

Палеонтологический журнал, 2020, № 4, стр. 13-20
Новый позднепалеозойский род гастропод Alanstukella gen. nov. (Trochonematidae)
А. В. Мазаев *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: mazaev.av@mail.ru
Поступила в редакцию 13.05.2019
После доработки 08.07.2019
Принята к публикации 26.07.2019
Аннотация
Описан новый род гастропод – Alanstukella gen. nov., семейство Trochonematidae. Типовой вид, A. rossica (Stuckenberg, 1905), входит в немногочисленную группу самых крупных позднепалеозойских представителей класса Gastropoda. Показано, что A. rossica обладает педоморфными чертами. Несмотря на чрезвычайную редкость находок, вид имеет широкое географическое и стратиграфическое распространение: Восточно-Европейский кратон, Южное Предуралье и Ферганская долина; верхний пенсильваний–нижняя пермь (от базальной части касимовского яруса до базальной части сакмарского яруса).
ВВЕДЕНИЕ
Изучение описанного здесь материала было инициировано находками отпечатков очень крупных трохиформных раковин в рифогенных известняках Шахтау. Систематическое определение этих экземпляров из-за плохой сохранности вызывало затруднения, поэтому их описание не вошло в монографию, посвященную гастроподам Шахтау (Mazaev, 2019). После изучения подобных отпечатков из карбонатов верхнего пенсильвания и нижней перми Восточно-Европейского кратона в коллекциях ПИН РАН было установлено, что весь этот ископаемый материал принадлежит одному виду, который был описан А.А. Штукенбергом как Mourlonia rossica Stuckenberg, 1905. Кроме того, к этому виду относится экземпляр из нижнепермских отложений южного обрамления Ферганской долины (ассель, хр. Карачатыр), который был описан И.А. Поповой (1972) как новый подвид Amaurotoma komishani karaschatyrica.
Вид M. rossica был установлен Штукенбергом (1905) по единственному отпечатку из верхнего пенсильвания Самарской Луки. К сожалению, оригиналы гастропод к этой работе Штукенберга, которые хранились в Казанском ун-те, утрачены. Судя по карандашному изображению слепка, оригинал был представлен отпечатком фрагмента завитка. Из-за неполноты оригинала подшовная площадка на последнем обороте выглядит сильно наклоненной, а детали строения начальных оборотов, базальной части раковины и устья неизвестны (Штукенберг, 1905, табл. 12 , фиг. 19 ). С другой стороны, идентификация этого вида упрощается, благодаря специфическому облику раковины и характерной скульптуре, состоящей из многочисленных спиральных ребрышек и крупных бугорков, маркирующих периферию подшовной площадки.
Изученные экземпляры в основном представлены неполными отпечатками. Исключение составляет только один из них, который является самым мелким. Благодаря хорошей сохранности, на нем можно наблюдать морфологические изменения первых оборотов. Таким образом, диагностика этого вида складывается как пазл из сопоставления признаков, наблюдаемых на разрозненных отпечатках, которые успешно дополняют друг друга.
Автор благодарит Т.Б. Леонову (ПИН РАН) за прочтение рукописи и полученные им ценные замечания, а также А.А. Школина за продуктивное сотрудничество во время полевых работ в 1990-х гг. и переданный им ископаемый материал.
ОБСУЖДЕНИЕ
Причины, по которым Попова не привела сравнений описанного ею экземпляра с видом, установленным Штукенбергом, неизвестны (Попова, 1972, с. 91). Однако ее родовое определение, с учетом сохранности изученного экземпляра, представляется весьма правдоподобным. Сходство признаков M. rossica с некоторыми диагностическими признаками рода Amaurotoma вполне очевидно. При более детальном изучении описанного здесь материала оказалось, что особенности строения линий роста и скульптурных элементов M. rossica не укладываются в диагноз рода Amaurotoma, поэтому для его размещения предлагается новый род – Alanstukella gen. nov.
У всех видов Amaurotoma линии роста на подшовной площадке и боковой поверхности взрослых оборотов в той или иной мере синусообразно изогнуты, в то время как у Alanstukella rossica (Stuckenberg, 1905) линии роста почти прямые и лежат на разных частях оборота почти в одной плоскости. Такое строение линий роста характерно для ювенильных стадий развития телеоконха Amaurotoma, поэтому сохранение этого признака у A. rossica на дефинитивных стадиях может свидетельствовать о педоморфном происхождении этого вида.
Несмотря на относительно обширное географическое распространение (рис. 1), A. rossica – очень редко встречающийся вид. Во всех коллекциях, собранных автором, а также в коллекциях, собранных в Самарской Луке (Штукенберг, 1905) и в Южной Фергане (Лихарев, 1967, 1968; Попова, 1972), этот вид представлен единичными экземплярами. При этом коллекции из местонахождений Южной Ферганы и Восточно-Европейского кратона собирались в течение десятилетий. Редкость этого вида особенно подчеркивается его крупными размерами (трудно не заметить во время сбора материала).
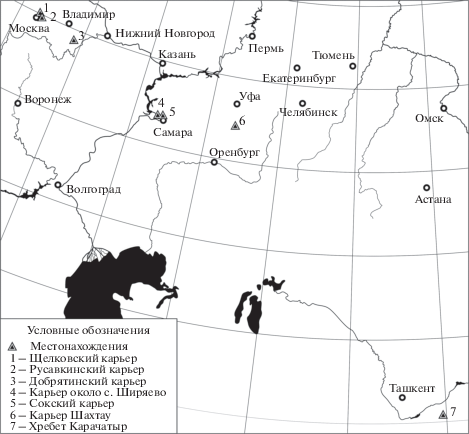
Интересно, что Штукенберг в описании экземпляра, который имел “4 см в длину”, указал на его относительно крупные размеры. Это еще раз подчеркивает тот факт, что гастроподы, в целом, – животные маленькие. Раковины позднепалеозойских гастропод, размер которых превышает 3.5 см, обычно попадают в размерный класс “крупные”. Поэтому находки экземпляров, достигающих 15 см, воспринимаются как нечто необычное (Kues, Duchene, 1990). Что же касается A. rossica, то он входит в число самых крупных видов позднепалеозойских гастропод. Диаметр самого крупного отпечатка, который был обнаружен в позднеассельских–раннесакмарских известняках Шахтау, составлял около 25 см. В настоящий момент единственный вид, которому A. rossica уступает в размерах, – это Murchisonia yabei Hayasaka, 1943 из поздней перми Японии (кэпитен), достигающий в длину 40 см (Hayasaka 1943; Hayasaka, Hayasaka, 1953). Родовое определение этого вида требует ревизии. Одновременно из этого местонахождения были описаны другие крупные гастроподы и бивальвии, в том числе – Pleurotomaria yokoyamai Hayasaka. Последний достигает в размерах 18×16 см (Hayasaka, 1943). Относительно недавно P. yokoyamai был предложен в качестве типового вида для рода Nipponomaria Asato et Kase, 2016.
Из-за сходства формы раковины, сечения оборотов и скульптуры Nipponomaria yokoyamai (Hayasaka, 1943) и Alanstukella rossica являются видами-двойниками. Однако их разделяет одно важное морфологическое отличие – Nipponomaria обладает узкой и глубокой селенизоной (Asato, Kase, 2016).
МАТЕРИАЛ
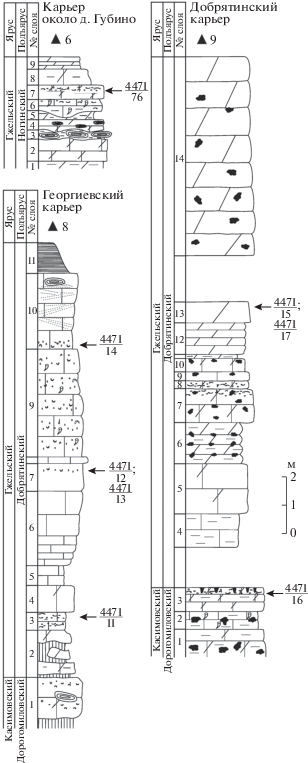
Описанный вид представлен в колл. ПИН РАН 11 экземплярами. Наиболее важный материал происходит из Добрятинского карьера. Номера местонахождений в разрезах карьеров Добрятино, Георгиевское (Владимирская обл.) и Губино (Московская обл.) были опубликованы ранее (Mazaev, 2011, рис. 7 ). К сожалению, на этапе редактирования в издательстве номера местонахождений были изменены и вышли в печать с ошибками. Поэтому в настоящей работе повторно приведены все три разреза с оригинальными номерами местонахождений (рис. 2).
Рис. 2.
Стратиграфическое положение местонахождений моллюсков в разрезах Георгиевского, Добрятинского карьеров и карьера около дер. Губино.
Самый крупный экземпляр описанного здесь вида наблюдался нами в рифогенных известняках Шахтау (местонахождение № 5538/1, экспедиция 2015 г.). Он был представлен отпечатком завитка в огромном блоке известняка, длиной более 10 м. К сожалению, этот блок был уничтожен карьерной техникой уже через день после того, как был обнаружен. Два подобных экземпляра, но меньших размеров, были найдены только через год во время третьей экспедиции. Оба экземпляра были обнаружены А.С. Бяковым (Северо-Восточный комплексный н.-и. ин-т ДВО РАН, Магадан). Один из них представлен ядром, второй – внешним отпечатком начальных оборотов с деталями скульптуры (экз. №№ 5538/11-44, 5538/11-45). Третий, самый крупный из имеющихся экземпляров, был обнаружен автором во время пятой экспедиции, осенью 2017 г. Его максимальный диаметр достигает 18 см (рис. 3). Известняки всех трех местонахождений приурочены к пограничным отложениям ассельского и сакмарского ярусов. Эти находки определяют самую верхнюю границу распространения вида.
Рис. 3.
Alanstukella rossica (Stuckenberg, 1905), экз. ПИН, № 5538/15-1, полный отпечаток и ядро, покрытые корками кристаллов кальцита и озокеритом; республика Башкортостан, Шахтау; нижняя пермь, пограничные отложения ассельского и сакмарского яруса.

Самый древний экземпляр этого вида был обнаружен А.А. Школиным в отвалах разработок Тимирязевского радиуса Московского метрополитена в 1988 г. Стратиграфический интервал, из которого происходит этот экземпляр, датируется суворовской свитой кревякинского подъяруса касимовского яруса. Экземпляр довольно крупных размеров. Внутреннее ядро несет отпечатки элементов скульптуры внешней поверхности раковины (рис. 4, ж). Такая специфическая сохранность объясняется растворением раковины, почти синхронным с процессом компакции осадка, и характерна для всех глинистых известняков Московского бассейна. Экземпляр деформирован, но обладает всеми признаками, необходимыми для определения его видовой принадлежности.
Рис. 4.
Alanstukella rossica (Stuckenberg, 1905), ×1: a–г – экз. ПИН, № 4471/17-38: а, б – ядро, внутренний слепок начала оборота: а – вид сбоку, б – вид с устья; в, г – латексный слепок: в – вид сбоку, г – вид снизу; д – экз. ПИН, № 4471/17-39, вид сбоку; Владимирская обл., Добрятинский карьер, гжельский ярус, добрятинский подъярус; е – экз. ПИН, № 4471/3-57, ядро; Московская обл., карьер Русавкино; гжельский ярус, добрятинский подъярус; ж – экз. ПИН, № 4471/20-120, ядро c отпечатком элементов скульптуры наружной поверхности; Москва, отвалы метро Тимирязевской линии; касимовский ярус, кревякинский подъярус.

Детали строения скульптуры, а также особенности онтогенетических изменений наблюдаются на экземплярах, собранных автором в 1991 г. в карьере Добрятино. Два крупных экземпляра представлены отпечатками завитков и происходят из базальной части гжельского яруса (добрятинский подъярус, местонахождение № 4471/17). На отпечатках хорошо сохранились элементы скульптуры, однако начальные обороты сильно потерты, устье отсутствует (рис. 4, в–д). Самые маленькие экземпляры представлены двумя более полными отпечатками. Оба экземпляра происходят из терминальной части касимовского яруса (дорогомиловский горизонт, местонахождение № 4471/16). В настоящее время это единственные экземпляры, на которых можно наблюдать строение плохо сохранившихся протоконхов и хорошо сохранившихся начальных оборотов (рис. 5, а–в).
Рис. 5.
Alanstukella rossica (Stuckenberg, 1905): а–в – экз. ПИН, № 4471/16-42, латексный слепок: а, б – вид с устья и вид снизу, ×5; в – плохо сохранившийся протоконх и начальные обороты, ×10; Владимирская обл., Добрятинский карьер; касимовский ярус, дорогомиловский подъярус; г – экз. ПИН, № 4471/4-36, латексный слепок; Московская обл., Щелковский карьер; гжельский ярус, добрятинский подъярус, ×2.5.

Морфология начальных и взрослых оборотов этого вида значительно изменяется в онтогенезе. Поэтому важным дополнением оказался отпечаток завитка средних размеров из Щелковского карьера Московской обл. (гжельский ярус, рис. 5, г).
Еще один экземпляр представлен крупным ядром (рис. 4, е), которое было найдено К. Лебедевым (Геологическая школа Московского государственного ун-та им. М.В. Ломоносова) в карьере Русавкино Московской обл., местонахождение № 4471/3. Форма и размерный класс этого ядра идентичны ядру из Шахтау (экз. № 5538/11-44).
Одиннадцатый из изученных экземпляров был обнаружен автором в ассельских доломитизированных известняках Сокского карьера (Самарская лука) в 2013 г. Экземпляр (№ 4919/36-1) представлен отпечатком фрагмента крупной раковины с хорошо сохранившейся скульптурой.
Материал хранится в Палеонтологическом музее им. Орлова, Палеонтологический ин-т им. А.А. Борисяка РАН (ПИН РАН), колл. №№ 4471, 4919 и 5538.
СЕМЕЙСТВО TROCHONEMATIDAE ZITTEL, 1895
Род Alanstukella Mazaev, gen. nov.
Название рода образовано из первых букв имени, отчества и фамилии Александра Антоновича Штукенберга – одного из основоположников Русской геологии.
Типовой вид – Mourlonia rossica Stuckenberg, 1905; Россия, Самарская Лука; беллерофоновый горизонт; верхний карбон, гжельский ярус.
Диагноз. Протоконх почти планиспиральный. Первые обороты телеоконха округлые в сечении. Боковая поверхность взрослых оборотов сверху и снизу ограничена отчетливыми килями. Подшовная площадка широкая. Умбиликальная и базальная поверхности разделены отчетливым килеватым перегибом. Поверхность раковины орнаментирована многочисленными спиральными шнуровидными ребрышками; первые взрослые обороты несут очень тонкие коллабральные ребрышки, форма которых совпадает с формой линий роста. Верхний киль на взрослых оборотах может маркироваться рядом крупных бугорков. Линии роста на подшовной площадке, боковой и базальной поверхностях почти прямые, лежат в одной плоскости, отчетливо наклонены прозоклинно. Устье голостомное, на внутренней поверхности раковины под дистальной частью верхнего киля имеется отчетливый, хорошо развитый канал.
Видовой состав. Типовой вид.
Сравнение. От Amaurotoma Knight, 1942 отличается прямыми линиями роста на подшовной площадке и боковой поверхности, развитием тонких коллабральных ребер на начальных оборотах, а также четким разделением базальной и умбиликальной поверхности отчетливым перегибом.
Alanstukella rossica (Stuckenberg, 1905)
Mourlonia rossica: Штукенберг, 1905, с. 96, табл. 12 , фиг. 19 .
Amaurotoma komishani karaschatyrica: Попова, 1972, с. 91, табл. 41 , фиг. 4 [non Amaurotoma komishani Licharew, 1967].
Голотип утрачен.
Описание (рис. 3–5). Очень крупная турбиниформная раковина.
Протоконх крупный, около 0.5 мм, почти планиспиральный. Три начальных оборота округлые в сечении, на третьем обороте боковая поверхность уплощенная. Поверхность оборотов покрыта примерно девятью тонкими шнуровидными спиральными ребрышками, межреберные промежутки уплощенные, в 1.5–2 раза шире толщины ребер. Швы глубокие канальчатые. На третьем обороте наблюдаются четкие линии роста: почти прямые или очень слабо прозоциртные, наклонены прозоклинно под углом к оси около 27°.
На четвертом обороте формируется широкая подшовная площадка, отделенная от боковой поверхности резким килем. Шов тонкий и четкий, мелкий. Поверхность подшовной площадки слабовогнута, около шва выпуклая, орнаментирована пятью или шестью шнуровидными спиральными ребрами. Их толщина примерно такая же, как и на предыдущем обороте; ширина межреберных промежутков заметно увеличивается, некоторые межреберные промежутки могут нести очень тонкое дополнительное ребро второго порядка. Появляются четкие шнуровидные коллабральные ребра. Их толщина меньше спиральных. Форма ребер совпадает с формой линий роста: прямые, прозоклинные, примыкают к шву под углом около 60°, непосредственно около шва резко изогнуты. Боковая поверхность оборота прямая или слабовогнутая, наклонена к оси раковины примерно под углом 17°, орнаментирована примерно так же, как и подшовная площадка, число спиральных ребер от восьми до девяти. Ширина межреберных промежутков постепенно увеличивается книзу. Коллабральные ребра на боковой поверхности прямые, наклонены под углом около 30°. Базальная поверхность в профиле почти прямая, отчетливо наклонена, отделена от боковой и умбиликальной поверхности резкими килями, орнаментирована семью или восемью массивными шнуровидными ребрами. Межреберные промежутки резко вогнутые, их ширина примерно сопоставима с толщиной ребер, немного уменьшается ближе к умбиликальной области. Умбиликальная область слабовыпуклая, орнаментирована шестью или семью шнуровидными спиральными ребрами, ближе к пупку их толщина заметно уменьшается. Межреберные промежутки широкие, уплощенные, ближе к пупку становятся узкими. Коллабральные ребра на базальной и умбиликальной поверхности отчетливые, но тоньше, чем на подшовной площадке и боковой поверхности оборота.
На последующих оборотах число основных спиральных ребер почти не меняется, ширина межреберных промежутков заметно увеличивается. Дополнительные ребра второго и третьего порядка появляются в различных вариациях на подшовной площадке, а также на боковой и базальной поверхности, однако исчезают на геронтической стадии. Толщина коллабральных ребер постепенно уменьшается до тонких, волосовидных. Начиная с пятого оборота, на их месте формируются очень резкие, почти канальчатые линии роста. Примерно с шестого или седьмого оборота на верхнем киле начинают формироваться крупные бугорки, незначительно вытянутые коллабрально. На последующих оборотах они увеличиваются в размерах, в плане округлые или незначительно вытянуты вдоль коллабральных ребер. Профили верхнего и нижнего килей становятся более пологими, отчетливый перегиб между базальной и умбиликальной поверхностями исчезает. На последнем обороте самых крупных экземпляров бугорки могут быть развиты в различной степени, межреберные промежутки становятся очень широкими и уплощенными. Устье голостомное, с коротким каналом, который расположен под верхним килем. Края устья тонкие, лежат почти в одной, резко скошенной плоскости. Колюмеллярная губа слабо арковидно изогнута, плавно переходит в базальную. Пупок сквозной, узкий. Форма линий роста на взрослых оборотах совпадает с линиями роста на первых оборотах.
Размеры в мм:
| Экз. № | Высота раковины | Макс. диаметр |
|---|---|---|
| 4471/3-57 | $ \gg $873.0 | 100.0 |
| 4471/4-36 | $ \gg $27.0 | >32.0 |
| 4471/16-35 | >12.5 | >13.7 |
| 4471/16-42 | 12.9 | 12.9 |
| 4471/17-38 | 61.4 | >61.2 |
Распространение. Касимовский и гжельский ярусы в Московском бассейне и на Окско-Цнинском вале; касимовский, гжельский и ассельский ярусы в Самарской Луке; ассельский и базальная часть сакмарского яруса в башкирских шиханах; ассельский ярус южного обрамления Ферганской долины.
Материал. Всего 11 экз.: местонахождение № 4471/3 – 1 экз.; мест. № 4471/4 – 1 экз.; мест. № 4471/16 – 2 экз.; мест. № 4471/17 – 2 экз.; мест. № 4471/20 – 1 экз.; мест. № 4919/36 – 1 экз.; мест. № 5538/11 – 2 экз.; мест. № 5538/15 – 1 экз.
Список литературы
Лихарев Б.К. Скафоподы и гастроподы – Archaeogastropoda (исключая подотряд Bellerophontina и подотряд Neritopsina) из верхнего палеозоя Южной Ферганы // Тр. Всес. н.-и. геол. ин-та. Нов. сер. 1967. Т. 116. Вып. 2. С. 1–115.
Лихарев Б.К. Скафоподы и гастроподы верхнего карбона и нижней перми Южной Ферганы. М.: Недра, 1968. 119 с.
Попова И.А. Каменноугольные гастроподы Южной Ферганы // Новые данные по фауне палеозоя и мезозоя Узбекистана. Ташкент: ФАН, 1972. С. 80–103.
Штукенберг А.А. Фауна верхнекаменноугольной толщи Самарской Луки // Тр. Геол. ком. Нов. сер. 1905. Вып. 23. С. 1–144.
Asato K., Kase T. Nipponomaria, a new pleurotomarioidean gastropod genus (Mollusca) from the Permian Akasaka Limestone, central Japan // Paleontol. Res. 2016. V. 20. P. 385–393.
Hayasaka I. On some Permian gastropods from Kinsyozan, Akasaka-Mati, Gihu Prefecture // Mem. Fac. Sci., Taihoku Imp. Univ. Ser. 3. 1943. V. 1. P. 23–48.
Hayasaka I., Hayasaka S. Fossil assemblages of molluscs and brachiopods of unusually large sizes from the Permian of Japan // Trans. and Proc. Palaeontol. Soc. Japan. N. S. 1953. V. 10. P. 37–44.
Kues B.S., Duchene H.R. The largest Late Paleozoic bellerophontid gastropod // J. Paleontol. 1990. V. 64. № 3. P. 478–479.
Mazaev A.V. Pennsylvanian gastropods of the suborders Murchisoniina Cox et Knight, 1960 and Sinuspirina Mazaev subordo nov. from the Central regions of the Russian Platform: morphology, taxonomy, and phylogeny // Paleontol. J. 2011. V. 45. № 12. P. 1533–1599.
Mazaev A.V. Lower Permian gastropods of Shakhtau (the border deposits of Asselian and Sakmarian Stages, southern Cisuralia) // Paleontol. J. 2019. V. 53. № 12. P. 1237–1345.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал