

Палеонтологический журнал, 2020, № 3, стр. 11-19
Представители рода Anmarginulina (Lagenida, Foraminifera) из позднего плинсбаха (ранняя юра) Арктики
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
b Новосибирский государственный университет
Новосибирск, Россия
* E-mail: NikitenkoBL@ipgg.sbras.ru
Поступила в редакцию 03.07.2019
После доработки 24.10.2019
Принята к публикации 29.11.2019
Аннотация
Представители рода Anmarginulina широко распространены в позднеплинсбахских арктических бассейнах. Группа имеет важное значение для биостратиграфии, межрегиональных корреляций и палеобиогеографии. Впервые проведено монографическое исследование позднеплинсбахских видов Anmarginulina из разрезов Арктической Сибири.
ВВЕДЕНИЕ
В юрских и меловых Арктических бассейнах обычно преобладали агглютинирующие таксоны фораминифер, численность отдельных видов иногда могла достигать десятков тысяч экземпляров на образец. Но на отдельных временных уровнях юры и мела, обычно связанных с глобальными событиями: потеплениями климата и эвстатическим подъемом уровня Мирового Океана, значительную роль в сообществах фораминифер Арктических бассейнов начинали играть представители Lagenida (Foraminifera). На эти моменты обычно резко вырастало их таксономическое разнообразие и количественная представительность.
Многие виды некоторых родов отряда Lagenida были широко распространены в Арктических бассейнах, относительно быстро эволюционировали (рис. 1) и поэтому имеют высокое значение для биостратиграфических исследований и разработки зональных шкал.
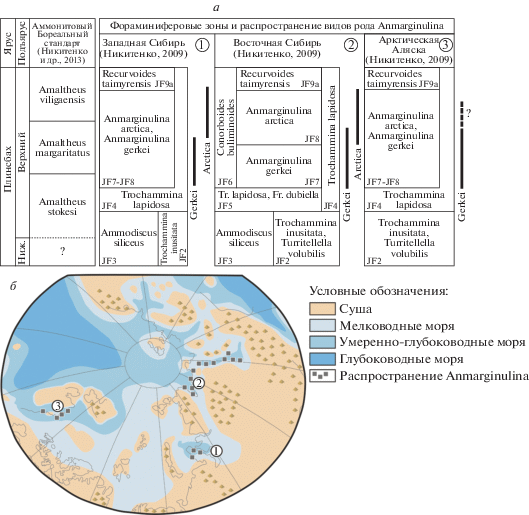
Рис. 1.
Стратиграфическое (а) и палеогеографическое (б) распространение представителей рода Anmarginulina в позднем плинсбахе.
Одна из таких групп в юре представлена видами рода Anmarginulina. Первоначально эти формы включались в состав рода Marginulina. На начальном этапе микропалеонтологических исследований юры Сибири отмечалось своеобразие гладких маргинулин с многослойной стенкой, в значительной степени объемлющими и быстро увеличивающимися в размерах, по мере роста, камерами, отсутствием ребристых форм и своеобразной “кубаревидной” формой раковины на ранних этапах развития (Герке, 1961а), что резко отличало их от типичных маргинулин. В эту группу входили новые виды: Marginulina arctica Schleifer, M. commaeformis Schleifer и M. turbiformis Schleifer. Особо отмечалась узкая стратиграфическая приуроченность этих форм только к среднему лейасу (плинсбаху) (Герке, 1961а). В конце 50-х и начале 60-х годов поступало огромное количество первых данных по микропалеонтологии мезозоя Арктики. В объемной и информативной монографии А.А. Герке (1961а) приоритет отдавался массовому изображению новых видов, была описана только часть новых видов и составлен микропалеонтологический атлас, состоящий из 122 палеонтологических таблиц с изображениями практически всех позднепалеозойских и раннемезозойских фораминифер, включая виды рода Marginulina. Монографические описания остальных основных изученных форм по техническим причинам не могли быть включены в данную монографию (Герке, 1961а) и предполагалось, что они будут приведены в последующих работах, тогда как изображения и фототаблицы уже опубликованы в данной сводке. В последующем были опубликованы монографические описания ряда видов (Герке, 1961б, 1962 и др.), ранее изображенных в этой монографии Герке (1961а). К сожалению, монографические исследования представителей сибирских Marginulina и стратиграфически важной группы видов arctica – commaeformis – turbiformis, так и остались неопубликованными, хотя были приведены детальные изображения как их внешней морфологии, так и внутреннего строения (Герке, 1961а).
В последующие годы изучение типового вида (Marginulina raphanus d’Orbigny) рода Marginulina, а также многочисленных представителей этого рода из юры разных бассейнов показало, что эти формы обладают значительным полиморфизмом, связанным как с внутривидовой (чередование разных поколений) и межвидовой изменчивостью, так и с существенной неоднородностью состава самого рода (Басов, Яковлева, 1975). Недооценка полиморфизма вела к значительным проблемам в систематике, возникали сложности в разграничении разных генераций некоторых видов родов Marginulina, Marginulinopsis, Dentalina, Nodosaria. Многие формы, относимые к маргинулинам, имели признаки, существенно отклоняющиеся от таковых у типового вида и авторского диагноза рода. Это привело к тому, что из состава рода Marginulina были обособлены формы, описанные как новые родовые таксоны, например, Marginulinita (Кузнецова, 1972). Внутри рода Marginulina были выделены четыре основные группы видов рода Marginulina, встречающиеся в юрских и меловых бассейнах бореальных регионов (Басов, Яковлева, 1975).
Одна из этих групп рода Marginulina представляет виды arctica – commaeformis – turbiformis из верхнего плинсбаха Сибири. К ним добавился вид suprajurensis из верхов средней и верхней юры. Из всех видов этой группы видов только M. suprajurensis Gerke et Scharovskaja имел опубликованное монографическое описание (Шаровская, 1961) и полностью соответствовал требованиям Международного кодекса зоологической номенклатуры (2004). По мнению исследователей, статус этой группы видов мог иметь подродовое или родовое значение (Басов, Яковлева, 1975). Однако на тот момент времени еще не сложилось устоявшейся точки зрения на уровень ранга признаков этой группы видов.
Таким образом, очевидно, что виды рода Marginulina нуждаются в серьезной ревизии. Детальное исследование выборок группы видов arctica – commaeformis – turbiformis, с учетом внутреннего строения раковин, современных представлений о диморфизме и изменчивости, из разрезов верхнего плинсбаха севера Сибири позволило обосновать обособление этой группы в новый род Anmarginulina (Никитенко, 1992, 2009; Никитенко и др., 2000; Nikitenko, Mickey, 2004, 2009). В качестве типового вида рода был предложен вид Marginulina arctica Schleifer. Для рода Anmarginulina характерны высокие, сильно объемлющие, быстро увеличивающиеся в размерах камеры, типа псевдонодозариевых, лучистое устье, слаборазвитый изогнутый отдел. Выделенный род, по-видимому, следует относить к нодозариидам.
Анализ вертикального и латерального распределения этих форм показал, что в выборках “виды” arctica, commaeformis и turbiformis практически всегда встречаются совместно в разных соотношениях. Экземляры, идентичные “виду” arctica, обладают морфологическими особенностями, характерными для мегасферических генераций: крупный пролокус (60–140 мкм), крайне слабо проявленный изогнутый отдел, хорошо развитый выпрямленный отдел (Герке, 1961а, табл. 80 , фиг. 4–6 ; Никитенко, 2009, табл. f-30, фиг. 13–17 , табл. f-31, фиг. 1–3 ). Тогда как “виды” commaeformis и turbiformis имеют изогнуто-субтреугольную форму раковины, маленький пролокус (30–56 мкм), вогнутую брюшную сторону (Герке, 1961а, табл. 78 , фиг. 2–8 ; табл. 80 , фиг. 1–3 ; Никитенко, 2009, табл. f-31, фиг. 4 ), что позволяет относить их к микросферическим особям. Таким образом, “виды” commaeformis – turbiformis и “вид” arctica являются разными генерациями одного вида. Проведенная ревизия позволила рассматривать “виды” commaeformis и turbiformis в качестве младших синонимов Anmarginulina arctica (Schleifer).
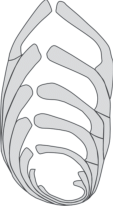
Рис. 2.
Внутреннее строение Anmarginulina arctica (Schleifer, 1961), вид сбоку, продольный срез; мегасферическая особь; Нордвикский фациальный район, п-ов Юрюнг-Тумус, обн. 5, пачка 9, сл. 1, обр. 4; верхний плинсбах, фораминиферовая зона Anmarginulina arctica JF8, аиркатская свита.
Микропалеонтологические исследования плинсбаха Сибири позволили выявить предковую форму вида Anmarginulina arctica (Schleifer), которая появляется в первой половине позднего плинсбаха (рис. 1, а). Эта форма была обособлена в самостоятельный вид A. gerkei Nikitenko, который был также только изображен в публикациях (Никитенко, 1992, 2009).
Анализ вертикального распределения этих видов в разных бассейнах показал, что они имеют четкую стратиграфическую приуроченность (рис. 1, а). Anmarginulina gerkei Nikitenko распространен в верхней части аммонитовой зоны Amaltheus stokesi и в большей части зоны Amaltheus margaritatus. Тогда как вид Anmarginulina arctica (Schleifer) встречается в верхней части аммонитовой зоны Amaltheus margaritatus и низах зоны Amaltheus viligaensis. Позднеплинсбахские представители Anmarginulina имеют широкое географическое распространение и являются маркерами для межрегиональных корреляций, их находки отмечаются в разных Арктических бассейнах, на севере Западной и Восточной Сибири, Арктической Аляске (рис. 1, б). Все это обусловило широкое использование этих видов в стратиграфии (Стратиграфия…, 1976; Решения…, 1981; Шурыгин и др., 2000; Решение, 2004; Никитенко, 2009 и др.).
Относительно высокие темпы эволюции, частота встречаемости, широкое географическое распространение позволили рассматривать виды Anmarginulina gerkei Nikitenko и A. arctica (Schleifer) в качестве видов-индексов верхнеплинсбахских фораминиферовых зон комплексного обоснования JF7 и JF8 (рис. 1, а) (Никитенко, 1992, 2009). На “закрытых” территориях (Западная Сибирь, Северная Аляска) столь дробное расчленение этой части разреза пока еще вряд ли возможно, из-за редкости отбора керна и, иногда, специфических фаций. Тем не менее, на севере Западной Сибири и на севере Аляски удается распознавать нерасчлененные слои с Anmarginulina arctica, A. gerkei JF7-JF8 (рис. 1).
В последние годы активно создаются электронные базы данных морских организмов, например, такие как World Register of Marine Species (WoRMS) или World Foraminifera Database. В эти базы включены род Anmarginulina и входящие в него виды (Hayward et al., 2018, 2019). За долгую историю исследований самостоятельность этих таксонов, высокий биостратиграфический потенциал и широкое географическое распространение признается специалистами по раннеюрским фораминиферам. Однако отсутствие опубликованных монографических описаний этих видов, особенно A. arctica (Schleifer), выбранного в качестве типового вида рода Anmarginulina, согласно Международному кодексу зоологической номенклатуры (2004) позволяет рассматривать эти таксоны только в категории nomen nudum. Таким образом, несмотря на многолетнюю историю использования этих таксонов в таксономии, биостратиграфии, межрегиональных корреляциях, палеобиогеографии, полноценное их использование невозможно без публикации подробного монографического описания видов. Для этого необходима валидизация видов и рода Anmarginulina путем проведения монографического исследования и публикации первого подробного описания этих таксонов.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Материалом для монографических исследований послужили коллекции фораминифер из разрезов верхнего плинсбаха арктических регионов Западной (п-ов Ямал, Усть-Енисейский район) и Восточной Сибири (Восточный Таймыр, п-ов Юрюнг-Тумус, Анабарский залив, нижнее течение бассейна р. Оленек, междуречья рр. Оленек и Лена, Приверхоянского прогиба, Вилюйского бассейна). Для сравнения привлекались топотипические коллекции фораминифер (НИИГА – ВНИИОкеангеология) из разрезов, вскрытых скважинами в Нордвикском р-не, побережье Хатангского залива (сопка Кожевникова, р. Тигян). Также привлекались результаты микропалеонтологических исследований разрезов верхнего плинсбаха, вскрытого скважинами в Арктической Аляске (рис. 1) (Никитенко, 2009).
Исследуемые представители Anmarginulina в выборках обычно представлены 3–10 экз., реже 10–15 экз. Раковины фораминифер в изучаемых и сравниваемых выборках разных местонахождений имеют хорошую сохранность, следы сортировки или транспортировки во многих случаях отсутствуют или слабо представлены. В выборках также обычно многочисленны ювенильные особи. Все это позволяет предположить, что исследуемые выборки сделаны из ископаемых автохтонных или незначительно перемещенных захоронений и происходят из ископаемых популяций. Морфология раковин одного и того же вида фораминифер может значительно различаться, в зависимости от принадлежности к микро- или мегасферической генерации. Поэтому анализируемые выборки видов рода Anmarginulina оценивались с учетом современных представлений о диморфизме фораминифер. Это позволило провести ревизию и синонимизацию ряда таксонов.
Фораминиферы изучались под стереомикроскопом Discovery V20 (Carl Zeiss), микрофотографирование проводилось при помощи электронного сканирующего микроскопа Zeiss LEO-1430 vp и на световом микроскопе Discovery V20 с использованием фотокамеры AxioCamMRc5 (Carl Zeiss Microscopy).
При описании видов использована система высших таксонов фораминифер, предложенная A. Лебликом и Х. Таппан (Loeblich, Tappan, 1987, 1992). При исследовании видов и их замеров применялись следующие сокращения: dp – диаметр начальной камеры, L – длина раковины, B – ширина раковины, L/B – степень удлиненности раковины, N – общее количество камер. Коллекция хранится в лаб. микропалеонтологии и ЦКП “Коллекция ГЕОХРОН” Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН.
Автор искренне признателен В.С. Вишневской за ценные комментарии и замечания, способствовавшие улучшению статьи. Работа выполнена по планам исследований ФНИ № 0331-2019-0005, при поддержке проектов РНФ № 19-17-00091 и РФФИ № 18-05-70035.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
СЕМЕЙСТВО NODOSARIIDAE EHRENBERG, 1838
ПОДСЕМЕЙСТВО NODOSARIINAE EHRENBERG, 1838
Род Anmarginulina Nikitenko, 2000
Marginulina (part.): Герке, 1961а, табл. 78 , фиг. 2–8 ; табл. 80 , фиг. 1–6 ; табл. 81 , фиг. 1, 2; табл. 82 , фиг. 1–7 ; табл. 84 , фиг. 7, 8 ; Басов, Яковлева, 1975, с. 87–91 (part.).
Anmarginulina: Никитенко, 1992, с. 10, табл. 2, фиг. 9 ; Никитенко и др., 2000, с. 95, рис. 3 , табл. II , фиг. 15 ; Nikitenko, Mickey, 2004, fig. 6 i–k ; Никитенко, 2009, табл. f-30, фиг. 10–17 ; табл. f-31, фиг. 1–10 .
Типовой вид – Marginulina arctica Schleifer, 1961; верхний плинсбах; север Средней Сибири, Нордвикский район.
Диагноз. Раковина билатерально-симметричная, на ранней стадии в разной степени изогнутая на брюшную сторону, поздний отдел обычно слабоизогнутый или прямолинейный; сечение округло-овальное. Камеры сильно объемлют друг друга и быстро увеличиваются в диаметре (рис. 2). Септальные швы поверхностные, наклонные. Поверхность камер гладкая; устье лучистое, смещенное к спинному краю. Стенка раковины толстая, многослойная.
Видовой состав. A. mirifica (Gerke) (карний Сибири), A. gerkei Nikitenko (средняя часть верхнего плинсбаха Сибири и Арктической Аляски, рис. 1), A. arctica (Schleifer) (верхняя часть верхнего плинсбаха Сибири, рис. 1), A. suprajurensis (Gerke et Scharovskaja) (бат – основание верхнего оксфорда Сибири и Арктической Аляски) (Герке, 1961а; Шаровская, 1961; Никитенко, 2009).
Сравнение. От наиболее близкого рода Pandaglandulina Loeblich et Tappan, 1955 отличается сильным изгибом сериальной оси, расположением устья, смещенным к спинному краю, и не параллельными боковыми сторонами. От Marginulina d’Orbigny, 1826 отличается сильно объемлющими, быстро увеличивающимися в диаметре камерами.
Замечания. Изображенные и описанные виды (Герке, 1961а; Шаровская, 1961) относились ранее к роду Marginulina, но типовой вид рода – Marginulina raphanus d’Orbigny из плиоцена характеризуется практически не объемлющими камерами, постепенно и равномерно увеличивающимися как в диаметре, так и в высоту.
Возможно, к роду Anmarginulina следует относить кайнозойский вид Marginulina glabra d’Orbigny (=M. similis d’Orbigny), характеризующийся слабым развитием спирали и, возможно, сильно объемлющими камерами (Басов, Яковлева, 1975). Однако последний признак иллюстрируется схематичными рисунками, или фотографиями раковины без деталей внутреннего строения. Поэтому включать этот вид в состав рода Anmarginulina без исследований и ревизии оригинального материала представляется пока преждевременным.
Anmarginulina gerkei Nikitenko, 1992
Табл. II , фиг. 1–7 (см. вклейку)
Marginulina terquemi: Герке, 1961а, табл. 81 , фиг. 1, 2; табл. 82 , фиг. 1–7 ; табл. 84 , фиг. 7, 8 .
Anmarginulina gerkei: Никитенко, 1992, с. 10, табл. 2, фиг. 9 ; 2009, табл. f-30, фиг. 10–12 ; табл. f-31, фиг. 5 .
Голотип – ИНГГ СО РАН, № 1048/101; западный берег Анабарского залива, обн. 12, сл. 50, обр. 1; верхний плинсбах, верхи аммонитовой зоны Amaltheus stokesi, фораминиферовая зона Anmarginulina gerkei.
Описание. Раковина от маленькой до средней по размерам, удлиненная, в очертании вытянуто-эллипсоидная, каплевидная, толстая, в начальной части быстро расширяющаяся, постепенно в поздней, почти прямая или слабоизогнутая в начальной части на брюшную сторону. Брюшной край в начальной части слабо изогнут, в поздней части прямой, широко округлый. Спинной край прямой, широко округлый. Поперечное сечение выпрямленной части округлое. Начальная часть раковины приостренно-округлое. Раковина состоит из 5–9 камер. Начальная камера от сферической до эллипсоидальной, 28–60 мкм в диаметре. Камеры в начальной части сильно объемлющие, быстро увеличивающиеся в диаметре. Последующие камеры более высокие, менее объемлющие, рост в диаметре практически прекращается и увеличивается длина камер. Камеры практически невыпуклые, лишь у взрослых экземпляров последние 2–3 слабовыпуклые. Септальные швы поверхностные, прямые. Устьевая поверхность последней камеры округлая или округло-треугольная. Устье лучистое, смещенное к спинному краю последней камеры. Поверхность раковины гладкая. Стенка раковины толстая, многослойная.
Размеры в мм:
| Экз. № | dp | L | B | H | L/B | N |
|---|---|---|---|---|---|---|
| Мегасферическая генерация | ||||||
| 1048/101, голотип | 0.042 | 1.02 | 0.32 | 0.32 | 3.18 | 9 |
| 1048/102 | 0.042 | 0.97 | 0.37 | 0.34 | 2.62 | 8 |
| 1048/190 | 0.042 | 1.03 | 0.42 | 0.42 | 2.46 | 6 |
| 1048/185 | 0.056 | 0.69 | 0.31 | 0.26 | 2.22 | 6 |
| 1048/193 | 0.056 | 1.05 | 0.39 | 0.35 | 2.69 | 8 |
| 1048/194 | 0.056 | 0.73 | 0.21 | 0.21 | 3.46 | 6 |
| 1048/195 | 0.056 | 0.63 | 0.28 | 0.28 | 2.25 | 5 |
| 1048/203 | 0.07 | 1.02 | 0.39 | 0.38 | 2.61 | 7 |
| Пределы изменений | 0.042–0.07 | 0.52–1.05 | 0.2–0.43 | 0.21–0.42 | 2.22–3.46 | 3–9 |
| Микросферическая генерация | ||||||
| 1048/200 | 0.02 | 0.4 | 0.23 | 0.18 | 1.7 | 5 |
| 1048/103 | 0.042 | 0.55 | 0.19 | 0.18 | 2.89 | 7 |
| 1048/196 | 0.028 | 0.7 | 0.25 | 0.25 | 2.8 | 5 |
| 1048/191 | 0.04 | 0.92 | 0.28 | 0.28 | 3.3 | 7 |
| Пределы изменений | 0.02–0.042 | 0.3–1.0 | 0.19–0.28 | 0.18–0.28 | 1.7–3.3 | 3–8 |
Изменчивость. Проявлена в большей или меньшей изогнутости начального отдела и степени перекрывания камер. Микросферические формы имеют быстро расширяющиеся в ширину камеры по мере роста на начальных стадиях, что придает каплевидную форму раковине в очертании. Раковина в начальной части может быть приостренной, со слаборазвитым шипом в основании; диаметр начальной камеры составляет 20–40 мкм. Мегасферические экземпляры имеют сильно удлиненную вытянутую форму раковины. Раковина практически однорядная, в основном выпрямленная, первые две–три камеры нарастают по оси, очень слабоизогнутой на брюшную сторону. В основании раковины округлая начальная камера диаметром 42–80 мкм.
Сравнение. От A. mirifica (Герке, 1961а) из верхнего триаса севера Сибири отличается более крупными размерами, невыпуклыми камерами, прямыми поверхностными швами. От A. arctica (Герке, 1961а, табл. 78 , фиг. 2–8 ; табл. 80 , фиг. 1–6 ; Nikitenko, Mickey, 2004; Никитенко, 2009; настоящая работа) из верхнего плинсбаха севера Сибири отличается более крупной и вытянутой раковиной (L/B = 2.22–3.46 против 1.17–2.5), большим количеством камер в выпрямленной части раковины, менее объемлющими и более высокими камерами в поздней части раковины.
Распространение. Верхний плинсбах, верхняя часть аммонитовой зоны Amaltheus stokesi – зона Amaltheus margaritatus; фораминиферовая зона Anmarginulina gerkei JF7 – низы зоны Anmarginulina arctica JF8; север Западной и Восточной Сибири, Арктическая Аляска (рис. 1).
Материал. Более 70 раковин различной сохранности из разрезов западного берега Анабарского залива, обн.12 (Никитенко, 2009); п-ова Юрюнг-Тумус, обн. 4а, обн. 1, обн. 2, обн. 8 (Никитенко, 2009); Восточный Таймыр (м. Цветкова) обн. 7 (Никитенко, 2009); разрезы естественных выходов и скважин бассейна р. Оленек, междуречья рр. Оленек и Лена, Приверхоянский прогиб, Вилюйский бассейн; разрезы, вскрытые скважинами в арктической части Западной Сибири и Арктической Аляски (Никитенко, 2009).
Anmarginulina arctica (Schleifer, 1961)
Табл. II , фиг. 8–12 ; табл. III , фиг. 1–9 (см. вклейку)
Marginulina arctica: Герке, 1961а, табл. 80 , фиг. 4–6 .
Marginulina commaeformis: Герке, 1961а, табл. 80 , фиг. 1–3 .
Marginulina turbiformis: Герке, 1961а, табл. 78 , фиг. 2–8 .
Anmarginulina arctica: Nikitenko, Mickey, 2004, рис. 6 i–k ; Никитенко, 2009, табл. f-30, фиг. 13–17 ; табл. f-31, фиг. 1–4 .
Голотип – изображен в работе А.А. Герке (1961а), табл. 80 , фиг. 4 ; север Средней Сибири, Нордвикский район, сопка Кожевникова, скв. К-115, глубина 305–306 м; средний лейас.
Описание (рис. 2). Раковина от маленькой до средней по размерам, в очертании от изогнуто-субтреугольной, до неправильно-эллипсоидальной, короткая, толстая, быстро расширяющаяся по мере роста, изогнутая в начальной части на брюшную сторону (рис. 2). Брюшной край прямой или слабовогнутый, широко-округленный. Спинной край прямой или слабоизогнутый, от широко-округлого до притупленно-угловатого. Поперечное сечение выпрямленной части округло-овальное. Начальная часть раковин варьирует от округлой до приостренной с коротким шипиком. Боковые стороны не параллельны друг другу, сходящиеся у основания. Раковина состоит из 4–9 камер; в изогнутой части обычно 3–4 камеры, в выпрямленной части 1–5. Начальная камера от сферической до эллипсоидальной, 42–140 мкм в диаметре. Последующие камеры сильно объемлют предыдущие, быстро увеличиваются в диаметре. Камеры слабовыпуклые, лишь у взрослых экземпляров последняя камера заметно выпуклая. Септальные швы поверхностные, прямые, в начальной части скошенные. Устьевая поверхность последней камеры округло-треугольная, устье лучистое, расположено у спинного края последней камеры. Поверхность раковины гладкая. Стенка раковины толстая, многослойная, радиально-лучистая.
Размеры в мм:
| Экз. № | dp | L | B | H | L/B | N |
|---|---|---|---|---|---|---|
| Мегасферическая генерация | ||||||
| 1048/99 | 0.08 | 0.35 | 0.21 | 0.22 | 1.66 | 4 |
| 1048/183 | 0.09 | 0.98 | 0.47 | 0.43 | 2.05 | 9 |
| 1048/96 | 0.084 | 0.7 | 0.42 | 0.42 | 1.66 | 8 |
| 1048/95 | 0.09 | 0.75 | 0.41 | 0.41 | 1.82 | 6 |
| 1048/185 | 0.11 | 0.57 | 0.39 | 0.40 | 1.46 | 5 |
| 1048/186 | 0.09 | 0.45 | 0.28 | 0.29 | 1.6 | 4 |
| 1048/187 | 0.084 | 0.39 | 0.22 | 0.22 | 1.75 | 4 |
| 1048/188 | 0.11 | 1.05 | 0.42 | 0.43 | 2.5 | 7 |
| 1048/98 | 0.06 | 0.28 | 0.24 | 0.21 | 1.17 | 4 |
| Пределы изменений | 0.06–0.14 | 0.35–1.05 | 0.21–0.47 | 0.22–0.43 | 1.46–2.5 | 3–9 |
| Микросферическая генерация | ||||||
| 1048/184 | 0.042 | 0.43 | 0.196 | 0.18 | 2.21 | 4 |
| 1048/97 | 0.056 | 0.85 | 0.28 | 0.28 | 1.96 | 6 |
| 1048/100 | 0.042 | 0.53 | 0.26 | 0.24 | 2.03 | 5 |
| 1048/189 | 0.07 | 0.95 | 0.56 | - | 1.7 | 7 |
| Пределы изменений | 0.042–0.07 | 0.28–0.95 | 0.196-0.56 | 0.18–0.28 | 1.17–2.21 | 3–8 |
Изменчивость. Микросферические особи обладают изогнуто-субтреугольной формой раковины в очертании. Раковина в начальной части приостренная, с шипиком в основании; диаметр начальной камеры 30–56 мкм; брюшная сторона вогнутая. Мегасферические экземпляры имеют неправильно эллипсоидную форму раковины. Изогнутый отдел развит очень слабо, часто практически прямой. Раковина в основании округлая, диаметр начальной камеры 60–140 мкм, брюшной и спинной края обычно прямые или слабоизогнутые; начальная камера не выдается на поверхности раковины.
Сравнение. От A. mirifica (Герке, 1961а) из верхнего триаса севера Сибири отличается большими размерами, более толстой и короткой раковиной, более объемлющими камерами, положением устья, расположенным у спинного края последней камеры.
Распространение. Верхний плинсбах, верхняя часть аммонитовой зоны Amaltheus margaritatus – нижняя половина зоны Amaltheus viligaensis; фораминиферовая зона Anmarginulina arctica JF8; север Западной и Восточной Сибири (рис. 1).
Материал. Более 90 раковин хорошей и удовлетворительной сохранности из разрезов западного берега Анабарского залива, обн. 12 (Никитенко, 2009); п-ова Юрюнг-Тумус, пачка 9, обн. 8, обн. 5 (Никитенко, 2009); разрезы скважин Нордвикского р-на (Герке, 1961); р. Анабар, обн. 3 (Никитенко, 2009); Восточный Таймыр (м. Цветкова) обн. 7 (Никитенко, 2009); разрезы естественных выходов и скважин бассейна р. Оленек, междуречья рр. Оленек и Лена, Приверхоянский прогиб, Вилюйская синеклиза; разрезы, вскрытые скважинами в арктической части Западной Сибири (Никитенко, 2009).
Список литературы
Басов В.А., Яковлева С.П. Группа Marginulina–Marginulinopsis–Saracenaria–Saracenella // Проблемы систематики спиральных нодозариид. Вильнюс: Минтис, 1975. С. 83–102.
Герке А.А. Фораминиферы пермских, триасовых и лейасовых отложений нефтеносных районов севера Центральной Сибири. Л.: Гостоптехиздат, 1961а. 579 с. (Тр. НИИГА. Т. 120).
Герке А.А. Ректогландулины из пермских и нижнемезозойских отложений севера Центральной Сибири // Сб. статей по палеонтологии и биостратиграфии. Вып. 1. Л.: Изд-во НИИГА, 1961б. С. 5–34.
Герке А.А. Фрондикуларии из пермских, триасовых и лейасовых отложений севера Центральной Сибири // Проблемы нефтегазоносности Советской Арктики. Палеонтология и биостратиграфия. Л.: Гостоптехиздат, 1962. С.97–175.
Кузнецова К.И. Новый род Marginulinita К. Kuznetsova и некоторые его новые виды из поздней юры Русской платформы // Вопр. микропалеонтол. 1972. Вып. 15. С. 91–102.
Международный кодекс зоологической номенклатуры. Издание четвертое. М.: Т-во научн. изданий КМК, 2004. 223 с.
Никитенко Б.Л. Зональная шкала нижней и средней юры на севере Сибири по фораминиферам // Геол. и геофизика. 1992. № 1. С. 3–14.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Никитенко Б.Л., Сапьяник В.В., Глинских Л.А. Ранне-среднеюрские комплексы фораминифер и остракод северной и южной окраин Западно-Сибирской равнины // Новости палеонтологии и стратиграфии. Прилож. к журн. Геол. и геофизика. 2000. Т. 41. № 2–3. С. 87–109.
Решение 6-го межведомственного стратиграфического совещания по рассмотрению и принятию уточненных стратиграфических схем мезозойских отложений Западной Сибири, Новосибирск, 2003. Региональные стратиграфические схемы триасовых и юрских отложений Западной Сибири. Новосибирск: СНИИГГиМС, 2004. 113 с.
Решения 3-го Межведомственного регионального стратиграфического совещания по мезозою и кайнозою Средней Сибири. Новосибирск: ИГиГ СО АН СССР, 1981. 91 с.
Стратиграфия юрской системы севера СССР. М.: Наука, 1976. 436 с.
Шаровская Н.В. Некоторые виды фораминифер из верхнеюрских отложений Нордвикского района // Сб. статей по палеонтологии и биостратиграфии. Вып. 27. Л.: Изд-во НИИГА, 1961. С. 17–79.
Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П. и др. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск: Изд-во СО РАН, филиал “Гео”, 2000. 480 с.
Hayward B.W., Le Coze F., Gross O. World Foraminifera Database. Anmarginulina Nikitenko, 2000. 2018. http:// www.marinespecies.org/foraminifera…/aphia.php?p=taxdetails&id=1047733.
Hayward B.W., Le Coze F., Vachard D., Gross O. World Foraminifera Database. Anmarginulina Nikitenko, 2000. 2019. http://www.marinespecies.org/aphia.php?p=taxdetails&id= 1047733.
Loeblich A., Tappan H. Foraminiferal genera and their classification. N.Y., 1987. V. 1, 2. 1182 p.
Loeblich A.R., Tappan, H. Present status of foraminiferal classification, in Studies in Benthic Foraminifera // Proc. of the Fourth Intern. Symp. on Benthic Foraminifera, Sendai, 1990 (Benthos’90). Tokyo: Tokai Univ. Press, 1992. P. 93–102.
Nikitenko B.L., Mickey M.B. Foraminifera and ostracodes across the Pliensbachian-Toarcian boundary in the Arctic Realm // Geol. Soc. Spec. Publ. 2004. V. 230. P. 137–173.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал