

Палеонтологический журнал, 2020, № 3, стр. 87-102
Самый молодой триасовый комплекс наземных позвоночных в России: состав и датировка
В. П. Твердохлебов a, *, А. Г. Сенников b, c, **, И. В. Новиков b, c, ***, Н. В. Ильина d, ****
a Саратовский государственный университет
Саратов, Россия
b Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
c Казанский федеральный университет
Казань, Россия
d Институт геологии Коми НЦ УрО РАН
Сыктывкар, Россия
* E-mail: galtver@yandex.ru
** E-mail: sennikov@paleo.ru
*** E-mail: inovik@paleo.ru
**** E-mail: ilyina@geo.komisc.ru
Поступила в редакцию 06.06.2019
После доработки 27.06.2019
Принята к публикации 31.07.2019
Аннотация
Самый молодой фаунистический комплекс наземных позвоночных триаса в России приурочен к верхам букобайской свиты (средний триас) Южного Приуралья в местонахождении Лысов (Оренбургская область). Кроме тетрапод и рыб, из этого местонахождения известны двустворчатые моллюски, разнообразная макрофлора и богатый палинокомплекс. Изучение последнего позволило уверенно подтвердить датировку вмещающих отложений ладинским веком. Букобайское время представляет собой особый (ладинский) этап развития фауны и флоры на территории Южного Приуралья.
ВВЕДЕНИЕ
На территории Восточной Европы и Южного Приуралья триасовые континентальные отложения предоставлены наиболее полно и охарактеризованы рядом последовательных и преемственных фаун тетрапод (Шишкин и др., 1995). Наиболее поздняя из них, так называемая “мастодонзавровая” фауна, известна из Южного (букобайская свита) и Северного (надкраснокаменская свита) Приуралья, а также в индерской свите Прикаспия. В Южном Приуралье эта фауна включает в себя из темноспондильных лабиринтодонтов – гигантских капитозавроидов “Mastodonsaurus” torvus Konzhukova11, Cyclotosauridae gen.indet., трематозавроида Bukobaja enigmatica Otschev, плагиозавра Plagiosternum danilovi Shishkin, а среди рептилий – пролацертилию Malutinisuchus gratus Otschev, эритрозухида Chalishevia cothurnata Otschev, рауизухид Energosuchus garjainovi Otschev и Jushatyria vjushkovi Sennikov, неопределимых до рода териодонтов, каннемейероидных дицинодонтов Elatosaurus facetus Kalandadze и Elephantosaurus jachimovitschi Vjuschkov. В Северном Приуралье, где, кроме континентальных, имеются и прибрежноморские отложения, в “мастодонзавровой” фауне представлены Bukobaja (?) sp., реликтовые антракозавры-хрониозухии Synesuchus muravjevi Novikov et Shishkin, эозауроптеригия Pistosaurus (?) sp., пролацертилия Malutinisuchus gratus, рауизухид Energosuchus garjainovi и каннемейероидные дицинодонты Planitorostris pechoriensis Surkov. В Прикаспии из индерской свиты прибрежноморского генезиса известны “Mastodonsaurus” torvus, Bukobaja sp., плагиозавр Plagioscutum caspiense Shishkin, неопределимые до рода проколофоны, рауизухиды, эритрозухиды и каннемейероидные дицинодонты. Возраст букобайской фауны тетрапод оценивается как ладинский (Шишкин и др., 1995).
В то же время букобайская фауна, отражающая столь важный этап эволюции тетрапод, остается одной из наименее изученных, так как она известна из ограниченного числа местонахождений, на которых не производились систематические раскопки. В Южном Приуралье это местонахождения Букобай I, II, III, V, VII и Ново-Александровка (верхняя точка) в бассейне р. Бердянка, Донгуз XI в бассейне р. Донгуз, Колтаево III в бассейне р. Бол. Юшатырь. Все эти местонахождения позвоночных приурочены к нижним циклитам букобайской свиты Южного Приуралья (Шишкин и др., 1995). В настоящей статье впервые приведено подробное описание этого единственного в верхах букобайской свиты комплекса тетрапод и рыб.
МЕСТОНАХОЖДЕНИЕ ЛЫСОВ: ГЕОЛОГИЧЕСКОЕ СТРОЕНИЕ И ГЕНЕЗИС
Местонахождение Лысов было открыто В.П. Твердохлебовым в 1964 г. во время геологической съемки около хут. Лысов в Оренбургском районе (рис. 1). Здесь, вдоль оврага Большой (приток овр. Елшанка), им был описан протяженный, но слабо обнаженный разрез верхов букобайской свиты среднего триаса (Твердохлебов, 1967; Шишкин и др., 1995; Tverdochlebov, 2004). На нескольких уровнях по всему разрезу были обнаружены кости позвоночных, двустворчатые моллюски, макрофлора хорошей сохранности, взяты пробы на миоспоры.
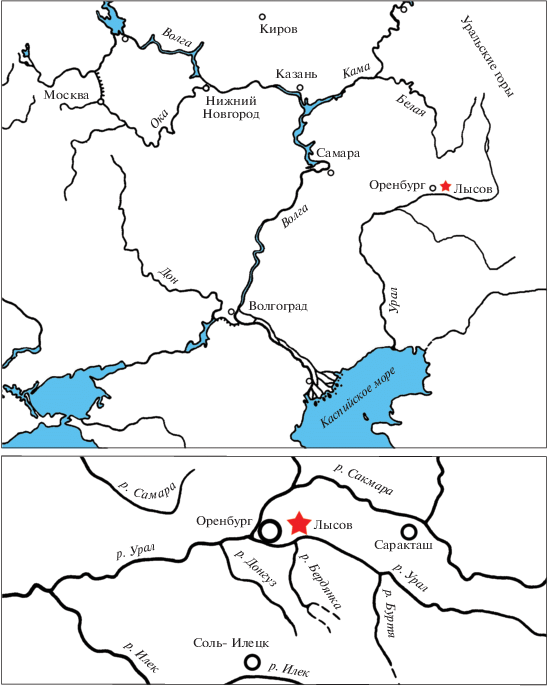
При изучении местонахождения в середине девяностых годов ХХ в. М.В. Сурков (Саратовский государственный ун-т) обнаружил в нижней части разреза этого местонахождения скопление костей позвоночных и собрал некоторое количество образцов, хранящихся ныне в СГУ. В 1997 г. в рамках совместной экспедиции ПИН РАН, СГУ и Музея Северной Аризоны, в которой приняли участие А.Г. Сенников, И.В. Новиков, А.Г. Власов, М.В. Сурков и М. Моралес, был обследован разрез местонахождения, произведены пробные раскопки на точке с остатками позвоночных в нижней части разреза, давшие новые материалы, хранящиеся в ПИН РАН. Также были взяты пробы на палинологический анализ.
Ниже приводится общий разрез местонахождения Лысов по В.П. Твердохлебову, снизу вверх (рис. 2):
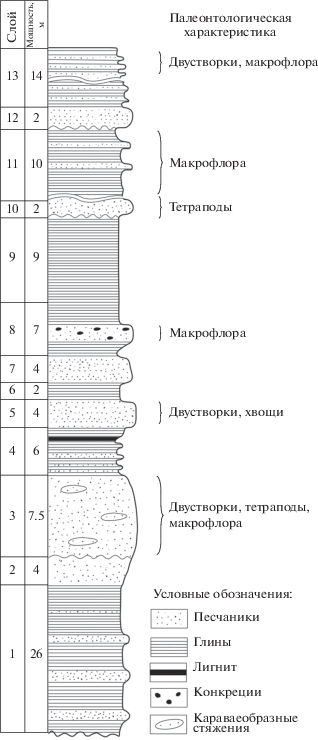
Слои 1. Глины пестрые с преобладанием красно-бурых тонов с прослоями серых песчаников, от тонко- до среднезернистых, мощностью до 1 м. Видимая мощность 26.0 м.
Слой 2. Песчаники серые тонкозернистые глинистые. Мощность 4.0 м.
Слой 3. Песчаники серые, табачно-серые мелкозернистые, косослоистые с караваеобразными стяжениями, содержащие остатки двустворчатых моллюсков. В слое найдены также кости тетрапод и остатки макрофлоры. Нижняя граница неровная, с размывом. Мощность 7.5 м.
Слой 4. Пачка переслаивания сероцветных глин и песчаников. В верхах линзообразный прослой лигнита мощностью 0.3 м. Мощность 6.0 м.
Слой 5. Песчаники серые мелкозернистые косослоистые с вертикально захороненными стволами хвощей и редкими раковинами двустворчатых моллюсков. Нижняя граница неровная, с размывом. Мощность 4.0 м.
Слой 6. Глины зеленовато-серые. Мощность 2.0 м.
Слой 7. Песчаники серые среднезернистые косослоистые. Нижняя граница неровная, с размывом. Мощность 4.0 м.
Слои 8. Пачка глин желтовато-серых с 2-х метровым прослоем серых тонкозернистых песчаников с карбонатно-глинистыми конкрециями, содержащими остатки макрофлоры хорошей сохранности. Мощность 7.0 м.
Слой 9. Глины желтовато-серые с розовыми и красными пятнами. Мощность 9.0 м.
Слой 10. Песчаники желто-серые средне- и крупнозернистые косослоистые. В слое найдены кости тетрапод. Нижняя граница неровная, с размывом. Видимая мощность 2.0 м.
Выше – перерыв в обнаженности 21 м.
Слой 11. Глины зеленовато-серые с прослоями (до 1м) песчаников зеленовато-серых мелкозернистых с остатками макрофлоры хорошей сохранности. Видимая мощность 10.0 м.
Слой 12. Песчаники желтовато-серые среднезернистые косослоистые. Нижняя граница неровная, с размывом. Видимая мощность 2.0 м.
Слои 13. Пачка переслаивания сероцветных глин и песчаников с преобладанием глин. В верхней части слоя 13 найдены остатки двустворчатых моллюсков и макрофлоры хорошей сохранности. В средней части – перерыв обнаженности 7 м. Видимая мощность 14.0 м.
Условия накопления вмещающих отложений местонахождения Лысов типичны для букобайской свиты Южного Приуралья (Шишкин и др., 1995; Tverdokhlebov, 2004). Оно происходило в обширных синклинальных просадочных структурах, представлявших собой в ландшафте полузамкнутые озерно-болотные и озерно-речные равнины. В целом, букобайские отложения имеют циклическое строение и представлены, как и в местонахождении Лысов, сероцветными песчаниками, блеклыми пестроцветными алевритами и глинами с многочисленными растительными остатками, прослоями растительного детрита и лигнитов. Это отражает произошедшую к букобайскому веку общую гумидизацию климата (лишь с некоторыми проявлениями сезонности), что привело к формированию обширных водных бассейнов, биотопов с обилием растительности и богатой кормовой базой, в которых создавались благоприятные условия для развития разнообразной фауны водных и наземных позвоночных. В это время впервые с начала триаса в данном регионе появились условия для угленакопления.
ФАУНА ПОЗВОНОЧНЫХ МЕСТОНАХОЖДЕНИЯ ЛЫСОВ
Костеносная линза в нижней части разреза местонахождения Лысов, раскопанная в 1997 г., содержала кости позвоночных, раковины двустворчатых моллюсков и макрофлору, и была приурочена к линзовидному прослою темных плотных коричнево-серых глин, примерно 1 м шириной, залегающему в слое серых мелкозернистых косослоистых песчаников (слой 3). В глинах встречаются также раковины крупных двустворчатых моллюсков и многочисленные растительные остатки, ожелезненные и обугленные стволы хвощей, отпечатки листьев, обугленные куски веток и древесины, черный растительный детрит, маломощные линзочки угля. Изолированные, часто фрагментарные кости позвоночных преимущественно крупные, очень плотные, черного цвета, вероятно, сильно ожелезненные, заключены в исключительно плотные и прочные слоистые коричневые глинисто-сидеритовые конкреции. Во время захоронения илистый осадок был, очевидно, очень мягким и пластичным, так как послойно облекал захораниваемые остатки. Для этой костеносной линзы можно предположить старичное происхождение. За пределами линзы вмещающая порода становится более светлой, песчанистой, без конкреций с костями, но с раковинами мелких двустворчатых моллюсков. В ней содержится меньшее количество детрита, нет кусков древесины и веток, но присутствуют отпечатки растений. В табачно-серых мелкозернистых песчаниках слоя 3 отмечены единичные находки костей без конкреций. Эти кости имеют иную сохранность, чем в основной части костеносной глинистой линзы, – светлые, коричневато-желтые, почти не ожелезненные, менее плотные и массивные, с лучше сохранившейся структурой поверхности. К таким образцам относится фрагмент нижней челюсти лабиринтодонта (ритидостеида). Для слоя песчаника наиболее вероятен прибрежно-отмельный генезис. Интенсивность водных потоков, переносивших органические остатки, была, очевидно, незначительна, так как кости, как и остатки растений, почти не окатаны.
Из местонахождения Лысов определены двоякодышащие рыбы Ceratodus bukobajensis Minich (Tverdokhlebov, 2004), капитозавроид “Mastodonsaurus” sp. (рис. 3, а, б), неопределимые до рода темноспондилы-ритидостеиды (рис. 3, и) и крупные каннемейероидные дицинодонты (Kannemeyeroidea gen. indet.). Комплекс позвоночных в целом соответствует известному из низов букобайской свиты. Особо следует отметить находку фрагмента остатков ритидостеида, первую для этой группы как в среднем триасе Южного Приуралья, так и для этого интервала времени вообще. Это свидетельствет о том, что представители семейства Rhytidosteidae, наиболее поздние находки которых до сих пор происходили из верхов нижнего триаса, еще продолжали существовать во второй половине среднего триаса, в ладинском веке.
Рис. 3.
Кости тетрапод из слоя 3 местонахождения Лысов: а – экз. ПИН, № 5655/1, левая заднетеменная кость “Mastodonsaurus” sp. с дорсальной стороны, б – экз. ПИН, № 5655/2, позвонок “Mastodonsaurus” sp. с краниальной стороны, в – экз. ПИН, № 5655/3, отпечаток фрагмента правой ветви нижней челюсти Rhytidosteidae gen. indet. на породе с латеральной стороны.

МАКРОФЛОРА МЕСТОНАХОЖДЕНИЯ ЛЫСОВ
Макрофлора из местонахождения Лысов (колл. ГИН № 3980, сборы В.П. Твердохлебова) была описана И.А. Добрускиной (1980, 1982). Комплекс макрофлоры включает остатки членистостебельных Equisetites arenaceus (Jaeger) Schenk, папоротников Cladophlebis shensiensis P’an, Danaeopsis rarinervis Turutanova-Ketova, пельтаспермов Lepidopteris microcellularis Dobruskina, гинкгофитов Sphenobaiera granulifer Sixtel, Glossophyllum sp., цикадофитов Apoldia surakaica (Prinada et Brick) Dobruskina и наиболее вероятно указывает на ладинский возраст местонахождения Лысов (Добрускина, 1982; Шишкин и др., 1995; Tverdokhlebov, 2004).
В средней части разреза (слой 5) В.П. Твердохлебов обнаружил захоронение зарослей гигантских членистостебельных Equisetites arenaceus (стволы и корневища) в прижизненном положении (Tverdochlebov, 2004). Десятки стволов этих древовидных хвощей расположены в обнажении вертикально, местами очень близко, почти вплотную друг к другу, образуя густые заросли (рис. 4). От одного корневища отходят до 10 стволов. Стволы хвощей имеют почти одинаковый диаметр – от 10 до 15 см, в высоту (в сохранившейся части) до 2–2.5 м, почти прямые, реже слабо изогнуты или разветвляются, заполнены плотной серой глинисто-алевритистой породой. Сохранились лишь нижние части стволов, верхушки часто отсутствуют (вероятно, обломаны). Основания стволов и корневища уходят в нижележащий слой темно-серых глин и алевритов, из которого они первоначально и росли, когда он представлял собой илистую отмель. Этот глинисто-алевритистый слой содержит многочисленные растительные остатки, в том числе, обугленные. Все стволы почти по всей своей высоте, от оснований до верхушек, захоронены в слое табачно-желтых мелкозернистых пологокосослоистых песков с редкими раковинами двустворчатых моллюсков. Песчаный слой с хвощами перекрыт слоем темно-серых глин и алевритов с многочисленными отпечатками растений и углистыми прослоями. Вероятный сценарий захоронения этих зарослей Equisetites arenaceus в прижизненном положении следующий. Заросли крупных Equisetites arenaceus произрастали на илистой отмели обширного водоема на этапе медленного спокойного накопления глинистых и алевритистых осадков. Этому этапу соответствует нижний глинисто-алевритистый слой 4. Затем режим осадконакопления сменился в сторону относительно кратковременного привноса и спокойной разгрузки в бассейне песчаного материала более полноводными, но не слишком быстрыми потоками, что отражается в пологой косой слоистости песков вышележащего слоя. Уровень воды поднялся, водоем стал достаточно глубоким. Вскоре вся роща древовидных хвощей оказалась затопленной и погребенной в песке в прижизненном вертикальном положении, так как их стволы секут ряд песчаных прослоев слоя 5. Вероятно, последним, наиболее интенсивным привносом песчаного осадочного материала роща Equisetites arenaceus была уничтожена, верхняя часть некоторых из их стволов, вероятно, была обломана (Tverdochlebov, 2004). Этому этапу соответствуют верхи слоя 5. Затем, после заполнения водоема песчаным материалом, скорость осадконакопления уменьшилась, и снова начали отлагаться илистые отмельные отложения (вышележащий глинисто-алевритистый слой 6). Погребенные стволы Equisetites постепенно заполнились и заместились этими тонкозернистыми осадками, образовавшими естественный внутренний отлив растений.

РЕЗУЛЬТАТЫ ПАЛИНОЛОГИЧЕСКОГО ИЗУЧЕНИЯ БУКОБАЙСКИХ ОТЛОЖЕНИЙ ПРЕДШЕСТВУЮЩИХ ЛЕТ
Миоспоры из букобайских отложений были изучены И.С. Макаровой (Шишкин и др., 1995; Tverdokhlebov, 2004). Для выявленного ею палинокомплекса характерно преобладание двухмешковой пыльцы, либо примерно равное соотношение пыльцы и спор. В общем виде он выглядит следующим образом.
Пыльцевая часть представлена немногочисленными видами родов Florinites, Alisporites, Protodiploxypinus (=Microcachryidites, по Макаровой), Minutosaccus, Sulcatisporites, видами Dipterella oblatinoides Maljavkina, Podocarpus (Neopodocarpus) tricoccaeformis f. triassica Maljavkina, Heliosaccus dimorphus Mädler, Chordasporites singulichorda Klaus, Brachysaccus sp., Platysaccus sp., Granosaccus sp., Lunatisporites sp. (=Taeniaesporites, по Макаровой), Vitreisporites pallidus Nilsson (=Caytoniapollenites sp., по Макаровой).
Вид P. (Neopodocarpus) tricoccaeformis f. triassica, судя по описанию и изображениям, приведенным в публикациях В.С. Малявкиной, очень похож на широко распространенный в триасе вид Platysaccus queenslandi de Jersey.
Для споровой части характерно видовое разнообразие спор рода Aratrisporites. Присутствуют виды Punctatisporites punctatus Warjuchina, Carnisporites mesosoicus Mädler, Lycopodiacidites kuepperi Klaus, Duplexisporites gyratus Playford et Dettmann, Polypodites cladophleboides Brick, Acanthotriletes ilekensis Kopytova, Conosmundacidites aff. othmari Klaus, Baculatisporites sp., Verrucosisporites sp., Granulatisporites sp., Neoraistrickia sp., Stereisporites sp. Найдены также споры Leiotriletes rotundus Bolchovitina, Leiotriletes sp., Limbella ovaliformis Maljavkina. Последний из перечисленных видов, судя по довольно схематичному описанию и зарисовкам в публикациях Малявкиной, весьма близок, по нашему мнению, виду Nevesisporites limatulus Playford.
По своему видовому составу изученный И.С. Макаровой палинокомплекс в полной мере отвечает среднетриасовым комплексам миоспор в целом. О возможной его принадлежности к отложениям ладинского возраста может свидетельствовать лишь пыльца вида D. oblatinoides, если на основе данных Макаровой, а также полученных нами, считать указанный вид характерным для палинофлоры ладинского века.
ПАЛИНОКОМПЛЕКС МЕСТОНАХОЖДЕНИЯ ЛЫСОВ
Палинокомплекс из костеносной точки был изучен Н.В. Ильиной. Из образца ИГ КомиНЦ № 437/2002 был выделен чрезвычайно насыщенный и разнообразный комплекс миоспор с доминирующей пыльцевой частью. При анализе его таксономического состава за основу были приняты результаты крупных обобщающих исследований последних лет по триасовой палинофлоре Шпицбергена, акватории Баренцева моря, Южных Альп, севера Средней Сибири и Германского бассейна (Круговых, Могучева, 2000; Ильина, Константинов, 2018; Vigran et al., 1998, 2014; Bachmann, Kozur, 2004; Cirilli, 2010; Hochuli, Vigran, 2010; Kürschner, Herngreen, 2010; Mietto et al., 2012; Hochuli et al., 2015; Ilyina, Konstantinov, 2015 и др.).
По видовому составу палинокомплекс местонахождения Лысов отвечает типичной среднетриасовой, ладинской палинофлоре, хорошо изученной в триасовых осадочных бассейнах (табл. 1). В палинокомплексе весьма многочисленны миоспоры широкого стратиграфического диапазона, унаследованные от раннетриасовой эпохи и с разной степенью детальности распространенные практически повсеместно. В данном случае к ним отнесены пыльца родов Alisporites, Chordasporites, Falcisporites, Platysaccus, Striatoabieites, Sulcatisporites и споры родов Anapiculatisporites, Apiculatisporites, Aratrisporites, Calamospora, Carnisporites, Concavisporites, Cyathidites, Dictyophylidites, Nevesisporites, Polycingulatisporites, Todisporites (Vigran et al., 1998, 2014; Hochuli, Vigran, 2010; Kustatscher et al., 2012; Hochuli et al., 2015).
Таблица 1.
Распределение миоспор палинокомплекса из образца ИГ Коми НЦ № 437/2002 по их ботанической принадлежности и экогруппам
| Ботаническая принадлежность | Таксономический состав | Экогруппы |
|---|---|---|
| Мохообразные | Nevesisporites spp. Polycingulatisporites triangularis Retitriletes sp. |
Влажные низины, речные берега |
| Плауновидные | Anapiculatisporites spp. Aratrisporites spp. |
Прибрежные области |
| Carnisporites spp. | Влажные низины | |
| Anaplanisporites cf. echinatus | ? | |
| Папоротники | Apiculatisporites sp. Concavisporites spp. Converrucosisporites cf. Lygodium japoniciforme Leschikisporites aduncus Cyathidites spp. |
Влажные низины |
| Dictyophyllidites mortoni Gleicheniidites sp. |
Сухие низины | |
| Kyrtomisporis spp. | ? Сухие низины | |
| Duplexisporites spp. | Прибрежные области | |
| Hymenophyllum cf. granulatum | Эпифиты | |
| Lycopodiacidites kuepperi Todisporites spp. |
Влажные низины, речные берега | |
| Punctatosporites walkomi | Влажные низины | |
| Camptotriletes cerebriformis | ? | |
| Членистостебельные | Calamospora spp. | Речные берега |
| Птеридоспермы | Falcisporites spp. | Территории с холмистым рельефом и территории, удаленные от побережья |
| Sulcatisporites spp. Vitreisporites pallidus |
Сухие низины | |
| Хвойные | Araucariacites sp. Florinites spp. Podosporites spp. |
Прибрежные области |
| Protodiploxypinus spp. | Прибрежные области/пионерная флора | |
| Cyclosaccus cf. podocarpoides Dacrycarpites spp. Enzonalasporites spp. Lunatisporites rhaeticus Ovalipollis spp. Piceaepollenites sp. Podocarpidites spp. cf. Triadispora spp. Voltziaceaesporites heteromorpha Lueckisporites triassicus |
Территории с холмистым рельефом и территории, удаленные от побережья | |
| Kuglerina meieri | ?Сухие низины | |
| Patinasporites densus | Сухие низины | |
| Vallasporites ignacii | Сухие низины, ?территории с холмистым рельефом и территории, удаленные от побережья | |
| Dipterella oblatinoides Minutosaccus spp. Plicatisaccus badius Praecirculina granifer Pseudenzonalasporites summus Samaropollenites speciosus |
?Территории с холмистым рельефом и территории, удаленные от побережья | |
| ?Хвойные | Infernopollenites spp. Institisporites cf. crispus Succinctisporites grandior |
?Территории с холмистым рельефом и территории, удаленные от побережья |
| Хвойные + птеридоспермы | Alisporites spp. | Влажные низины, территории с холмистым рельефом и территории, удаленные от побережья |
| Chordasporites spp. Pityosporites cf. scaurus Striatoabieites spp. |
Территории с холмистым рельефом и территории, удаленные от побережья | |
| Platysaccus spp. | Прибрежные области | |
| Цикадовые | Brachysaccus spp. | ?Сухие низины |
| Chasmatosporites spp. Eucommiidites spp. |
Сухие низины | |
| Цикадовые, гинкговые, беннетитовые, птеридоспермы | Cycadopites sp. | Сухие низины |
| Вероятные гимноспермовые | Accinctisporites spp. Colpectopollis ellipsoideus |
? |
Палинокомплекс местонахождения Лысов содержит также группу таксонов, которые составляют основу всех среднетриасовых комплексов миоспор. При этом значительная их часть, представленная преимущественно единичными экземплярами, появляется в самых верхах верхнеоленекского подъяруса и его возрастных аналогов, а систематическое распространение связано уже со среднетриасовыми отложениями. Из них в палинокомплексе представлены споры рода Duplexisporites и виды L. kuepperi, Camptotriletes cerebriformis Naumova, Converrucosisporites cf. Lygodium japoniciforme Ivanova. Пыльца представлена видами родов Florinites (Illinites chitonoides Klaus в англоязычной литературе), Minutosaccus, Protodiploxypinus, Triadispora и видами Accinctisporites circumdatus Jain, Podocarpidites keuperianus Schuurman, Podosporites amicus Scheuring, Succinctisporites grandior Leschik, Voltziaceaesporites heteromorpha Klaus (Ильина, 2008; Hochuli et al., 1989; Hochuli, Vigran, 2010; Kürschner, Herngreen, 2010; Vigran et al., 1998, 2014).
Другую часть этой группы составляют миоспоры, последовательное появление которых связано с анизийским веком. Из форм, первое появление которых связано с нижнеанизийским уровнем, в палинокомплексе местонахождения Лысов встречена пыльца Accinctisporites ligatus (Leschik) Clarke, Araucariacites australis Cookson, Colpectopollis ellipsoideus Visscher, Infernopollenites cf. сlaustratus Dolby, Institisporites cf. сrispus Pautsch, Lueckisporites triassicus Clarke, Ovalipollis spp., Samaropollenites speciosus Dolby et Balme и споры Retitriletes sp., а со среднеанизийским – пыльца Brachysaccus neomundanus (Leschik) Mädler, Chasmatosporites spp., Dacrycarpites europeaus Mädler, Enzonalasporites cf. vigens Leschik, Eucommiidites cf. microgranulatus Scheuring, Kuglerina meieri Scheuring, Praecirculina granifer (Leschik) Klaus emend. Scheuring, Pseudenzonalasporites summus Scheuring, Vallasporites ignacii (Leschik) Scheuring и споры Leschikisporites aduncus (Leschik) Potonie.
Все перечисленные виды переходят в комплексы миоспор ладинского и позднетриасового возраста. Следует подчеркнуть, что первоначально первое появление многих из них было зафиксировано на более высоких стратиграфических уровнях, а приуроченность к анизийскому ярусу и его аналогам определена в результате палинологических исследований последних лет, причем в большинстве случаев уточненное стратиграфическое положение подтверждено сопутствующими комплексами морских беспозвоночных (Круговых, Могучева, 2000; Visscher, Krystyn, 1978; Visscher, Brugman, 1981; Hochuli et al., 1989, 2015; Vigran et al., 1998, 2014; Ilyina, Egorov, 2008; Hochuli, Vigran, 2010; Kürschner, Herngreen, 2010; Ilyina, Konstantinov, 2015).
В следующую группу палинокомплекса местонахождения Лысов входят миоспоры, появление которых связано с ладинским веком. Из них определены пыльца Cyclosaccus cf. podocarpoides Mädler, Patinasporites densus (Leschik) Scheuring, Plicatisaccus badius Pautsch и споры Hymenophyllum cf. granulatum Faddeeva, Kyrtomisporis cf. ervii Van der Eem, Kyrtomisporis specious Mädler, Punctatosporites walkomi de Jersey. В этой группе также есть виды, первоначально ставшие известными из более молодых отложений, а впоследствии встреченные с аммоноидеями в отложениях ладинского возраста (Ильина, Константинов, 2018; Vigran et al., 1998, 2014; Ilyina, Egorov, 2008; Hochuli et al., 2015).
Последней и самой интересной в составе палинокомплекса местонахождения Лысов является группа миоспор, которые до недавнего времени были известны только из верхнетриасовых отложений. Это пыльца видов D. oblatinoides, Lunatisporites rhaeticus (Schulz) Warrington, сf. Pityosporites scaurus (Nilsson) Schulz, и споры Kyrtomisporis cf. gracilis Bjaerke et Manum и Gleicheniidites sp.
Стратиграфическое положение некоторых указанных выше таксонов спор и пыльцы требует пояснения. Пыльца V. ignacii, P. densus, P. summus и S. speciosus включена в группу видов, маркирующих начало карнийского века. Первоначально первое появление этих видов было установлено в аммоноидной зоне Daxatina canadensis, подошва которой в разрезе Prati di Stuores/Stuores Wiesen Южных Альп принята за стратотип границы между средним и верхним триасом (Cirilli, 2010; Mietto et al., 2012). Однако в дальнейшем они были обнаружены и в среднетриасовых отложениях, датированных аммоноидеями, а также в изученном нами образце из местонахождения букобайской фауны тетрапод. Этот факт существенно расширяет стратиграфический диапазон указанных видов и позволяет считать их характерными для среднетриасовых отложений в целом и палинокомплексов ладинского возраста в частности и, таким образом, выводит из группы видов-индексов для определения начала карнийского века.
Отдельно следует отметить споры рода Gleicheniidites, типичного элемента позднемезозойских отложений, который до недавнего времени (Vigran et al., 2014) в триасовых отложениях известен не был. В карнийских отложениях акватории Баренцева моря был определен вид Gleicheniidites senonicus Ross. В Южных Альпах вид Gleicheniidites sp. определен в составе палинокомплекса из верхней части лонгобардского подъяруса (Hochuli et al., 2015). Вероятно, следует признать, что в палинологической летописи споры Gleicheniidites появились гораздо раньше, чем это было принято считать.
Принадлежность миоспор D. oblatinoides, L. rhaeticus, сf. P. scaurus и K. cf. gracilis к отложениям ладинского возраста необходимо проверить на дополнительном фактическом материале.
Наряду с перечисленными выше группами, палинокомплекс местонахождения Лысов содержит единичные миоспоры, характерные для отложений индского и раннеоленекского возраста, а на данном стратиграфическом уровне, вероятнее всего, являющиеся переотложенными. К ним относятся споры Lundbladispora willmottii Balme и пыльца Cordaitina minor Pautsch, Crustaesporites globosus Jansonius, Klausipollenites cf. schaubergeri (Potonie et Klaus) Jansonius, Lunatisporites acutus (Leschik) Scheuring, L. noviaulensis (Leschik) Scheuring, Protohaploxypinus microcorpus (Schaarschmidt), Clarke sensu Morbey, P. samoilovichii (Jansonius) Hart, Striatites jacobii Jansonius.
В палинокомплексе местонахождения Лысов выделяется также большая группа юрско-меловых миоспор: споры Cingulatitriletes sp. и пыльца Cedripites sp., Piceae valanjinica Rovnina, P. parvireticulata Rovnina, P. pseudorotundiformis Maljavkina, Picea sp., Pseudopicea variabiliformis (Maljavkina) Bolchovitina, Pinuspollenites sp. Присутствие перечисленных форм в букобайских отложениях может быть результатом их привноса из вышележащих толщ.
ДОПОЛНИТЕЛЬНОЕ ОБОСНОВАНИЕ ЛАДИНСКОГО ВОЗРАСТА ПАЛИНОКОМПЛЕКСА МЕСТОНАХОЖДЕНИЯ ЛЫСОВ
На территории Европейской России отложения среднетриасового возраста палинологически наиболее полно изучены в Прикаспийской впадине и в Тимано-Североуральском регионе. В Прикаспийской впадине в основу биостратиграфического расчленения среднетриасовых отложений по миоспорам положены результаты их изучения в опорной скважине 1-Хобда. Сходство видового состава палинокомплекса местонахождения Лысов наблюдается с палинокомплексом Heliosaccus dimorphus, который характеризует верхнекиильскую посвиту, мастексайскую и акмамыкскую свиты (Ярошенко и др., 2001). Первой общей чертой комплексов является доминирующая роль пыльцы в целом и двухмешковых форм в частности. Из форм широкого стратиграфического диапазона общими являются споры Anapiculatisporites spiniger (Leschik) Reinhardt и A. telephorus Klaus, L. kuepperi, Carnisporites spp., Todisporites spp., а также многочисленные в прикаспийском комплексе Aratrisporites spp., Limatulasporites spp. (синоним Nevesisporites), Polycingulatisporites spp. В пыльцевой части – это представители родов Alisporites, Chordasporites, Falcisporites, Platysaccus, Striatoabieites, Sulcatisporites, Vitreisporites, Cycadopites и вид V. heteromorpha.
Сходство также наблюдается по группе миоспор, составляющих основу среднетриасовых палинокомплексов: это споры Duplexisporites spp., Converrucosisporites и пыльца Accinctisporites spp., Illinites spp. (синоним Florinites spp.), Minutosaccus spp., Protodiploxypinus spp. (синоним Microcachryidites spp.), Triadispora spp., B. neomundanus, C. ellipsoideus, P. keuperianus, P. amicus. Из таксонов, последовательное появление которых связано с анизийским веком, общими являются споры L. aduncus, P. walkomi и пыльца C. apertus, Enzonalasporites sp., E. microgranulatus, Ovalipollis spp., P. granifer. Пыльца рода Infernopollenites представлена в прикаспийском комплексе миоспор видом I. sulcatus (Pautsch) Scheuring.
Виды, характерные для отложений ладинского возраста, в сравниваемых комплексах представлены спорами K. ervii.
Еще одной общей чертой обсуждаемых комплексов миоспор является заметное присутствие пыльцы хвойных растений группы Circumpolles и некоторых одномешковых форм, многочисленной в отложениях ладинского возраста и в верхнем триасе Тетической области и весьма редкой в Бореальных разрезах. В прикаспийском комплексе эта группа представлена родами Camerosporites, Duplicisporites и видом P. granifer, а одномешковые – определенным в открытой номенклатуре видом Enzonalasporites sp. В палинокомплексе хут. Лысов – это пыльцевые зерна E. cf. vigens, P. granifer, P. densus, P. summus и V. ignacii.
По видовому составу миоспор палинокомплекс местонахождения Лысов близок палинокомплексу Converrucosisporites conferteornatus–Florinites pseudostriatus Тимано-Североуральского региона, ладинский возраст которого определен по совместному нахождению с комплексом тетрапод фауны Mastodonsaurus в надкраснокаменской свите и на основе широкой межрегиональной корреляции с отложениями ладинского возраста морского происхождения (Ильина, 2001). В палинокомплексе C. conferteornatus–F. pseudostriatus также преобладает пыльца, но споровая часть при этом более насыщенная.
В группе миоспор широкого стратиграфического диапазона общие для двух палинокомплексов споры родов Anapiculatisporites, Apiculatisporites, Aratrisporites, Nevesisporites, Carnisporites, Todisporites, Dictyophyllidites, Cyathidites и Concavisporites многочисленнее и в видовом отношении разнообразнее. Присутствуют виды L. kuepperi, C. cerebriformis. Их дополняют споры родов Verrucosisporites и Cyclotriletes и видов Cyclogranisporites arenosus Mädler, Cycloverrutriletes presselensis Schulz, Camarozonosporites rudis (Leschik) Klaus, не встреченные в палинокомплексе местонахождения Лысов. Пыльца родов Alisporites, Chordasporites, Falcisporites, Platysaccus, Striatoabieites, Sulcatisporites в палинокомплексе C. conferteornatus–F. pseudostriatus также многочисленна, как и в палинокомплексе местонахождения Лысов.
Пыльцевая часть группы таксонов, составляющих основу среднетриасовых комплексов миоспор, у рассматриваемых палинокомплексов по качественному и количественному составу практически одинакова. Споры разнообразнее в палинокомплексе C. conferteornatus–F. pseudostriatus. Кроме общих Duplexisporites spp., здесь определены представители родов Converrucosisporites, Concentricisporites, Baculatisporites, Foveosporites, Uvaesporites и вид Conbaculatisporites mesosoicus Klaus.
Из миоспор, характерных для разных уровней среднетриасовых отложений, общими являются споры L. aduncus, P. walkomi и Kyrtomisporis sp. В палинокомплексе C. conferteornatus–F. pseudostriatus эта группа более представительна за счет спор Uvaesporites spp., Acanthotriletes spp., Granulatisporites spp., Microreticulatisporites opacus (Leschik) Klaus, Neoraistrickia taylori Playford et Dettmann, Porcellispora longdonensis (Clarke) Scheuring emend. Morbey, P. cladophleboides, Sellaspora rugoverrucata van der Eem и некоторых других. В пыльцевой части общих таксонов больше: это многочисленные Ovalipollis spp., Chasmatosporites spp. и виды S. speciosus, L. triassicus, P. badius, P. granifer, P. summus, P. densus, Eucommiidites sp.
Для дополнительного обоснования возраста палинокомплекса хут. Лысов следует обратиться к материалам по палинологии отложений ладинского яруса Шпицбергена, акватории Баренцева моря и Южных Альп, где они наиболее полно изучены и надежно датированы фауной аммоноидей. В Баренцевоморском бассейне отложения ладинского возраста охарактеризованы комплексом миоспор палинозоны Echinitosporites iliacoides (Hochuli et al., 1989; Vigran et al., 1998, 2014). В доминирующей пыльцевой части здесь определены некоторые виды родов Lunatisporites и Alisporites. В большом количестве присутствуют разнообразные Angustisulcites spp., Illinites spp., Podosporites spp., Protodiploxypinus spp., Striatoabieites spp., Triadispora spp. Широкое распространение имеют виды Infernopollenites parvus Scheuring и I. sulcatus, Staurosaccites quadrifidus Dolby, Ovalipollis pseudoalatus (Thiergart) Schuurman, Chasmatosporites sp. A sensu Vigran et al., 2014, A. circumdatus, A. australis, Cordaitina gunyalensis (Pant et Srivastava) Balme, K. meieri, Dyupetalum vicentinense Brugman, E. microgranulatus и некоторые другие. Многочисленна пыльца E. iliacoides Schulz et Krutzsch. Подчеркнем присутствие редких экземпляров Duplicisporites granulatus (Leschik) Scheuring, D. scurrilis Scheuring и P. densus.
В споровой части многочисленны Aratrisporites spp., Striatella seebergensis Mädler (синоним D. gyratus), L. aduncus и гладкие треугольные споры родов Deltoidospora и Dictyophyllidites. Определены Acanthotriletes spp., Annulispora spp., Gordonispora spp., Perotrilites spp., Stereisporites spp., Uvaesporites spp., виды A. spiniger, Baculatisporites comaumensis (Cookson) Potonié, Conbaculatisporites hopensis Bjærke et Manum, C. rudis, Lycopodiacidites kokenii van der Eem, N. taylori. Следует обратить внимание на развитие в баренцевоморском палинокомплексе спор Kraeuselisporites cooksonae (Klaus) Dettmann, K. dentatus Leschik, Zebrasporites interscriptus (Thiergart) Klaus, Z. kahleri Klaus, Semiretisporis sp. A sensu Vigran et al., 2014, Ischyosporites sp., Gleicheniidites sp. и особенно Densoisporites velatus Weyland et Krieger. Последний из перечисленных видов является характерным для юрско-меловых отложений. Появление его в отложениях ладинского яруса либо меняет его стратиграфический диапазон, либо является следствием засорения пробы. Наряду с перечисленными формами, в палинокомплексе определено присутствие раннетриасовых спор Verrucosisporites spp., Cyclotriletes spp., C. presselensis.
В Южных Альпах ладинский ярус охарактеризован четырьмя палинокомплексами, выделенными по последовательному обновлению видового состава палинофлоры за счет первого появления в разрезе характерных групп миоспор (Hochuli et al., 2015). У палинокомплекса местонахождения Лысов с альпийской палинофлорой много общих черт.
В альпийских палинокомплексах также преобладает пыльца, и она гораздо разнообразнее в видовом отношении. Из форм широкого стратиграфического диапазона определены представители родов Alisporites, Chordasporites, Lunatisporites, Platysaccus, Striatoabieites, Sulcatisporites, вид V. heteromorpha. Остальная часть – это пыльца родов Angustisulcites, Araucariacites, Brachysaccus, Camerosporites, Duplicisporites, Enzonalasporites, Eucommidites, Illinites, Infernopollenites, Ovalipollis, Triadispora, виды Aulisporites astigmosus (Leschik) Klaus, C. gunyalensis, D. vicentinense, E. iliacoides, H. dimorphus, K. meieri, P. keupeianus, P. amicus, S. quadrifidus, а также некоторые другие формы, которые не были встречены в рассмотренных выше осадочных бассейнах.
В споровой части из форм широкого стратиграфического диапазона присутствуют Aratrisporites spp., Densoisporites spp., Punctatisporites spp., Verrucosisporites spp., Cyclogranisporites spp., Todisporites spp., C. mesozoicus, A. spiniger. Остальные споры – это Annulispora spp., Baculatisporites spp., Converrucosisporites spp., Convolutispora spp., Foveosporites spp., Reticulatisporites spp., Uvaesporites spp., K. dentatus, K. ervii, L. kokenii, N. taylori, P. longdonensis, Sellaspora foveorugulata van der Eem, S. rugoverrucata, Gleicheniidites sp. и некоторые другие виды. Следует еще раз подчеркнуть, что присутствие спор Gleicheniidites sp. в палинофлоре ладинского яруса, скорее всего, не случайно.
По нашему мнению, сходство палинокомплексов из отложений ладинского возраста Баренцевоморского бассейна, Тимано-Североуральского региона, Прикаспийской впадины и Южных Альп несомненно. Особенно важно то, что в Баренцевоморском бассейне и Южных Альпах они датируются этим временем непосредственно, по сопутствующей морской фауне. Общность видового состава палинофлоры перечисленных осадочных бассейнов и палинокомплекса местонахождения Лысов убедительно свидетельствует о ладинском возрасте последнего.
ЭКОЛОГИЧЕСКАЯ ИНТЕРПРЕТАЦИЯ РЕЗУЛЬТАТОВ ПАЛИНОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Комплекс листовой флоры из местонахождения Лысов представлен фрагментами членистостебельных, папоротников, птеридоспермов, гинкгофитов и цикадофитов (см. выше). Вместе с тем, видовой состав изученного палинокомплекса позволяет предположить существование в ладинском веке более разнообразной растительности. По результатам палинологических исследований была проведена реконструкция растительного покрова. Затем видовой состав палинокомплекса Лысова был проанализирован с помощью метода экогрупп спороморф (SEG-analysis) для установления условий произрастания растений-продуцентов и метода морфогрупп, определяющего степень их гигрофильности и ксерофильности (Visscher, van der Zwan, 1981; Balme, 1995; Abbink, 1998; Abbink et al., 2004; Kustatscher et al., 2010, 2012; Lindström et al., 2017; Paterson et al., 2017 и др.). Список растительности дополнили мохообразные, плауновидные и хвойные, ставшие доминирующей группой с высоким содержанием ксерофильных элементов (табл. 1, рис. 5). Количественное распределение миоспор по экогруппам показано на рис. 6.
Рис. 5.
Количественное распределение миоспор палинокомплекса из образца ИГ Коми НЦ № 437/2002 по их ботанической принадлежности: 1 – мохообразные, 2 – плауновидные, 3 – папоротники, 4 – членистостебельные, 5 – птеридоспермы, 6 – хвойные + птеридоспермы, 7 – хвойные + ? хвойные, 8 – цикадовые + цикадовые, гинкговые, беннетитовые, птеридоспермы, 9 – вероятные гимноспермовые.
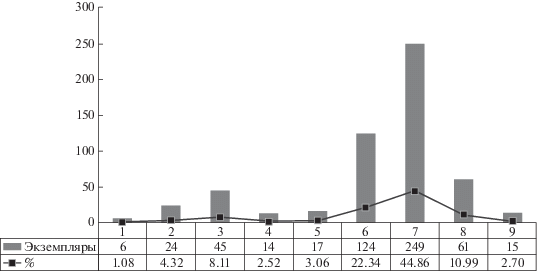
Рис. 6.
Количественное распределение миоспор палинокомплекса из образца ИГ Коми НЦ № 437/2002 по экогруппам, составленное с помощью метода экогрупп спороморф: 1 – влажные низины, 2 – прибрежные области, 3 – речные берега, 4 – сухие низины, 5 – территории с холмистым рельефом и территории, удаленные от побережья, 6 – миоспоры с неустановленной принадлежностью какой-либо экогруппе.
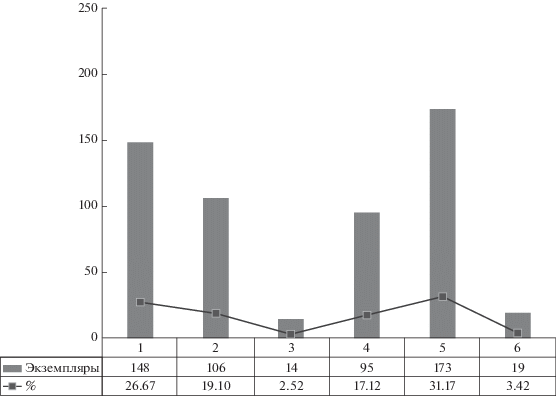
Среди споровых растений преобладающей группой оказались папоротники, которые произрастали в условиях теплого (субтропического и тропического) и влажного климата, занимали берега рек, влажные низины, пресноводные марши, были частью болотных ассоциаций, а также формировали подлесок лесных массивов (Kustatscher et al., 2012). Плауновидные колонизировали прибрежные области, влажные низины, заболоченные участки. Членистостебельные произрастали по берегам стоячих водоемов и рек, в заболоченных поймах. Пыльца птеридоспермов, найденная в составе палинокомплекса хут. Лысов, принадлежала растениям сухих низин. В целом же птеридоспермы были растениями, произраставшими в разных климатических условиях, от засушливых до жарких и влажных. В объемной группе пыльцы хвойных основную часть зерен продуцировали растения, обитавшие в пределах территорий с холмистым рельефом вдали от прибрежных областей. Только небольшую часть пыльцы продуцировали растения сухих низин и пойм. Существенная часть двухмешковой пыльцы в палинокомплексе хут. Лысов могла быть продуктом как хвойных, так и птеридоспермов. Местом обитания цикадовых были равнинные территории. Вместе с папоротниками и членистостебельными они являются индикатором гумидных обстановок.
Миоспоры экогрупп болот, влажных и сухих низин, прибрежных областей и берегов рек в совокупности представляют растительный покров равнинной территории с достаточно широким развитием увлажненных биотопов. Следовательно, необходимо найти объяснение присутствию большого количества двухмешковой пыльцы растений из более засушливых областей. Поскольку растения-источники двухмешковой пыльцы имеют высокую репродуктивную способность, а основным способом транспортировки их пыльцы является перенос ветром, то ее высокое содержание в комплексах миоспор может быть результатом пыльцевых дождей (Kustatscher et al., 2012). Поэтому вполне вероятно, что в палинокомплексе местонахождения Лысов двухмешковая пыльца большей частью является привнесенной.
ЗАКЛЮЧЕНИЕ
Изучение позвоночных, двустворчатых моллюсков и растительных остатков из местонахождения Лысов дает новую важную информацию для характеристики верхней части букобайской свиты среднего триаса в Южном Приуралье. Комплекс позвоночных в этой части разреза не показывает изменений по сравнению с более полно фаунистически охарактеризованной нижней частью букобайской свиты. Вместе с тем, неожиданной оказалась находка остатков темноспондильных амфибий из семейства Rhytidosteidae, так как их распространение во всем мире до сих пор было ограничено ранним триасом. Макрофлора из данного местонахождения, наиболее вероятно, имеет ладинский возраст. Из местонахождения Лысов изучен насыщенный и разнообразный комплекс миоспор. Его ладинский возраст определяется на основе широкой межрегиональной корреляции его видового состава с другими палинокомплексами, в том числе, надежно датированными фауной аммоноидей. Результаты реконструкции растительного покрова по данным палинологических исследований показали, что его формирование происходило в условиях равнины с достаточно широким развитием увлажненных биотопов, что согласуется с данными по условиям захоронения остатков тетрапод.
Работа выполнена при финансовой поддержке РФФИ в рамках научных проектов №№ 17-04-00410, 17-54-10013, а также в рамках государственной программы повышения конкурентоспособности Казанского (Приволжского) федерального университета среди ведущих мировых научно-образовательных центров.
Список литературы
Добрускина И.А. Стратиграфическое положение флороносных толщ триаса Евразии. М.: Наука, 1980. 163 с.
Добрускина И.А. Триасовые флоры Евразии. М.: Наука, 1982. 196 с.
Ильина Н.В. Палиностратиграфия среднего триаса Тимано-Североуральского региона. Екатеринбург: УрО РАН, 2001. 230 с.
Ильина Н.В. Граница нижнего и среднего триаса в Тимано-Североуральском регионе // Стратигр. седиментол. нефтегаз. басс. 2008. № 1. С. 15–30.
Ильина Н.В., Константинов А.Г. Палиноассоциации ладинских (средний триас) отложений в разрезе мыса Цветкова, Восточный Таймыр, север Средней Сибири // Эволюция жизни на Земле: Матер. V междунар. симпоз. / Ред. Подобина В.М. Томск: Издательский дом Томского гос. ун-та, 2018. С. 153–155.
Круговых В.В., Могучева Н.К. Палино- и фитостратиграфия опорного разреза триаса мыса Цветкова на Восточном Таймыре // Геол. геофиз. 2000. Т. 41. № 4. С. 535–550.
Твердохлебов В.П. Стратиграфия среднего и верхнего триаса Южного Приуралья // Вопр. геол. Южного Урала и Поволжья. 1967. Вып. 4. Ч. 1. С. 78–91.
Шишкин М.А., Очев В.Г., Твердохлебов В.П. и др. Биостратиграфия континентального триаса Южного Приуралья. М.: Наука, 1995. 206 с.
Ярошенко О.П., Макарова И.С., Романовская Г.М. и др. Палинокомплексы отложений среднего и верхнего триаса Прикаспийской впадины // Недра Поволжья и Прикаспия. 2001. Вып. 26. С. 10–18.
Abbink O.A. Palynological identification in the Jurassic of the North Sea region // LPP contrib. ser. 1998. V. 8. P. 1–192.
Abbink O.A., Van Koijnenburg-Van Cittert J.H.A., Visscher H. A sporomorph ecogroup model for the Northwest European Jurassic–Lower Cretaceous: Concepts and framework // Neth. J. Geosci. 2004. V. 83. P. 17–38.
Bachmann G.H., Kozur H. The Germanic Triassic: correlations with the international chronostratigraphic scale, numerical ages and Milankovitch-cyclicity // Hall. J. Geowiss. 2004. Bd 26. P. 17–62.
Balme B.E. Fossil in situ spores and pollen grains: an annotated catalogue // Rev. Palaeobot. Palynol. 1995. V. 87. P. 81–323.
Cirilli S. Upper Triassic–lowermost Jurassic palynology and palynostratigraphy: a review // The Triassic Timescale / Ed. Lucas S.G. Geological Society, London, Spec. Publ. V. 334. L.: Geol. Soc., 2010. P. 285–314.
Hochuli P.A., Colin J.P., Vigran J.O. Triassic biostratigraphy of the Barents Sea area // Correlation in Hydrocarbon Exploration / Ed. Collinson J.D. L.: Graham & Trotman, 1989. P. 131–153.
Hochuli P.A., Roghi G., Brack P. Palynological zonation and particulate organic matter of the Middle Triassic of the Southern Alps (Seseda and Val Gola-Margon sections, Northern Italy) // Rev. Palaeobot. Palynol. 2015. V. 218. P. 28–47.
Hochuli P.A., Vigran J.O. Climate variations in the Boreal Triassic – inferred from palynological records from the Barents Sea // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2010. V. 290. P. 20–42.
Ilyina N.V., Egorov A.Yu. The Upper Triassic of northern Middle Siberia: stratigraphy and palynology // Polar Res. 2008. V. 27. P. 372–392.
Ilyina N., Konstantinov A. New biostratigraphic data on Anisian (Middle Triassic) miospores from Cape Tsvetkov section, Northern Middle Siberia, Russia // Ber. Inst. Erdwiss. Karl-Franzens-Univer. Graz. 2015. Bd 21. S. 174.
Kürschner W.M., Herngreen G.F.W. Triassic palynology of central and northwestern Europe: a review of palynofloral diversity patterns and biostratigraphic subdivisions // The Triassic Timescale / Ed. S.G. Lucas. Geological Society, London, Spec. Publ. V. 334. L.: Geol. Soc., 2010. P. 263–283.
Kustatscher E., Heunisch C., Van Konijnenburg-Van Cittert J.H.A. Taphonomical implications of the Ladinian megaflora and palynoflora of the Thale (Germany) // PALAIOS. 2012. V. 27. P. 753–764.
Kustatscher E., Van Konijnenburg-Van Cittert J.H.A., Roghi G. Macrofloras and palynomorphs as possible proxies for palaeoclimatic and palaeoecological studies: A case study from the Pelsonian (Middle Triassic) of Kühwiesenkopf/Monte Prà della Vacca (Olang Dolomites, N-Italy) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2010. V. 290. P. 71–80.
Lindström S., Erlström M., Piasecki S. et al. Palynology and terrestrial ecosystem change of the Middle Triassic to lowermost Jurassic succession of the eastern Danish Basin // Rev. Palaeobot. Palynol. 2017. V. 244. P. 65–95.
Mietto P., Manfrin S., Preto N. et al. The global boundary stratotype section and point (GSSP) of the Carnian Stage (Late Triassic) at Prati di Stuores/Stuores Wiesen Section (Southern Alps, NE Italy) // Episodes. 2012. V. 35. P. 414–430.
Paterson N.W., Mangerud G., Mørk A. Late Triassic (early Carnian) palynology of shallow stratigraphical core 7830/5-U-1, offshore Kong Karl Land, Norwegian Arctic // Palynol. 2017. V. 41. P. 230–254.
Tverdokhlebov V.P. Buried Equisetites thickets from the Middle Triassic of the south Cis-Urals, Russia // N. J. Geol. Paläontol. Monats. 2004. Bd 10. S. 595–604.
Vigran J.O., Mangerud G., Mørk A. et al. Biostratigraphy and sequence stratigraphy of the Lower and Miidle Triassic deposits from the Svalis Dome, Central Barents sea, Norway // Palynology. 1998. V. 22. P. 89–141.
Vigran J.O., Mangerud G., Mørk A. et al. Palynology and geology of the Triassic succession of Svalbard and the Barents Sea // Geol. Surv. Norway Spec. Publ. 2014. V. 14. 270 p.
Visscher H., Brugman W.A. Ranges of selected palynomorphs in the Alpine Triassic of Europe // Rev. Palaeobot. Palynol. 1981. V. 34. P. 115–128.
Visscher H., Krystyn L. Aspects of Late Triassic palynology. 4. A palynological assemblage from ammonoid-controlled late Karnian (Tuvalian) sediments of Sicily // Rev. Palaeobot. Palynol. 1978. V. 26. P. 115–128.
Visscher H., van der Zwan C.J. Palynology of the circum-Mediterranean Triassic: Phytogeographical and palaeoclimatological implications // Geol. Rundschau. 1981. Bd 70. P. 625–634.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал