

Палеонтологический журнал, 2020, № 4, стр. 89-92
Новая ящерица (Agamidae, Iguania) из позднего палеоцена Южной Монголии
В. Р. Алифанов *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: valifan@paleo.ru
Поступила в редакцию 27.02.2018
После доработки 23.09.2019
Принята к публикации 27.12.2019
Аннотация
Приведено описание ящерицы Epileolis reshetovi gen. et sp. nov. из позднего палеоцена Монголии. Новая форма установлена по левой зубной кости в составе семейства Agamidae. Она отличается большим числом клыкообразных зубов, зауженным основанием зубов пренатальной серии и высоким короноидным отростком зубной кости.
ВВЕДЕНИЕ
Представители семейства Agamidae (Iguania, Lacertilia), распространенного ныне в Старом Свете и в Австралии, отличаются разделением зубов на три серии: “клыки”, “предкоренные” и “коренные”. С увеличением возраста большая часть или все зубы локализуются вблизи свободного края высокого зубного гребня челюстной кости. Постклыковые зубы всегда срастаются с несущей костью и теряют способность к сменности. Зубную систему агамид часто рассматривают примером акродонтного типа прикрепления, не имеющего ясного определения.
Представления о надродовой классификации современных агамовых сформулировал С. Муди (Moody, 1980). По ним, в частности, роды Uromastyx и Leiolepis классифицированы в семействе Uromastycidae. Их связь поддерживается фено-кладистически (напр., Gauthier et al., 2012), но исключается данными молекулярной систематики (напр., Pyron et al., 2013). Заметим, что у Leiolepis, как у типичных Agamidae, гетеродонтия полная (разделение на все три отмеченных выше серии зубов представлено не только на нижней, но и на верхней челюсти), чего нет у Uromastyx, у которых верхнечелюстные “клыки” всегда отсутствуют.
Нами семейства Uromastycidae и Agamidae классифицированы в микроотряде Pachyglossa (верхнечелюстные кости входят в обрамление суборбитальных окон, зубы лабио-лингвально уплощены) инфраотряда Agamognatha, в котором также рассмотрены ископаемые Polyglyphanodontia sensu lato (Adamisauridae, Polyglyphanodontidae: Алифанов, 2000, 2012). Пахиглоссы, помимо современных семейств, включают в себя позднемеловых Isodontosauridae и палеогеновых Changjiangosauridae. У первых зубы сохраняют сменность (Алифанов, 2004), а у вторых они прирастают к несущим костям без возможности смены, но при опоре зубов на субдентальный гребень, что сопровождается низким зубным гребнем (Алифанов, 2009).
Данные по Isodontosauridae интересны с точки зрения обсуждения проблемы места происхождения пахиглосс. Им, вероятнее всего, являлась Северная Америка, поскольку именно отсюда архаичные пахиглоссы в виде позднемелового семейства Isodontosauridaе могли проникнуть и в Южную Америку (Gueragama Simões et al., 2015), и в Центральную Азию (Isodontosaurus Gilmore, 1943, Parauromastyx Alifanov, 2004). Поочередная связь Северной Америки сначала с Южной Америкой, а затем с Центральной Азией в конце раннего мела и на рубеже раннего и позднего мела соответственно предполагалась ранее (Каландадзе, Раутиан, 1992; Kalandadze, Rautian, 1997).
Начальная радиация агамид проходила в палеогене Центральной Азии на фоне процветания Changjiangosauridae (Changjiangosaurus Hou, 1976 и Qianshanosaurus Hou, 1974, поздний палеоцен Китая; Creberidentat Li, 1991, средний эоцен Китая; Khaichinsaurus Alifanov, 2009, Lentisaurus Alifanov, 2009, Acrodontopsis Alifanov, 2009, Agamimus Alifanov, 2009, Graminisaurus Alifanov, 2009 и Lavatisaurus Alifanov, 2009, средний эоцен Монголии) и Uromastycidae (Pseudotinosaurus Alifanov, 1991, средний эоцен Монголии и Китая; Brevidensilacerta Li, 1991, средний эоцен Китая; Mergenagama Gao et Dashzeveg, 1999, Talosaurus Gao et Dashzeveg, 1999 и Zephyrosaurodion Gao et Dashzeveg, 2000, средний эоцен Монголии). Разнообразие ранних Agamidae тесно связано с родом Tinosaurus, который четкого диагноза не имеет (Estes, 1983; Алифанов, 2000, 2012; Smith et al., 2011). К последнему отнесен ряд видов из Китая (T. doumuensis Hou, 1974, поздний палеоцен; T. yuanquensis Li, 1991 и T. lushihensis Dong, 1965, поздний эоцен), Казахстана (T. postremus Averianov, 2000, поздний палеоцен–ранний эоцен) и Индии (T. indicus Prasad et Bajpai, 2008, ранний эоцен). Некоторые материалы из Монголии представлены в открытой номенклатуре (Tinosaurus sp.; средний палеоцен: Алифанов, 2000, 2012; ранний олигоцен: Böhme, 2007). Находки без ясного таксономического определения указаны (Smith et al., 2011) также из среднего эоцена Китая (Дзилинь).
В палеогене существовали агамоподобные формы с коническими и трехвершинными постклыковыми зубами. Принадлежность их к Agamidae доказывается наличием клыковидных верхнечелюстных зубов, которые точно известны у ящериц из верхнего палеоцена провинции Аньхой (‘Tinosaurus’ doumuensis Hou, 1974, Anhuisaurus huainanensis Hou, 1974: Dong et al., 2016) и среднего эоцена провинции Гирин (Smith et al., 2011) в Китае. Они также установлены у Suratagama neeraae Rana et al., 2013 и ‘Tinosaurus’ indicus Prasad et Bajpai, 2008 из нижнего эоцена Индии (Rana et al., 2013). Для большинства видов выявлено наличие U-образного контура нижнего края зубов постнатальной серии (неагамидным пахиглоссам свойственен их V- или Y-образный контур). Некоторые из них несут диастему между зубами пре- и постнатальной серий. Эта же особенность выражена у Anhuisaurus huainanensis Hou, 1974 из верхнего палеоцена Китая (Dong et al., 2016) и у Tinosaurus europeocaneus Auge et Smith, 1997 из нижнего эоцена Бельгии.
Ниже описан новый ископаемый представитель семейства Agamidae. Находка происходит из местонахождения Цаган-Сайр (сухое русло р. Цаган-Сайр, урочище Цаган-Хушу, Нэмэгэтинская котловина) в Южной Монголии, где представлены отложения пачки жигден (свита наран), имеющей делювиально-пролювиальный генезис. Образец обнаружен в 1979 г. Южногобийским отрядом Совместной Советско-Монгольской палеонтологической экспедиции. В Цаган-Сайре найдены также разнообразные позднепалеоценовые тетраподы, в основном млекопитающие (Бадамгарав, Решетов, 1985; Lopatin, 2006).
Первая ящерица из Цаган-Сайра – Tsagansaurus nemegetensis Alifanov, 2018 – описана недавно в семействе Parasaniwidae (Platynota, Anguimorpha). Это самый поздний представитель этой группы на территории Центральной Азии (Alifanov, 2018). Впоследствии парасанивиды здесь исчезают, уступив место более крупным и, видимо, более продвинутым Varanidae рода Saniwa.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ПОДОТРЯД IGUANIA
ИНФРАОТРЯД AGAMOGNATHA
МИКРООТРЯД PACHYGLOSSA
СЕМЕЙСТВО AGAMIDAE SPIX, 1825
Род Epileolis Alifanov, gen. nov.
Название рода – анаграмма от родового названия Leiolepis.
Типовой вид – Epileolis reshetovi sp. nov.
Диагноз. Короноидный отросток зубной кости высокий и широкий в основании. Супраангулярный отросток расположен на уровне середины высоты заднего края. Нижний край зубной кости не утолщен. Фланг субдентального гребня относительно узкий, без окципитального расширения. Клыкообразных зубов семь; они постепенно увеличиваются по размеру назад, меняя ориентацию их длинной оси по отношению к субдентальному гребню от рострального наклона у наиболее передних и окципитального наклона у наиболее задних зубов. Окципитально клыковидные зубы довольно крупные. Пренатальных заклыковых зубов четыре, все они трехвершинные и оторваны от субдентального гребня. Первые три зуба увеличены в окципитальном направлении, имеют зауженное основание. Последний зуб пренатальной серии уменьшен в размерах. Между зубами пре- и постнатальной серии выражен промежуток. Зубов постнатальной серии четыре, их боковые вершинки расположены асимметрично и прижаты к центральному конусу. По высоте зубы этой серии больше, чем по ширине их продольного диаметра в основании. Контур нижней части постнатальных зубов округлый. Последние два зуба касаются субдентального гребня.
Видовой состав. Типовой вид.
Сравнение. От большинства известных современных и ископаемых Agamidae новый род отличается высоким и широким в основании короноидным отростком, относительно низким расположением супраангулярного отростка зубной кости, наибольшим числом и крупными размерами “клыков”. От многих Agamidae (Agaminae) он отличается отсутствием утолщения нижнего края зубной кости, асимметричной трехвершинностью заклыковых зубов, незначительными размерами последнего зуба пренатальной серии, небольшим числом и заужением оснований зубов этой же серии, развитием диастемы между зубами пре- и постнатальной серии, а также немногочисленностью зубов пренатальной серии. От Anhuisaurus Hou, 1974 из позднего палеоцена Китая новый род дополнительно отличается относительно более узким флангом субдентального гребня зубной кости, более высокими постнатальными зубами, разным углом наклона передних и задних клыковидных зубов, а также небольшими и прижатыми к центральной вершинке боковыми зубчиками.
Замечание. Филогенетически новый род близок к Anhuisaurus Hou, 1974 из позднего палеоцена Китая. Две формы сближают большое число и относительно крупные размеры зубов клыкообразной серии, трехвершинность постклыковых зубов, небольшое число и заужение оснований пренатальных зубов, их значительный отрыв от субдентального гребня, а также уменьшенный размер последнего зуба этой же серии и округлый контур основания зубов постнатальной серии.
Epileolis reshetovi Alifanov, sp. nov.
Leiolepidae gen. et sp. nov.: Алифанов, 2000, с. 79.
Название вида в честь палеотериолога В.Ю. Решетова, автора находки.
Голотип – ПИН, № 4758/3, левая зубная кость c поврежденными клыковидными зубами и зубами постнатальной серии; Монголия, Южногобийский аймак, местонахождение Цаган-Сайр; верхний палеоцен, свита Наран, пачка Жигден.
Описание (рис. 1). Зубная кость вытянутая, слабо расширяется окципитально. Высота короноидного отростка равна половине высоты остальной части окципитального края зубной кости, а его ширина в 1.5 раза больше ширины самого крупного зуба постнатальной серии. Ангулярный отросток зубной кости либо не развит, либо разрушен. На лабиальной стороне кости в ее передней половине выражено пять–шесть относительно мелких губных отверстий. Фланг субдентального гребня не расширен, составляет половину от высоты зубной кости в ростральной части и более четверти высоты от ее средней части. На лабиальной стороне зубной кости расположен сравнительно высокий гребень, который широкой дугой протягивается от короноидного отростка вниз к нижнему краю кости до середины зубного ряда. Супраангулярный отросток расположен низко и клиновидный по форме.
Рис. 1.
Зубная кость с зубами Epileolis reshetovi gen. et sp. nov., голотип ПИН, № 4758/3: а – лабиально, б – лингвально; Монголия, Южногобийский аймак, Нэмэгэтинская котловина, местонахождение Цаган-Сайр; верхний палеоцен, свита Наран, пачка Жигден. Длина масштабной линейки – 3 мм.
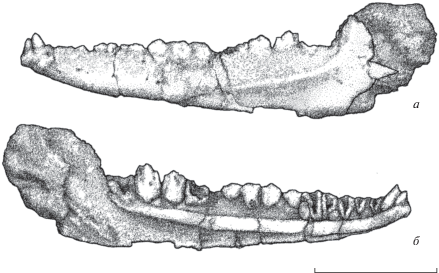
Вершины зубов клыкообразной серии, кроме первых двух, обломаны и демонстрируют наличие обширной внутренней полости (она, как и связь их оснований с субдентальным гребнем, позволяет отнести зубы к указанной серии). Первый клыкообразный зуб значительно уступает по размерам второму и наклонен рострально под углом близким к 60°. Основание зубов пренатальной серии (кроме последнего) заужено и удлинено, придавая зубам ромбовидную форму; остальные зубы имеют U-образный контур. Все зубы постнатальной серии крупнее (шире и выше примерно в 1.5 раза) пренатальных зубов. Они расположены плотно и касаются друг друга. Первый и последний зубы этой серии на образце разрушены, кроме их самой базальной части. Полнее остальных сохранился третий зуб. Он достигает своей нижней частью субдентального гребня. Последний зуб серии соприкасается с тем же гребнем довольно плотно.
Размеры голотипа в мм. Длина зубной кости – 9, ее высота в средней и задней части – 1 и 2, высота наиболее крупного верхнечелюстного зуба – 1.4, высота второго клыковидного зуба – 0.5.
Материал. Голотип.
Работа поддержана грантом РФФИ, № 16-05-00408.
Список литературы
Алифанов В.Р. Макроцефалозавры и ранние этапы эволюции ящериц Монголии. М.: ГЕОС, 2000. 126 с. (Тр. ПИН РАН. Т. 272).
Алифанов В.Р. Рarauromastyx gilmorei gen. et sp. nov. (Isodontosauridae, Iguania) – новая ящерица из позднего мела Монгольской Народной Республики // Палеонтол. журн. 2004. № 2. С. 87–92.
Алифанов В.Р. Новые акродонтные ящерицы (Lacertilia) из среднего эоцена Южной Монголии // Палеонтол. журн. 2009. № 6. С. 68–77.
Алифанов В.Р. Отряд Lacertilia // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Ред. Курочкин Е.Н., Лопатин А.В. М.: ГЕОС, 2012. С. 7–136.
Бадамгарав Д., Решетов В.Ю. Палеонтология и стратиграфия палеогена Заалтайской Гоби. М.: Наука, 1985. 104 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 25).
Каландадзе Н.Н., Раутиан А.С. Система млекопитающих и историческая зоогеография // Филогенетика млекопитающих / Ред. Россолимо О.Л. М.: Изд-во МГУ, 1992. С. 44–152 (Сб. трудов Зоол. музея МГУ. Т. 29).
Alifanov V.R. A new platynotan lizard (Parasaniwidae, Anguimorpha) from Late Paleocene of South Mongolia // Paleontol. J. 2018. V. 52. № 12. P. 1432–1435.
Böhme M. Herpetofauna (Anura, Squamata) and palaeoclimatic implications: preliminary results // Ann. Naturhist. Mus. Wien. 2007. Bd 108A. P. 43–52.
Dong L., Evans S.E., Wang Y. Taxonomic revision of lizards from the Paleocene deposits of the Qianshan Basin, Anhui, China // Vertebr. PalAsiat. 2016. V. 54. № 3. P. 243–268.
Estes R. Sauria terrestria, Amphisbaenia. Stuttgart, N.Y.: Fischer, 1983. 249 S. (Handbuch der Paläoherpetologie. T. 10A).
Gauthier J.A., Kearney M., Maisano J.A. et al. Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record // Bull. Peabody Mus. Natur. Hist. 2012. V. 53. № 1. P. 3–308.
Kalandadze N.N., Rautian A.S. Historical zoogeography of terrestrial tetrapods and new method of global palaeogeographical reconstructions // Evolution of the biosphere / Eds A.Yu. Rozanov, P. Vickers-Rich, C. Tassel. Launceston, 1997. P. 95–98 (Rec. Queen Victoria Mus., Art Gallery Launceston. № 104).
Lopatin A.V. Early Paleogene insectivore mammals of Asia and establishment of the major groups of Insectivora // Paleontol. J. 2006. V. 40. Suppl. 3. P. 205–405.
Moody S. Phylogenetic and historical biogeographical relationships of the genera in the Agamidae (Reptilia: Lacertilia). Ph. D. thesis. Univ. Michigan, 1980. 373 p.
Pyron A., Burbrink F.T., Wiens J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes // BMC Evol. Biol. 2013. V. 13. P. 1–93.
Rana R.S., Augé M., Folie A. et al. High diversity of acrodontan lizards in the Early Eocene Vastan Lignite Mine of India // Geol. Belgica. 2013. V. 16. № 4. P. 290–301.
Smith K.T., Schaal S.F.K., Wei S. et al. Acrodont iguanians (Squamata) from the Middle Eocene of the Huadian basin of Jilin province, China, with a critique of the taxon “Tinosaurus” // Vertebr. PalAsiat. 2011. V. 49. № 1. P. 69–84.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал