от других таксонов, в различных бассейнах? Какое место в филогении Parkinsoniidae занимает род Okribites? Мы надеемся со временем ответить хотя бы на часть этих вопросов.
Список литературы
Безносов Н.В., Кутузова В.В. Систематика паркинсониид (Ammonitida) // Палеонтол. журн. 1982. № 3. С. 41–52.
Безносов Н.В., Митта В.В. Позднебайосские и батские аммонитиды Северного Кавказа и Средней Азии. М.: Недра, 1993. 347 с.
Безносов Н.В., Митта В.В. Полимофизм юрских аммоноидей // Палеонтол. журн. 1995. № 2. С. 41–50.
Борисяк А. Фауна Донецкой юры. 1. Cephalopoda // Тр. Геол. ком. Нов. сер. 1908. Вып. 37. С. 1–94.
Гуляев Д.Б. Аммониты и инфразональная стратиграфия зоны besnosovi нижнего бата Русской плиты // Стратигр. Геол. корреляция. 2019. Т. 27. № 1. С. 103–125.
Кутузова В.В. Djanaliparkinsonia – новый подрод рода Parkinsonia Bayle из верхнего байоса Юго-Западного Гиссара // Новые данные по стратиграфии мезозойских отложений нефтегазоносных регионов юга СССР. М.: ВНИГНИ, 1975. С. 89–92 (Тр. ВНИГНИ. Вып. 171).
Мазарович А.Н. Среднеюрские отложения реки Иловли // Вестн. Моск. горн. акад. 1923. Т. 2. № 1. С. 29–60.
Митта В.В. Sokurella galaczi gen. et sp. nov. и другие среднеюрские Parkinsoniidae (Ammonoidea) Нижнего Поволжья // Палеонтол. журн. 2004. № 3. С. 30–35.
Митта В.В. Род Oraniceras (Parkinsoniidae, Ammonoidea) в нижнем бате юга европейской части России // Палеонтол. журн. 2015. № 6. С. 38–42.
Митта В.В. О некоторых Rarecostites (Parkinsoniidae, Ammonoidea) зоны Parkinsoni верхнего байоса Северного Кавказа // Палеонтол. журн. 2017. № 5. С. 13–26.
Митта В.В. О первых находках Djanaliparkinsonia (Stephanoceratidae, Ammonoidea) в верхнем байосе Северного Кавказа // Палеонтол. журн. 2018. № 4. С. 29–37.
Митта В.В., Барсков И.С., Грюндель Й. и др. Верхний байос и нижний бат в окрестностях Саратова // VM-Novitates. 2004. № 12. С. 1–39.
Митта В.В., Савельева Ю.Н., Федорова А.А., Шурекова О.В. Биостратиграфия пограничных отложений байоса и бата бассейна р. Большой Зеленчук (Северный Кавказ) // Стратигр. Геол. корреляция. 2017. Т. 25. № 6. С. 30–49.
Митта В.В., Савельева Ю.Н., Федорова А.А., Шурекова О.В. Аммониты, микрофауна и палиноморфы нижней части зоны Parkinsoni верхнего байоса бассейна р. Большой Зеленчук, Северный Кавказ // Стратигр. Геол. корреляция. 2018. Т. 26. № 5. С. 49–67.
Мурашкин П.К. Среднеюрские аммониты северной оконечности Доно-Медведицкого вала // Бюлл. Моск. об-ва испыт. природы. Отд. геол. 1930. Т. 38. Вып. 8. С. 139–159.
Сазонов Н.Т. Новые данные об оксфордских и кимериджских аммонитах // Тр. ВНИГНИ. 1960. Вып. 16. С. 133–161.
Arkell W.J. A classification of the Jurassic ammonites // J. Paleontol. 1950. V. 24. P. 356–364.
Arkell W.J. Jurassic ammonitina // Treatise of Invertebrate Paleontology. Pt. L. Mollusca. N.Y., Lawrence, 1957. P. 232–344.
Buckman S.S. Yorkshire Type Ammonites. Type Ammonites. Bd 1–7. L.: Wesley, 1909–1930. 790 pl.
Callomon J.H., Dietl G., Galácz A. et al. Zur Stratigraphie des Mittel- und unteren Oberjuras in Sengenthal bei Neumarkt/Opf. (Fränkische Alb) // Stuttg. Beitr. Naturk. Ser. B. 1987. № 132. 53 S.
Dietl G. The Braunjura (Brown Jurassic) in Southwest Germany // Stuttg. Beitr. Naturk. Ser. B. 1977. № 25. 41 p.
Dietl G. Die Ammoniten-Gattung Caumontisphinctes aus dem südwestdeutschen Subfurcaten-Oolith (Bajocium, Mittl. Jura) // Stuttg. Beitr. Naturk. Ser. B. 1980. № 51. 43 S.
Dietl G., Hugger R., Schaaf D. Die Lage der Bajocium/Bathonium-Grenze (Mittlerer Jura) in der südwestlichen Schwäbischen Alb, SW-Deutschland // Jh. Ges. Naturk. Württemberg. 1983. Bd 138. S. 75–84.
Dietze V., Dietl G. Feinstratigraphie und Ammoniten-Faunenhorizonte im Ober-Bajocium und Bathonium des Ipf-Gebietes (Schwäbische Alb, Südwestdeutschland) // Stuttg. Beitr. Naturk. Ser. B. 2006. № 162. 51 S.
Donovan D.R., Callomon J.H., Howarth K.M. Classification of the Jurassic Ammonitina // The Ammonoidea / Eds. House H.R., Senior J.R. L.: Acad. Press, 1980. P. 101–155 (Syst. Assoc. Spec. Pap. V. 18).
Hahn W. Die Parkinsoniidae S. Buckman und Morphoceratidae Hyatt (Ammonoidea) des Bathoniums (Brauner Jura ɛ) im südwestdeutschen Jura // Jh. Geol. Landesamt Baden-Württemberg. 1970. № 12. S. 7–62.
Nicolesco C.-P. Étude monographique du genre Parkinsonia // Mém. Soc. géol. France. N.S. 1927. T. 9. P. 1–84.
Pavia G. Ammoniti del Baiociano superiore di Digne (Francia SE, Dip. Basses-Alpes) // Boll. Soc. Paleontol. Ital. 1971 (1973). V. 10. № 2. P. 75–142.
Schindewolf O.H. Ueber die Ausgestaltung der Lobenlinie bei den Neoammonoideen Wdkd. // Centralbl. Miner. Geol. 1923. Bd 23. S. 337–350.
Schindewolf O.H. Studien zur Stammesgeschichte der Ammoniten. Lief. IV // Akad. Wiss. Literatur. Abh. math.-naturwiss. Lk. 1965. № 3. S. 407–508.
Schlegelmilch R. Die Ammoniten des süddeutschen Doggers: e. Bestimmungsbuch für Fossiliensammler u. Geologen. Stuttgart – N.Y.: Fischer, 1985. 284 s.
Westermann G.E.G. Phylogenie der Stephanocerataceae und Perisphinctaceae des Dogger // N. Jb. Geol. Paläontol. Abh. 1956. Bd 103. S. 233–279.
Wetzel W. Faunistische und stratigraphische Untersuchung der Parkinsonienschichten des Teutoburger Waldes bei Bielefeld // Palaeontogr. Ser. A. 1911. Bd 58. S. 139–277.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал