

Палеонтологический журнал, 2020, № 4, стр. 40-49
Позднекайнозойские остракоды семейства Ilyocyprididae Kaufmann Западно-Сибирской равнины
В. А. Коновалова *
Томский государственный университет
Томск, Россия
* E-mail: konovalova@ggf.tsu.ru
Поступила в редакцию 02.04.2019
После доработки 02.09.2019
Принята к публикации 21.01.2020
Аннотация
Представлены результаты ревизии сибирских представителей семейства Ilyocyprididae на базе коллекций неоген–четвертичных остракод, собранных Т.А. Казьминой, И.И. Тетериной и автором. Установлено, что остракоды семейства Ilyocyprididae представлены двумя родами – Ilyocypris Brady et Norman, 1889 и Fossilyocypris Schornikov et Krstić, 2004. Обсуждается видовой состав и стратиграфическое положение 12 таксонов данного семейства. Дано описание одного нового и четырех сомнительных видов.
ВВЕДЕНИЕ
Остракоды семейства Ilyocyprididae относятся к группе пресноводных ракушковых рачков и имеют широкое географическое и стратиграфическое распространение в отложениях позднего мезозоя и кайнозоя. Морфологическая пластичность раковин у видов данного семейства вызывает большие трудности при идентификации ископаемых рачков и нередко приводит к необоснованному выделению новых таксонов, или наоборот, сведению разных видов в объем одного (Попова-Львова, 1965, 1988; Казьмина, 1975; Дыкань, 2003; Дикань, 2006).
В позднем кайнозое Западной Сибири остракоды семейства Ilyocyprididae представлены двумя родами – Ilyocypris Brady et Norman, 1889 и Fossilyocypris Schornikov et Krstić, 2004 (Казьмина, 1975; Коновалова, 2012, 2015; Хазин, 2013; Тетерина, 2015, 2016; Хазин и др., 2017; и др.).
Род Ilyocypris Brady et Norman, 1889. Типовой вид – Ilyocypris gibba (Ramdohr, 1808). Остракоды этого рода характеризуются толстостенной субпрямоугольной раковиной с развитой ямчатой поверхностью и двумя поперечными центрально-дорсальными бороздами. Морфология раковин Ilyocypris отличается высокой внутривидовой изменчивостью, что затрудняет их правильную идентификацию. Систематика рода базируется на морфологии конечностей. Для диагностики раковины используются два признака: очертания раковины и скульптура. В настоящее время обсуждается вопрос об использовании третьего признака: краевые рябевидные структуры – “marginal ripplets”, расположенные в заднебрюшной области на внутреннем краевом валике левой створки (Mazzini et al., 2014). По данным автора, эти структуры не всегда наблюдаются на ископаемом материале из-за различной сохранности створок и, соответственно, их постоянство невозможно проследить во времени. Поэтому использовать данный признак в идентификации видов, вероятно, можно только на современном материале.
Род Fossilyocypris Schornikov et Krstić, 2004. Типовой вид – Fossilyocypris bella (Sharapova, 1961). В объем этого рода включены Ilyocyprididae с субпрямоугольными или субквадратными раковинами и с крупными ямками и ячейками на поверхности створок. Самые крупные ячейки расположены в постероцентральной части створок, у самок они более крупные, чем у самцов, хотя это различие не всегда соблюдается. Длина раковины у самок и самцов также разнится: у типового вида раковины самок относительно короткие, у самцов – удлиненные. Однако длина раковины не может рассматриваться в качестве диагностического признака рода. Так как к данному роду на сегодняшний день можно отнести 8 видов, то установить расширенный диагноз пока не представляется возможным. Дополнительной характеристике рода, конечно же, будет способствовать описание новых видов.
Целью работы является ревизия таксономического состава семейства Ilyocyprididae из плейстоценовых отложений Западной Сибири на основе детального морфологического изучения раковин и створок с помощью сканирующей электронной микроскопии.
МАТЕРИАЛ И МЕТОДЫ
Материалом для данной работы послужила авторская коллекция неоген–четвертичных остракод, собранная из обнажений и скважин, расположенных на территории центра и юга Западно-Сибирской равнины, и хранящаяся в лаборатории микропалеонтологии геолого-географического факультета Томского государственного ун-та (ЛМП ГГФ ТГУ) под № 119-5. Кроме того, были изучены рабочие коллекции неоген–четвертичных остракод (б/н) Т.А. Казьминой, хранящиеся в ИНГГ СО РАН, г. Новосибирск; коллекции четвертичных остракод №№ 41, 84, 132 М.И. Мандельштама и коллекция остракод № 786 из четвертичных отложений Зауралья Э.Д. Яскевич, хранящиеся во ВНИГРИ, С.-Петербург; рабочие коллекции четвертичных остракод, собранные из 16 скважин Томской обл. О.И. Саевой, хранящиеся в ЛМП ГГФ ТГУ; коллекции неоген-четвертичных остракод № БА-I О.Ю. Буткеевой и И.И. Тетериной, хранящиеся в геологическом музее СибГИУ (г. Новокузнецк).
При обработке образцов на микрофаунистический анализ использовалась традиционная методика (Практическое…, 1989). Для изучения морфологии раковин применялись стереомикроскопы Carl Zeiss Stemi 2000-C c фотокамерой Canon PowerShot A620 и Leica EZ 4D. Для уточнения строения отдельных скульптурных элементов раковины был использован сканирующий электронный микроскоп VEGA II LMU, установленный в Центре коллективного пользования (ЦКП) “Аналитический центр геохимии природных систем” ТГУ.
Для характеристики относительных размеров раковин остракод приняты следующие обозначения: небольшие – 0.5–1.0 мм, крупные – 1–1.5 мм. При описании использованы сокращения: D – длина раковины, Hmax – наибольшая высота, Hп.к – высота переднего конца, Hз.к – высота заднего конца, L – степень удлиненности раковины (L = D/Hmax).
Авторская коллекция хранится в лаборатории микропалеонтологии геолого-географического факультета Томского государственного ун-та (ЛМП ТГУ), № 119-5. Типовые экземпляры описанных видов хранятся в следующих организациях: Ин-т нефтегазовой геологии и геофизики Сибирского отделения Российской академии наук (ИНГГ СО РАН, Новосибирск), Всероссийский нефтяной научно-исследовательский геологоразведочный ин-т (ВНИГРИ, Санкт-Петербург), Музей Ин-та биологии моря ДВО РАН (Музей ИБМ ДВО РАН, Владивосток), Британский музей естественной истории (Англия, Лондон), Ун‑т Чукурова (Турция, Адана).
СТРАТИГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ВИДОВ СЕМЕЙСТВА ILYOCYPRIDIDAE В ОТЛОЖЕНИЯХ ВЕРХНЕГО КАЙНОЗОЯ ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ
В результате проведенных исследований в составе позднекайнозойских остракод Западно-Сибирской равнины на сегодняшний день установлено 10 видов (один из них новый) семейства Ilyocyprididae, и два таксона оставлено в открытой номенклатуре: Ilyocypris bradyi Sars, 1890, I. gibba, I. salebrosa Stepanaitys, 1959, I. biplicata (Koch, 1838), I. lacustris Kaufman, 1900, I. decipiens Masi, 1905, I. pustulata Konovalova, 2012, I. monstrifica (Norman, 1862) [ранее I. ex gr. tuberculata (Brady in Mandelstam et al., 1962)], Ilyocypris sp., Fossilyocypris sarizensis (Safak, Nasik et Senol, 1992) [ранее I. caspiensis (Negadaev, 1957) s. Kazmina, 1975, part.], F. gracilis sp. nov. [ранее I. ex gr. divisa (Klie, 1926) с мелкими ямками], F. aff. bella Scharapova, 1961 [ранее I. caspiensis (Negadaev, 1957) s. Kazmina, 1975, part.].
Для выяснения их стратиграфического значения необходимо проследить изменения в составе семейства Ilyocyprididae на протяжении всего позднего кайнозоя. Работа в этом направлении начата относительно недавно, но уже сейчас можно сделать некоторые выводы. В связи с изменением положения границы четвертичной системы до уровня 2.588 млн лет, изменился объем неогеновой и четвертичной систем, часть плиоценовых стратонов перешла в четвертичную систему. Поэтому необходимо подчеркнуть, что развитие остракод семейства Ilyocyprididae в данной работе рассматривается в рамках скорректированных региональных стратонов (рис. 1).
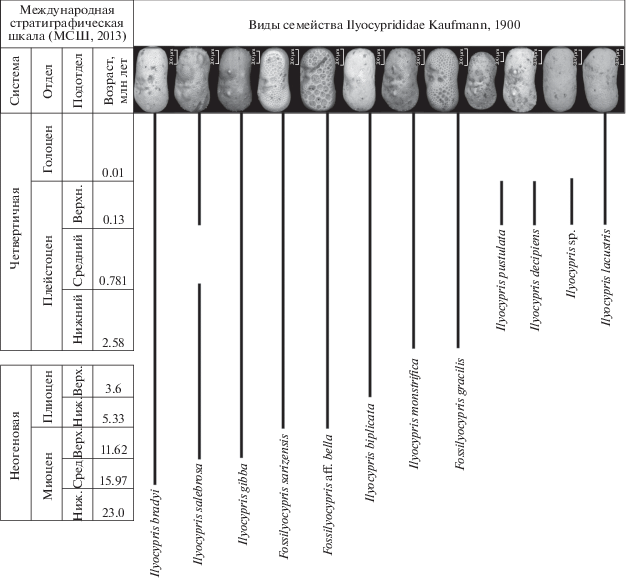
Первые единичные представители рода Ilyocypris (вид I. bradyi) появляются на территории Западной Сибири в отложениях таволжанской свиты среднего–верхнего миоцена (Казьмина, 1975, 1989; Зыкин, 2012). Вид I. bradyi характеризуется прямоугольной раковиной с двумя поперечными бороздами, центральной ямкой и ямчатой мезоскульптурой. Позднее, в новостаничной свите верхнего миоцена, по данным Казьминой, отмечены такие виды, как I. gibba и I. salebrosa, которые отличаются от I. bradyi хорошо развитыми буграми на раковине.
В плиоцене Западной Сибири происходит увеличение численности и разнообразия видов данного семейства, появляются первые представители рода Fossilyocypris. В раннем плиоцене (отложения рытовской свиты) появляются виды F. sarizensis и F. aff. bella, а в верхнем плиоцене (отложения аксорской свиты) впервые зафиксировано появление вида I. biplicata (Зыкин, 2012).
Первоначально Казьмина (1975) отнесла сибирские формы с удлиненной раковиной, определенные к настоящему времени как F. sarizensis, к виду I. caspiensis. К этому же виду она отнесла и короткие субквадратные раковины, которые принадлежат виду F. aff. bella (табл. V , фиг. 13 , см. вклейку), считая тех и других самцами и самками вида I. caspiensis. Описанный К.Н. Негадаевым-Никоновым (1957) вид, несмотря на внешнее сходство раковины с экземплярами с субквадратной формой, имеет терминальные хорошо развитые зубы замка, в отличие от слаборазвитых пластинчатых зубов у раковин рода Ilyocypris и принадлежит Limnocytheridae, роду Scordiscia Krstić et Schornikov, 1993. Кроме того, у раковин рода Ilyocypris замок правоваликовый, а у лимоноцитерид – левоваликовый. В этой же работе упоминается и вид Ilyocypris bella (nom. nud.), который встречается совместно с Scordiscia caspiensis (Negadaev, 1957). Вероятно, Негадаев-Никонов все же считал их разными видами.
Мезоскульптура сибирских форм характеризуется ямками и ячейками различной величины: в постеро-центральной части ячейки более крупные, чем на остальной поверхности. Такая скульптура соответствует описанию рода Fossilyocypris. По мнению Н. Крстич и Е.И. Шорникова, вид, описанный Казьминой как I. caspiensis (Negadaev) s. Kazmina, 1975, part. (Казьмина, 1975, табл. 18 , фиг. 5, 6 ), конспецифичен виду F. sarizensis. Автор настоящей работы оставляет это название для сибирской формы с удлиненной раковиной, тем более что видовое название “caspiensis” уже используется для другого ревизованного таксона, Scordiscia caspiensis (Negadaev, 1957) (Krstić et al., 2004; Krstić, 2006; Шорников, 2008). Для снятия вопросов по правомерности отнесения сибирского вида к виду F. sarizensis в настоящей работе приводится его изображение и описание (табл. V , фиг. 11, 14, 15 ). Что касается коротких субквадратных раковин Fossilyocypris aff. bella Scharapova, 1961, то тут ситуация неоднозначная. По мнению Шорникова (личн. сообщ.), изображения этой формы, с короткой субквадратной раковиной, у разных авторов довольно различны, и не факт, что они принадлежат одному виду. Поэтому нужны дальнейшие скоординированные исследования как зарубежных, так и отечественных ученых по изучению данных форм, которые, возможно, в будущем приведут к выделению новых видов семейства Ilyocyprididae.
Второй плиоценовый вид I. biplicata имеет сходство с видом I. bradyi, но отличается от него более нежной ямчатой мезоскульптурой и более выпуклой задней частью раковины. Согласно К. Мейшу (Meisсh, 2000), I. biplicata принимается как младший синоним I. gibba и рассматривается им как форма без боковых выступов на раковине. Однако Н. Сари и др. (Sari et al., 2012), а также Р. Фурман (Fuhrmann, 2012) считают, что различия в скульптуре и развитии спинного и брюшного края достаточны для отнесения их к разным видам. Автор также придерживается этого мнения и приводит изображение и описание сибирской формы, относимой им к виду I. biplicata (табл. V , фиг. 5, 8 ).
В плейстоцене роды Ilyocypris и Fossilyocypris продолжают свое развитие, а род Ilyocypris начинает занимать доминирующее положение в составе комплексов остракод, наряду с представителями кандонид и лимноцитерид. В отложениях кочковской свиты эоплейстоцена, наряду с уже существующими видами данного рода, появляются формы, которые Казьмина (1975) относила к I. ex gr. tuberculata. В настоящее время I. tuberculata Brady рассматривается как младший синоним вида I. monstrifica (Meisch, 2000; Mazzini et al., 2014; Meisch et al., 2019). Сибирская форма немного отличается от типичного вида отсутствием ярко выраженных острых краевых шипов на переднем и заднем концах раковины и слабо развитыми конусовидными буграми, из которых задний заострен и направлен назад (табл. V , фиг. 3, 6 ). Однако эти различия можно отнести к внутривидовой изменчивости. Находки этого вида в озерных голоценовых и современных отложениях Западно-Сибирской равнины показывают, что наряду с бесшипными экземплярами в одном местонахождении встречались экземпляры с развитыми на переднем или на заднем конце острыми небольшими шипиками. Наблюдалась и вариабельность заднего бугра: иногда он был наклонен назад, а иногда просто заострен. По мнению автора, это фенотипическая изменчивость вида, связанная с экологическими параметрами, в частности, с колебаниями солености озера. Вариабельность развития бугорков и шипов у I. monstrifica подтверждают данные и зарубежных исследователей (Mazzini et al., 2014).
На границе эоплейстоцена и неоплейстоцена (0.781 млн лет) происходит исчезновение морфотипа “I. postsalebrosa”, которое имеет, по мнению автора, не эволюционную, а экологическую причину. Вновь он появляется в отложениях верхнего неоплейстоцена, но уже в совершенно другой ассоциации видов.
Следующее незначительное изменение в таксономическом составе семейства Ilyocyprididae происходит на границе раннего и среднего неоплейстоцена (0.427 млн лет). В отложениях тобольской свиты появляются уплощенные формы илиоципридид с мелкоячеистой скульптурой, которые Казьмина относила к виду I. ex gr. divisa Klie (Архипов, 1971). В результате сравнительно-морфологического анализа этих форм автор настоящей работы выделяет их в новый вид F. gracilis (табл. V , фиг. 7, 9, 10, 12 ).
В отложениях верхнего неоплейстоцена в составе рода Ilyocypris происходят значительные изменения – появляются виды I. lacustris, I. decipiens, I. рustulata, I. sp. Вид I. lacustris характеризуется хорошо выраженным возвышением над глазом. Ilyocypris sp. представлен пока небольшим количеством экземпляров, но его отличительной особенностью является овальная, без видимых угловых переходов раковина. У I. decipiens хорошо выраженные тупо закругленные бугры, в отличие от сходного вида I. gibba, у которого эти бугры имеют заостренную форму. Вид I. pustulata характеризуется большими размерами, наличием мелких редких бугорков на поверхности, сильновыпуклой раковиной и сильноизогнутым очертанием створок (Коновалова, 2012, 2015).
В голоцене семейство Ilyocyprididae представлено всеми вышеперечисленными видами, кроме I. pustulata.
Таким образом, в позднем кайнозое Западно-Сибирской равнины в настоящее время семейство Ilyocyprididae представлено 12 таксонами, стратиграфическое значение которых предстоит выяснить в ближайшем будущем.
Ниже приводится описание нового вида Fossilyocypris gracilis и сибирских форм, относимых к видам F. sarizensis, Ilyocypris salebrosa, I. biplicata, I. monstrifica. Выбор описанных таксонов определяется необходимостью привести в соответствие их описание с учетом собственных и литературных данных.
ПОДОТРЯД CYPRIDOCOPINA
НАДСЕМЕЙСТВО CYPRIDACEA BAIRD, 1845
СЕМЕЙСТВО ILYOCYPRIDIDAE KAUFMANN, 1900
ПОДСЕМЕЙСТВО ILYOCYPRIDINAE KAUFMANN, 1900
Род Ilyocypris Brady et Norman, 1889
Ilyocypris salebrosa Stepanaitys, 1959
Табл. V , фиг. 1, 2
Ilyocypris salebrosa: Степанайтыс, 1959, с. 299, табл. 1 , фиг. 1; Казьмина, 1975, с. 47, табл. 18 , фиг. 3–4 .
Ilyocypris gibba (Ramdohr) var. tuberculata: Агаларова, 1961, c. 51, табл. 19 , фиг. 2 , а–в.
Ilyocypris postsalebrosa: Дыкань, 2003, с. 122, рис. 1.2 ; Дикань, 2006, с. 104, табл. 18 , фиг. 6–8; Коновалова, 2015, табл. 1 , фиг. 11 .
Голотип – ГПК Управления геологии и охраны недр Туркменистана, № 1-75; Западная Туркмения, северный борт Прибалханской депрессии; ранний плейстоцен, бакинский ярус (цит. по Казьминой, 1975). Информации о месте хранения в настоящий момент голотипа не имеется.
Описание. Раковина небольшая, прямоугольно-округленная. Наибольшая высота расположена в передней трети раковины и совпадает с высотой переднего конца. Наибольшая выпуклость расположена в постеро-дорсальной области. Спинной край неровный – прямой в передней и средней части и выпуклый в задней части. Кардинальные углы выражены. Брюшной край вогнутый, вогнутость смещена к заднему концу. Передний конец выше заднего, равномерно закругленный, вверху с небольшим уступом. Задний конец ниже переднего, плавно закруглен в заднебрюшной части.
Раковина имеет развитую скульптуру. В передней половине раковины расположены две узкие и глубокие депрессии, между которыми расположены два небольших круглых бугра. К спинному краю депрессии становятся шире. Ниже депрессий располагаются три субцентральных ямки. По краям депрессии расположены два широких крупных бугра, наибольший из которых расположен в постеро-дорсальной области. В постеро-вентральной части развит еще один бугор, который имеет овальную форму. На брюшной стороне створки расположено слабо выраженное широкое ребро, которое отделено от заднебрюшного бугра неглубокой впадиной. По краям ребра прослеживаются контуры двух маленьких расплывчатых бугорков. Поверхность створок, бугров и ребра покрыты четкими крупными пяти- шестигранными ячейками. На переднем и заднем концах раковины развиты редкие, среднего размера шипы.
Размеры в мм:
| №№ экз. | D | Hп.к | Hз.к |
|---|---|---|---|
| Голотип № 1-75 | 0.96 | 0.53 | 0.47 |
| ИННГ СО РАН, экз. № 23-1 | 0.95 | 0.53 | 0.42 |
| Экз. № 119-5/69 | 0.95 | 0.55 | 0.48 |
| Экз. № 119-5/69-1 | 0.97 | 0.52 | 0.47 |
Сравнение. От вида I. kaifengensis Lee, 1962 отличается округленной формой раковины, меньшей вогнутостью брюшного края, круглыми, а не заостренными буграми, наличием мелких округлых бугорков в передней части раковины и крупноячеистой поверхностью с четко очерченными, а не округлыми ячейками.
Замечания. Здесь следует сделать некоторые замечания относительно высокой изменчивости экземпляров, относимых к виду I. salebrosa.
Крстич (Krstić, 2006) считает, что этот вид является синонимом китайского вида I. kaifengensis. Как сообщает Крстич, два номинальных таксона I. kaifengensis Lee, 1962 [или 1959, согласно Кемпфу (Kempf, 1997)] и I. salebrosa Stepanaitys, 1959 были объединены позднее в один таксон с наименованием I. salebrosa (Guan et al., 1978, цит. по Krstić, 2006). Тем не менее, она считает, что название вида I. kaifengensis должно оставаться валидным. Кроме того, данный вид по-прежнему фигурирует в публикациях китайских исследователей как Ilyocypris kaifengensis Lee, 1962 (Huang, 1985; Wang et al., 2015). По устному сообщению китайского исследователя Даю Чжой (Dayou Zhai, Yunnan University), описание Ли в 1959 г. не было официально опубликовано и, возможно, типовые экземпляры уже утеряны. Согласно Международному кодексу зоологической номенклатуры (2004), неопубликованные виды не могут считаться валидными. Возможно, вид I. kaifengensis был переописан позднее в 1962 г., хотя никакой информации об этом в доступных источниках нет, поэтому вид I. salebrosa должен считаться старшим синонимом, и по праву приоритета сохранить свое название, так как был описан и опубликован Н.Е. Степанайтыс в 1959 г., а не в 1960 г., как ошибочно считается многими зарубежными исследователями (Smith et al., 2011; Martens, Savatenalinton, 2011; Karanovic, Lee, 2013; Meisch et al., 2019). Шорников любезно предоставил мне для сравнения SEM-фотографии вида I. kaifengensis из позднего плиоцена Башкирии (Южный Урал) (колл. М.Г. Поповой-Львовой). На ней изображен экземпляр с конусовидным, а не широким постеродорcальным бугром и иной формой раковины (табл. V , фиг. 4 ), что не соответствует изображению голотипа вида I. salebrosa (см. Сравнение).
Н.И. Дыкань (2003) выделила вид I. postsalebrosa Dykan, 2003, который несколько отличался от типичного вида наличием слабо выраженного брюшного ребра, образовавшегося в результате слияния двух бугров (их контуры прослеживаются в рельефе ребра). Элементы макроскульптуры (форма бугров по краям депрессий) и пропорции размеров, как сообщает Дыкань, сохраняются и у ювенильных форм (Дикань, 2006). Вопрос о половом диморфизме в данном случае не возникал, так как в одном образце ни разу не встречались формы с разной степенью развития бугров, хотя размеры раковин варьируют. По мнению Дыкань, вид Ilyocypris postsalebrosa Dykan является промежуточным звеном в филогенетическом ряду рода Ilyocypris: I. salebrosa–I. postsalebrosa–I. carinata Kovalenko (Дыкань, 2003; Дикань, 2006).
На основании сравнительно-морфологического анализа сибирской формы и морфотипа “I. postsalebrosa” автор настоящей работы считает, что экземпляры со слабо выраженным брюшным ребром соответствуют экземплярам морфотипа “I. postsalebrosa” (Казьмина, 1975, табл. 18 , фиг. 3 ; Дикань, 2006, табл. 18 , фиг. 6 ). Экземпляры с одним–тремя маленькими бугорками на брюшной стороне, которые, по мнению Дыкань, соответствуют виду I. salebrosa, также встречаются в отложениях плейстоцена Западной Сибири. По мнению Шорникова (2017), морфотипы “I. salebrosa”, “I. postsalebrosa” и “I. carinata” представляют собой онтогенетические варианты развития морфологических структур и являются одной из форм проявления изменчивости вида I. salebrosa.
Распространение. Плиоцен–голоцен; Россия, Западная Сибирь, Омская обл. (Исилькульский, Горьковский, Саргатский, Большеуковский, Омский р-ны), Алтайский край (Калманский, Ельцовский р-ны); Западный Туркменистан; Северный Казахстан; Азербайджан, Бакинский архипелаг; Украина, Одесская обл., Ренийский р-н; Турция; Китай; Япония.
Материал. Около 50 створок хорошей сохранности.
Ilyocypris biplicata (Koch, 1838)
Табл. V , фиг. 5, 8
Cypris biplicata: Koch, 1838, с. 21, фиг. 16 a, b .
Ilyocypris biplicata: Sars, 1928, с. 106, табл. 49 ; Бронштейн, 1947, с. 89, табл. 1 , фиг. 4 ; Мандельштам и др., 1962, с. 96, табл. 6 , фиг. 10 ; Lister, 1975, с. 19–20, табл. 2 , фиг. 5, 6 ; Fuhrmann, 2012, табл. 68 , фиг. 1a–f, 2a–d .
Информации о местонахождении голотипа нет, в работе К. Коха (Koch, 1838) дано только описание и изображение вида. В последующих работах других исследователей также дается только ссылка на работу Коха (Koch, 1838) и дается переописание или просто изображение оригинального материала (см. синонимику). Оригинал – ЛМП ТГУ, № 119-5/81; Западная Сибирь, Томская обл., Западная скв. 8, обр. 18, гл. 14.0 м; поздний неоплейстоцен.
Описание. Раковина небольшая, субквадратная. Наибольшая высота расположена в передней трети раковины и немного превосходит 1/2 длины раковины. Спинной край прямой на левой створке и слегка выпуклый на правой створке, слабо наклонен к заднему концу. Кардинальные углы тупые, слабо выражены, передний более четкий, чем задний. Брюшной край вогнут посередине. Передний конец немного вытянут и равномерно закруглен, вверху немного скошен. Задний конец равномерно закруглен.
Две глубокие депрессии расположены в передней трети раковины над тремя субцентральными ямками. Депрессии сильно расширяются к спинному краю и образуют заметную впадину. Из-за этой впадины задняя половина раковины имеет приподнятый вид, особенно в средне-спинной части, и образует очерченную выпуклость. При виде сверху раковина сжата с боков, передний конец тупо заострен, а задний конец закруглен. Поверхность створок имеет тонкую ямчатую скульптуру.
Размеры в мм:
| №№ экз. | D | Hmax | Hп.к | Hз.к | L |
|---|---|---|---|---|---|
| Экз. № 119-5/55 | 0.90 | 0.47 | 0.47 | 0.45 | 1.9 |
| Экз. № 119-5/56 | 0.88 | 0.45 | 0.45 | 0.42 | 1.9 |
| Экз. № 119-5/81 | 1.00 | 0.52 | 0.52 | 0.50 | 1.9 |
| Экз. № 119-5/81-1 | 1.02 | 0.52 | 0.52 | 0.48 | 1.9 |
Сравнение. Форма раковины описываемого вида похожа на I. bradyi, но отличается тонкоямчатой поверхностью, отсутствием краевых зубчиков и плавными очертаниями створок. От I. gibba отличается отсутствием бугров на поверхности створок, меньшей длиной и большей высотой раковины, которая превышает 1/2 длины раковины, меньшей вогнутостью брюшного края.
Распространение. Плиоцен–голоцен; Западная Сибирь: Тюменская, Новосибирская, Томская обл.; Казахстан, Павлодарская обл.; Западный Туркменистан: Изат-Кули; Западная Европа: Германия, Турция.
Материал. Более 170 створок хорошей сохранности.
Ilyocypris monstrifica (Normann, 1862)
Табл. V , фиг. 3, 6
Cypris monstrifica: Normann, 1862, c. 45, табл. 3 , фиг. 4, 5 .
Ilyocypris monstrifica: Коваленко, 1976, с. 85, табл. 48 ; Meisch, 2000, с. 249, фиг. 105A-C ; Fuhrmann, 2012, c. 166, табл. 77 , фиг. 1a–f.
Лектотип – Британский музей естественной истории, экз. № 1911.11.8М3402; Англия, графство Лестершир; современный.
Описание. Раковина удлиненная, субпрямоугольная. Наибольшая высота расположена в передней половине створки. Спинной край неровный – прямой в передней части и немного выпуклый в задней части, наклонен к заднему концу. Брюшной край слегка вогнут посередине. При виде сбоку передний конец выше заднего, широко закруглен, в верхней части резко скошен. Задний конец ниже переднего, плавно закруглен. При виде сверху передний конец заужен, задний конец закруглен.
В передней половине раковины развиты две глубокие депрессии. Под депрессиями расположены три ямки, одна центральная крупнее и глубже, чем две другие. По краям депрессий расположены два конических бугра с широким основанием. Задний бугор наиболее развитый, высокий, гладкий, верхушкой направлен назад. Передний бугор менее развит, низкий, имеет гладкую верхушку. Третий бугор слабо выражен, расположен на брюшной стороне.
Поверхность створок имеет густую ямчатую скульптуру. На переднем и заднем концах раковины развиты мелкие редкие шипики.
Размеры в мм:
| №№ экз. | D | Hmax | Hп.к | Hз.к | L |
|---|---|---|---|---|---|
| Экз. № 119-5/71 | 0.88 | 0.50 | 0.50 | 0.45 | 1.76 |
| Экз. № 119-5/76 | 0.97 | 0.55 | 0.52 | 0.50 | 1.76 |
| Экз. № 119/5-76-1 | 0.88 | 0.50 | 0.50 | 0.47 | 1.76 |
Изменчивость. Встречаются раковины без ярко выраженных острых краевых шипов, иногда шипы на переднем или заднем конце слабо развиты. У заднего бугра варьирует степень наклона и заостренности верхушки.
Сравнение. От I. gibba отличается меньшей вогнутостью брюшного края, наличием мелких шипиков на обоих концах раковины, более узким основанием бугров, более зауженными концами раковины при виде сверху.
Распространение. Плейстоцен–голоцен; Западная Сибирь, Томская, Омская, Новосибирская обл., Алтайский край; Молдавия; Украина; Западная Европа: Англия, Франция, Германия, Венгрия, Словения, Чехия, Австрия, Италия.
Материал. Более 130 створок хорошей сохранности.
Род Fossilyocypris Schornikov et Krstić, 2004
Fossilyocypris sarizensis (Safak, Nasik et Senol, 1992)
Табл. V , фиг. 11, 14, 15
Ilyocypris sarizensis: Safak et al., 1992, с. 177, табл. 2 , фиг. 1–7 .
Fossilyocypris sarizensis: Krstić et al., 2004, с. 313, табл. 2 , фиг. 7, 8 .
Ilyocypris caspiensis (Negadaev): Казьмина, 1975, с. 45, табл. 18 , фиг. 5, 6 .
Голотип – Ун-т Чукурова, экз. № 1; Турция, пров. Кайсери, р-н Сарыз; плиоцен.
Описание. Раковина удлиненная, прямоугольно-округленная. Наибольшая высота и ширина находятся в передней части раковины. Спинной край прямой, брюшной вогнут ближе к переднему концу. Передний конец широко округленный, на правой створке заметно наклонен к брюшному краю. При виде сбоку задний конец более закругленный, чем передний. При виде сверху задняя половина раковины имеет сильно уплощенный вид, а передняя половина равномерно выпукла. Передний конец заострен, задний конец тупой.
В передней половине раковины расположены две узкие и очень глубокие депрессии. Субцентральная ямка также очень глубокая. Для данного вида характерно валикообразное окаймление по заднему краю, что создает небольшую вогнутость задней площадки. Валик имеет форму петли, которая начинается от центральной ямки, тянется вдоль спинного и заднего края, переходит на брюшную сторону и заканчивается снова у центральной ямки.
Поверхность створок с крупными ячейками в постероцентральной части и мелкими ямками в остальной части.
Половой диморфизм ярко выражен. Раковины самок более высокие и широкие. Раковины самцов удлиненные и низкие.
Размеры в мм:
| №№ экз. | D | Hmax | Hп.к | Hз.к | L |
|---|---|---|---|---|---|
| № 119-5/79 | 0.92 | 0.52 | 0.52 | 0.47 | 1.76 |
| № 119-5/82 | 0.82 | 0.47 | 0.47 | 0.40 | 1.74 |
| № 119-5/80 | 0.87 | 0.45 | 0.45 | 0.40 | 1.9 3 |
| № 119-5/80-1 | 0.90 | 0.47 | 0.47 | 0.42 | 1.91 |
Сравнение. От F. bella отличается наличием петлеобразного валика и наличием крупных ячеек только в постероцентральной части.
Распространение. Плиоцен–плейстоцен; Западная Сибирь, Новосибирская, Омская, Томская обл., Алтайский край; Сербия; Турция.
Материал. 370 створок хорошей сохранности.
Fossilyocypris gracilis Konovalova, sp. nov.
Табл. V , фиг. 7, 9, 10, 12
Fossilyocypris sp.: Коновалова, 2015, с. 85, табл. 1 , фиг. 14 .
Fossilyocypris cf. sarizensis: Коновалова, 2015, с. 85, табл. 1 , фиг. 12 .
Название вида gracilis лат. – нежный.
Голотип – ЛМП ТГУ, № 119-5/78, правая створка самки; Западная Сибирь, Омская обл., Старо-Солдатская скв. 20, обр. 14, гл. 13.0 м; поздний неоплейстоцен.
Описание. Раковина небольшая, толстостенная, прямоугольно-овальная, сильно уплощенная в задней части, полого спадает к переднему концу и спинному краю, менее полого к заднему концу и брюшному краю. Спинной край неровный, более выпуклый в задней части. Брюшной вогнут посередине. Наибольшая высота находится в передней трети раковины. Передний конец широко закруглен, немного скошен в верхней части. Задний конец ниже переднего, равномерно закруглен. При виде сверху раковина имеет слегка уплощенную постероцентральную часть и равномерно выпуклую переднюю половину. Передний конец заужен, задний конец закруглен.
В передней половине расположены две короткие поперечные депрессии, первая немного длиннее, чем вторая. Депрессии начинаются от спинного края, но не доходят до середины створки. Ниже второй депрессии располагается центральная ямка, чуть впереди ее – еще 2 маленькие ямки. По краям депрессий в средней части намечаются два слаборазвитых округлых бугра.
Створки имеют тонкую мелкоямчатую поверхность, за исключением постероцентральной части. Здесь расположены несколько ячеек среднего размера, упорядоченных в 3–4 ряда. У самцов ячейки более четко выражены, чем у самок.
На переднем крае раковины иногда наблюдаются густо расположенные мелкие сосочковидные образования. Порово-канальная зона узкая, пронизана редкими прямыми поровыми каналами.
Размеры в мм:
| №№ экз. | D | Hmax | Hп.к | Hз.к | L |
|---|---|---|---|---|---|
| Голотип № 119-5/78 | 0.80 | 0.45 | 0.43 | 0.38 | 1.8 |
| Паратип № 119-5/77 | 0.85 | 0.47 | 0.47 | 0.43 | 1.8 |
| Экз. № 119-5/72 | 0.78 | 0.42 | 0.40 | 0.38 | 1.8 |
Изменчивость. Варьирует длина и высота раковины. Возможно, более длинные и низкие раковины принадлежат самцам, а более короткие и высокие – самкам. Меняется характер ячеек в постероцентральной части. Иногда группа ячеек ярко выражена и ячейки имеют более четкие грани, а иногда ячейки более сглажены и группа из них выглядит расплывчато.
Сравнение. От F. sarizensis отличается отсутствием петлеобразного валика, крупных ячеек в постероцентральной части и укороченной раковиной. От F. bella отличается тонкой мезоскульптурой и плавными очертаниями раковины.
Замечания. По наличию более крупных ячеек в постероцентральной части данный вид можно отнести к роду Fossilyocypris. В отличие от типичного вида, задняя половина створок данной формы равномерно уплощена, а не выпуклая. Створки имеют сглаженную ямчатую мезоскульптуру и отсутствие валикообразного окаймления по сравнению с другими известными видами рода. Эти отличия дают основание для выделения нового вида.
Распространение. Западная Сибирь, Томская, Омская, Новосибирская обл., Алтайский край; неоплейстоцен–голоцен.
Материал. Около 200 створок хорошей сохранности.
ЗАКЛЮЧЕНИЕ
Изучены позднекайнозойские остракоды семейства Ilyocyprididae. В результате сравнительно-морфологического анализа и обзора публикаций по этой проблеме автор пришел к следующим выводам. Из верхнекайнозойских отложений Западно-Сибирской равнины были описаны виды семейства Ilyocyprididae с различной морфологией раковины, внесенные в состав одного рода Ilyocypris. Сибирские формы, о которых сообщалось ранее как I. caspiensis (Negadaev) s. Kazmina, 1975 и I. ex gr. divisa Klie, следует отнести к роду Fossilyocypris. Все экземпляры, относимые к этому роду, имеют уплощенную форму раковины в задней половине. Уплощенность раковины может рассматриваться как дополнительный родовой признак. Уточнена таксономическая принадлежность вида I. ex gr. tuberculata, который по своим морфологическим признакам отнесен к виду I. monstrifica. Приведены дополнительные таксономические признаки, на основании которых сделано переописание видов I. salebrosa и I. biplicata. На основании проведенной ревизии уточнено стратиграфическое положение изученных таксонов семейства Ilyocyprididae. Установлено, что наряду с широким стратиграфическим распространением большинства видов, есть группа видов, которая имеет довольно узкий стратиграфический интервал. Потенциал этих видов, вероятно, можно будет использовать в дальнейшем при стратиграфическом расчленении четвертичных отложений Западно-Сибирской равнины.
Список литературы
Агаларова Д.А., Кадырова З.К., Кулиева С.А. Остракоды плиоценовых и постплиоценовых отложений Азербайджана. Баку: Азгосиздат, 1961. 420 с.
Архипов С.А. Четвертичный период в Западной Сибири. Новосибирск: Наука, 1971. 329 с.
Бронштейн З.С. Фауна СССР. Ракообразные. Ostracoda пресных вод. Т. 2. Вып. 1. М.–Л.: Изд-во АН СССР, 1947. 339 с.
Дикань Н.I. Систематика четвертинных остракод Украïни (довiдник-визначник). Київ: IГН НАНУ, 2006. 430 с.
Дыкань Н.И. Эволюция вида Ilyocypris salebrosa Stepanaitys, 1959 на протяжении плиоцена–голоцена // Докл. НАНУ. 2003. № 9. С. 121–124.
Зыкин В.С. Стратиграфия и эволюция природной среды и климата в позднем кайнозое юга Западной Сибири. Новосибирск: ГЕО, 2012. 487 с.
Казьмина Т.А. Стратиграфия и остракоды плиоцена и раннего плейстоцена юга Западно-Сибирской равнины. Новосибирск: Наука, 1975. 108 с.
Казьмина Т.А. Неогеновые и четвертичные комплексы остракод юга Западной Сибири // Кайнозой Сибири и северо-востока СССР. Новосибирск: Наука, 1989. С. 66–71.
Коваленко А.Л. Современные остракоды бассейна Днестра. Кишинев: Штиинца, 1976. 180 с.
Коновалова В.А. Новые и малоизвестные виды остракод среднего неоплейстоцена – голоцена юго-востока Западно-Сибирской равнины // Палеонтол. журн. 2012. № 3. С. 21–28.
Коновалова В.А. Виды рода Ilyocypris Brady et Norman, 1889 из отложений плейстоцена Западной Сибири // Современная микропалеонтология: Сборн. трудов XVI Всеросс. микропалеонтол. совещ. Калининград, 2015. С. 85–88.
Мандельштам М.И., Маркова Л.П., Розыева Т.Р., Степанайтыс Н.Е. Остракоды плиоценовых и постплиоценовых отложений Туркменистана (справочник). Ашхабад: Изд-во АН Туркм. ССР, 1962. 288 с.
Международный кодекс зоологической номенклатуры. Изд. четвертое. М.: Т-во научн. изданий КМК, 2004. 223 с.
Негадаев-Никонов К.Н. К истории развития фауны остракод в четвертичных бассейнах Северного Прикаспия // Уч. зап. Кишиневск. гос. унив. Геол. сб. 1957. Т. 25. С. 41–52.
Попова-Львова М.Г. Остракоды из плиоценовых отложений Башкирского Предуралья // Кайнозой Башкирского Предуралья. Т. 2. Ч. 2. М.: Недра, 1965. С. 210–287.
Попова-Львова М.Г. Остракоды из опорных разрезов Башкирского Предуралья Чуй-Атасево и Горнова // Некоторые вопросы биостратиграфии, палеомагнетизма и тектоники кайнозоя Предуралья. Уфа: БНЦ УО АН CCCP, 1988. С. 24–60.
Практическое руководство по микрофауне СССР: Остракоды кайнозоя. Т. 3. Л.: Недра, 1989. 235 с.
Степанайтыс Н.Е. Новые виды остракод из неогеновых и четвертичных отложений Юго-Западной Туркмении // Тр. Ин-та геол. АН Туркм. ССР. 1959. Т. 2. С. 298–315.
Тетерина И.И. Остракоды плиоцена центральных и юго-западных районов Степного Алтая // Вестн. Сибирского гос. индустриального ун-та. 2015. № 3(13). С. 18–21.
Тетерина И.И. Остракоды миоцена Чуйской и Курайской впадин Юго-Восточного Алтая // Палеонтол. журн. 2016. № 3. С. 64–74.
Хазин Л.Б. Голоценовые остракоды юга Западно-Сибирской равнины и Северного Казахстана: эколого-таксономические ассоциации, климатостратиграфическая корреляция и палеогеографические связи. Автореф. дис. … канд. геол.-мин. наук. Новосибирск: Ин-т нефтегаз. геол. и геофиз. им. А.А. Трофимука СО РАН, 2013. 17 с.
Хазин Л.Б., Кривоногов С.К., Гусев В.А. Голоценовые остракоды озера Саргуль (Новосибирская область) // Фундаментальные проблемы квартера: итоги изучения и основные направления дальнейших исследований. Матер. X Всеросс. совещ. по изучению четвертичного периода. М.: ГЕОС, 2017. С. 448–449.
Шорников Е.И. Реликтовые остракоды в фауне оз. Кушмурун (Казахстан) // Новости палеонтологии и стратиграфии. Прил. к журн. “Геол. и геофизика”. 2008. Т. 49. Вып. 10–11. С. 484–488.
Шорников Е.И. Таксономические замечания в отношении остракод Понто-Каспийского бассейна // Палеонтол. журн. 2017. № 5. С. 56–63.
Fuhrmann R. Atlas quartärer und rezenter Ostrakoden Mitteldeutschlands // Altenburger naturwiss. Forsch. 2012. Bd 15. 320 s.
Huang B. Pleistocene Ostracoda from middle and lower reaches of Sanggan river valley and its geological significance // Mem. Nanjing Inst. Geol. and Palaeontol. Acad. Sin. 1985. № 21. P. 85–118 [in Chinese].
Karanovic I., Lee W. On the ostracod genus Ilyocypris, with description of one new species from Korea and the first report of males of I. bradyi (Crustacea: Ostracoda: Podocopida) // Proc. Biol. Soc. Washington. 2013. V. 126. № 1. P. 39–71.
Kempf E.K. Index and Bibliography of Nonmarine Ostracoda. Part 6, Index A (Suppl. 1) // Geol. Inst. Univ. Köln. Spec. Publ. 1997. № 109. P. 1–142.
Koch C.L. Deutschlands Crustaceen, Myriapoden und Arachniden: Ein Beitrag zur Deutschen Fauna. Heft 21. Spezies 16. Regensburg, 1838.
Krstić N. Pliocene ostracodes of the Paludinian beds in the Pannonian Plain, Serbian Pt. Beograd, 2006. 409 p.
Krstić N., Savic L., Markovic Z. et al. Some important ostracodes from Late Pliocene (Akchagylian) of the Mediterranean and Central and Eastern Europe // Boll. Soc. Paleontol. Ital. 2004. V. 43. № 1–2. P. 307–320.
Lister K.H. Quartenary freshwater Ostracoda from the Great Salt Lake Basin, Utah // Paleontol. Contrib. Univ. Kansas. 1975. Paper 78. P. 1–34.
Martens K., Savatenalinton S. A subjective checklist of the Recent, free-living, non-marine Ostracoda (Crustacea) // Zootaxa. 2011. № 2855. 79 p.
Mazzini I., Gliozzi E., Rossetti G., Pieri V. The Ilyocypris puzzle: a multidisciplinary approach to the study of phenotypic variability // Int. Rev. Hydrobiol. 2014. V. 99. P. 395–408.
Meisch C. Freshwater Ostracoda of Western and Central Europe // Subwasserfauna von Mitteleupora 8/3 / Eds. Schwoerbel J., Zwick P. Heidelberg: Spektrum Akad. Verlag, 2000. 522 p.
Meisch C., Smith R., Martens K. A subjective global checklist of the extant non-marine Ostracoda (Crustacea) // Eur. J. Taxon. 2019. № 492. 135 p.
Safak Ü., Nasik A., Senol M. Kayseri güneydoğusu (Sarız) Pliyosen ostrakod ve gastropod faunası (Pliocene ostracoda and gastropoda faunae of southeastern Kayseri (Sariz)) // Çukurova Üniv. Mühendislik ve Mimarlık Fak. Dergisi. Adana. 1992. V. 7. № 1. P. 171–195.
Sari N., Külköylüoğlu O., Akdemir D. First record and detailed description of the male of Ilyocypris inermis Kaufmann, 1900 (Crustacea, Ostracoda) // Turk. J. Zool. 2012. V. 36. № 4. P. 484–495.
Smith R., Janz H., Okubo I. Recent Cyprididae and Ilyocyprididae (Crustacea: Ostracoda) from Lake Biwa, Japan, including a summary of the lake’s ostracod fauna // Zootaxa. 2011. № 2874. P. 1–37.
Wang Y., Min L.-r., Dong J. et al. Sedimentary characteristics and stratigraphic division of Holocene series in Baiyang Dian, Hebei Province // Acta Geosci. Sin. 2015. V. 36. № 5. P. 575–582.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал