

Палеонтологический журнал, 2020, № 4, стр. 93-98
Первый ископаемый тоди (Aves: Todidae) на Кубе
Н. В. Зеленков a, *, С. Ф. Гонсалес b, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Национальный музей естественной истории Кубы
Гавана, Куба
* E-mail: nzelen@paleo.ru
** E-mail: sory@mnhnc.inf.cu
Поступила в редакцию 25.08.2019
После доработки 02.09.2019
Принята к публикации 25.10.2019
Аннотация
Тоди (Aves: Todidae) – эндемики Больших Антильских островов и одни из характерных компонентов современной фауны Кубы – до настоящего времени не были представлены в палеонтологической летописи острова. В статье описана первая для Кубы ископаемая находка представителя этого семейства. Коракоид из верхнего плейстоцена пещеры Эль Аброн (провинция Пинар-дель-Рио) общей морфологией и размерами схож с таковым современного кубинского тоди Todus multicolor, но отличается деталями строения и может представлять отдельный вид. Морфологическая обособленность позднеплейстоценового кубинского тоди также может свидетельствовать о морфологической вариабельности тоди, в большинстве случаев для птиц не характерной.
Современные тоди (семейство Todidae; отряд Coraciiformes) – очень мелкие тропические насекомоядные птицы с необычной внешней и внутренней морфологией, реликтовым распространением и загадочной эволюционной историей. Семейство включает один род Todus с пятью видами, распространенными исключительно на Больших Антильских островах. Два вида встречаются на Гаити и по одному – на островах Ямайка и Пуэрто-Рико. Кубинский тоди населяет Кубу, близлежащий остров Хувентуд и ряд расположенных поблизости более мелких островов.
Эволюционная история тоди изучена очень слабо. Традиционно семейство включали в отряд ракшеобразных (Coraciiformes), при этом еще ранние авторы (Murie, 1872; Sclater, 1872) считали ближайшими родственниками тоди в современной фауне момотов (семейство Momotidae) и зимородков (Alcadinidae). Тем не менее, многие крупные морфологи и систематики подчеркивали морфологическую уникальность тоди, в результате чего помещали этих птиц в отдельный отряд (Todiformes или Todi; Forbes, 1882; Lowe, 1948; Stresemann, 1959). В. Форбс (Forbes, 1882) считал тоди возможным морфологическим прототипом всех современных птиц из группы Anomalogonatae (немонофилетическая группировка, выделенная по отсутствию m. ambiens в задней конечности и включающая Strigiformes, Caprimulgiformes, Apodiformes, Coliiformes, Trogoniformes, Coraciiformes, Piciformes и Passeriformes; Beddard, 1898). Начиная с конца XX в. (Feduccia, 1977), тоди чаще всего объединяют в составе одного надсемейственного таксона (обычно называемого Alcedinoidea или Alcediniformes) вместе с момотами и зимородками в составе отряда Coraciiformes (см. Cracraft, 2013). При этом тоди, по современным молекулярным данным, являются наиболее базальными представителями этой группы (Prum et al., 2015).
Морфологическая обособленность тоди, отмеченная многими авторами, несомненно, свидетельствует о давней дивергенции этой группы. Молекулярные данные в сочетании с ископаемыми калибровками датируют дивергенцию тоди относительно других Alcedinoidea эоценом (от раннего до позднего; Prum et al., 2015; Claramunt, Cracraft, 2015). Заметное сходство с современными тоди в строении клюва характеризует раннеэоценовых Quasisyndactylus из нижнего эоцена Германии – вероятно, стволовых представителей Alcedinoidea (Mayr, 2004). Непосредственно к семейству Todidae относят вымерший род Palaeotodus с тремя видами, известными из нижнего олигоцена Вайоминга (США) и верхнего эоцена–нижнего олигоцена Германии и Франции (Olson, 1976; Mourer-Chauviré, 1985; Mayr, Knopf, 2007; Mayr, Micklich, 2010). Palaeotodus отличаются от современных тоди, в частности, пропорционально большими передними конечностями, что позволяет предполагать лучшие летные способности этих птиц по сравнению с современными Todidae (Olson, 1976). Представители Palaeotodus (кроме одного вида) также характеризуются более крупными размерами.
История заселения Todidae островов Карибского бассейна остается неясной, поскольку, начиная с верхнего олигоцена, представители этой группы практически полностью выпадают из палеонтологической летописи. Единственное исключение – два современных вида тоди (Todus subulatus Gray, 1847 и T. angustirostris Lafresnaye, 1851), определенные из четвертичных (по-видимому, верхнеплейстоценовых) отложений пещеры Cerro de San Franciso в Доминиканской Республике (Bernstein, 1965). Расхождение этих видов приурочивают к эпизоду существования двух островов на месте современного острова Гаити в плио-плейстоцене (Pregill, Olson, 1981). Тот факт, что в современной фауне тоди представлены морфологически очень близкими видами (иногда даже трактуемыми в рамке категории “superspecies”), свидетельствует о том, что радиация представителей рода Todus должна была иметь место в недалеком прошлом: не древнее раннего плейстоцена в современной трактовке (Bond, 1963; Kepler, 1977). Таким образом, эта группа птиц, будучи одной из характерных и эндемичных для Антильских островов, может иметь в этом регионе относительно недавнюю историю. В соответствии с данной концепцией предки тоди, уже обособившиеся от ведущей к момотам филогенетической линии, заселили Большие Антильские острова, как и многие другие представители фауны Карибов, из Центральной Америки – через полуостров Юкатан, который во времена падения уровня моря в плейстоцене был отделен от Кубы менее широкой водной преградой (Bond, 1963; Olson, 1976; Kepler, 1977; Pregill, Olson, 1981). При этом кубинский тоди представляется (Kepler, 1977) наиболее близким к предковой форме, давшей начало остальным видам – в первую очередь, сестринскому гаитянскому T. angustirostris (Overton, Rhoads, 2004).
Позднеплейстоцен–раннеголоценовая авифауна Кубы, предшествовавшая масштабному вымиранию позвоночных в Карибском бассейне в голоцене (Tyrberg, 2009; Cooke et al., 2017; Upham, 2017), существенно отличалась от современной и включала значительное число вымерших или ныне не встречающихся на острове видов (Orihuela, 2019). Сходная ситуация выявлена и для других островов Вест-Индии, в частности, Багамских (Steadman, Franklin, 2015). Особенно примечательна находка в четвертичных отложениях Кубы и о-ва Хувентуд мелких плохо летающих кричащих воробьиных Scytalopus – представителей южноамериканской группы, полностью отсутствующей в настоящее время на Карибах (Olson, Kurochkin, 1987). При этом важно отметить, что для завершающего интервала максимума последнего оледенения Кубы реконструируются самые низкие температуры (возможно, до 10°С ниже позднеголоценовых) и самый засушливый климат в этом регионе за последние 82 тыс. л. (Warken et al., 2019). Позднеплейстоценовые климатические условия ставят вопрос о сохранении в это время на Кубе отдельных реликтовых представителей тропической биоты, таких как тоди, которые до настоящего времени не были отмечены в палеонтологической летописи острова. В данной работе описана первая ископаемая находка тоди на Кубе – коракоид из верхнего плейстоцена (~завершающая фаза последнего ледникового максимума; см. ниже) пещеры Эль Аброн (провинция Пинар-дель-Рио), размерами и общей морфологией сходный с таковым современного кубинского тоди Todus multicolor, но отличающийся рядом деталей строения и таким образом, скорее всего, представляющий другой вид.
Описанная в этой работе находка – первое доказательство сосуществования представителей рода Todus в позднеплейстоценовых фаунах Кубы, но для установления точного систематического положения ископаемой формы необходимо сравнение с бóльшим разнообразием современных представителей рода (сравнение проводилось со скелетом T. multicolor). Отличия позднеплейстоценового кубинского тоди от современного могут быть интерпретированы двояко: либо позднеплейстоценовая фауна тодиевых Кубы отличалась от сегодняшней (например, включала большее разнообразие видов), либо тоди (все или только кубинские) характеризуются заметной морфологической изменчивостью или эволюцией на отрезке от позднего плейстоцена до современности. Вторая интерпретация, хотя и представляется возможной, но все же весьма необычна для птиц, поскольку современные виды (или комплексы близких видов) обычно неотличимы от ныне живущих, уже начиная с раннего–среднего плейстоцена (Зеленков, 2013, 2014; Zelenkov, 2016).
Заметная морфологическая эволюция не способных к длительным перелетам тоди могла бы быть объяснена существенными перестройками ландшафтно-климатических обстановок на рубеже плейстоцена и голоцена (климатическими факторами объясняется значительная часть вымираний позвоночных на Багамах в этот же временной промежуток; Steadman et al., 2015). В то же время сам факт присутствия тоди в позднеплейстоценовых фаунах не может служить индикатором каких-либо определенных экологических условий, поскольку современные тоди крайне эврибионтны. В частности, кубинский тоди населяет различные стации: от приморских пляжных порослей морского винограда Coccoloba unifera и ксерофитных кустарниковых зарослей до высокогорных (до 2840 м над уровнем моря) дождевых лесов (Kepler, 1977). Не исключено, что подобная эврибионтность может быть связана с большей морфологической пластичностью, в частности, выражающейся в индивидуальной изменчивости скелета – наподобие той, что выявлена для речных уток (Zelenkov, 2019).
ДАННЫЕ О МЕСТОНАХОЖДЕНИИ
Пещера Эль Аброн (22°40′ с.ш., 83°28′ з.д.) располагается в Западной части острова Куба в провинции Пинар-дель-Рио (рис. 1). Раскопки в пещере проводились сотрудниками департамента палеогеографии и палеобиологии Национального музея естественной истории Кубы в Гаване (далее MNHN), где и хранится описываемый материал. В процессе раскопок были вскрыты плейстоценовые отложения глубиной до 2.85 м, среди которых было выделено 9 слоев, некоторые из которых содержат богатый остеологический материал (подробнее о геологии местонахождения: Suárez, Díaz-Franco, 2003, 2011). Считается, что накопление костей позвоночных в пещере Эль Аброн связано с жизнедеятельностью сипух (Suárez, Díaz-Franco, 2003, 2011; Suárez, 2004). Для слоя VII, из которого происходит описываемый в настоящей статье фрагмент, по кости вымершей сипухи Tyto noeli Arredondo, 1972 была получена радиоуглеродная датировка – 17406 ± 161 л. н. (Suárez, Díaz-Franco, 2003, 2011).
Фауна местонахождения включает несколько видов птиц, в том числе вымершие Falco kurochkini Suarez et Olson, 2001, Capella sp., Tyto noeli, Siphonornis daiquiri Olson, 1985 и ряд современных (Suarez, 2004; Suarez, Díaz-Franco, 2011), а также несколько видов млекопитающих (Díaz-Franco, 2001; Suarez, Díaz Franco, 2003; González, 2015). Из разнообразной герпетофауны местонахождения пока описаны только амфисбены (Syromyatnikova et al., 2020).
ОПИСАНИЕ МАТЕРИАЛА
Полный левый коракоид (экз. MNHN, № 73.5395; рис. 2) из слоя VII местонахождения Эль Аброн имеет довольно хорошую сохранность – отломаны только наружные части латерального и медиального отростоков extremitas sternalis, форма которых у большинства птиц подвержена индивидуальной вариации; несколько повреждена медиальная вершина processus acrocoracoideus. Общие очертания коракоида, как и у других представителей Alcedinoidea и в отличие от остальных Coraciiformes, изящные, стержень тонкий и вытянутый, processus acrocoracoideus развит умеренно, имеет шишковидную форму при виде с вентральной стороны и несильно выступает медиально относительно стержня. Как и у Momotidae и Alcedinidae, processus acrocoracoideus ориентирован краниомедиально и несколько наклоняется вентрально. У Momotidae processus acrocoracoideus значительно более мощный и имеет краниокаудально широкое основание. У Alcedinidae общая форма processus acrocoracoideus сходна с таковым у ископаемого экземпляра и современных Todus, но при этом данный отросток сращен с processus procoracoideus.
Рис. 2.
Тоди (Aves: Todidae) из верхнего плейстоцена Кубы в сравнении с современным кубинским тоди (Todus multicolor): а, в – экз. MNHN, № 73.5395, полный левый коракоид; местонахождение Эль Аброн (провинция Пинар-дель-Рио), верхний плейстоцен; б, г – Todus multicolor Gould, экз. ПИН, № 1641-1, современный (Куба). Обозначения: con – углубление в каудальной части processus acrocoracoideus; cs – cotyla scapularis; es – extremitas sternalis; fah – facies articularis humeralis; fas – facies articularis sternalis; ib – impressio bicipitalis; ila – impressio lig. acrocoracohumeralis; lg – labrum glenoidale; li – linea intermuscularis; pa – processus acrocoracoideus; pp – processus procoracoideus. а, б – с дорсальной стороны; в, г – с вентральной стороны.
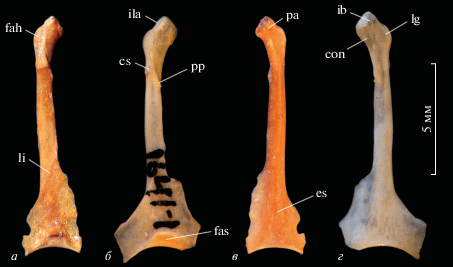
При виде с вентральной стороны processus acrocoracoideus заметно менее массивный в сравнении с таковым T. multicolor. В его центральной части, ближе к каудальному краю, присутствует неясное ребро, идущее от impressio bicipitis в каудолатеральном направлении и неявно переходящее каудально в вентральное ребро стержня. Латеральнее этого ребра развита вогнутость, окаймляющая несильно выступающее labrum glenoidale. У современного T. multicolor labrum заметно сильнее выпуклое, а указанное ребро и гребень в краниальной половине стержня, в который оно переходит, выражены заметно четче, чем у ископаемого экземпляра; на вентральной поверхности processus acrocoracoideus этот гребень заметно удален относительно каудального края отростка. Эта удаленность выражена за счет того, что каудальная часть основания processus acrocoracoideus у T. multicolor расширена (это обеспечивает массивность отростка) и дистально она переходит в гребень, медиально выступающий относительно sulcus supracoracoidei и оканчивающийся каудально на уровне вершины processus procoracoideus. У экз. MNHN, № 73.5395 этот гребень отсутствует. Кроме того, у T. multicolor медиальнее вышеупомянутого ребра на вентральной поверхности processus acrocoracoideus имеется неглубокое, но выраженное углубление (рис. 2, con), отсутствующее у ископаемого экземпляра. Impressio bicipitis у экз. MNHN, № 73.5395 сохранилось не очень хорошо, но, по-видимому, имело форму вытянутого прямоугольника, как у современного T. multicolor.
При виде с медиальной стороны processus acrocoracoideus выдается вентрально и формирует довольно острую вершину (более тупая у T. multicolor). Facies articularis clavicularis занимает вершину processus acrocoracoideus и частично ориентирована медиально. Sulcus supracoracoideus предствляет собой узкий косо ориентированный желобок вблизи processus procoracoideus, но краниальнее становится выпуклым, фактически переходя в край стержня. Impressio lig. acrocoracohumeralis слабо удлиненный, овальный и практически не изогнутый.
Facies articularis humeralis краниокаудально вытянутая и узкая, как у Todidae и Alcedinidae, неясно суживается каудально (у Momotidae короткая и широкая, особенно в своей каудальной части). Латеральная поверхность facies articularis humeralis вблизи контакта с cotyla scapularis несильно вогнутая; краниально facies слабо расширена, чем отличается от заметно расширенной facies articularis humeralis современного T. multicolor. Cotyla scapularis имеет субтреугольную форму, представляет собой продолжение facies articularis humeralis (как у Todidae и Alcedinidae) и неясно приподнято относительно последней в своей краниальной части. Каудальная часть cotyla заметно приподнята относительно прилегающей дорсальной поверхности стержня и отделена от нее невысоким “бортиком”. Дорсальная поверхность cotyla scapularis неявственно выпуклая в своей латеральной части и неявственно вогнутая – в медиальной; медиальный край cotyla каудально переходит в неявно выраженный рудиментарный (в отличие от Momotidae) processus procoracoideus, вершина которого заметно смещена каудально относительно каудального края cotyla. Краниальный скошенный край processus procoracoideus несет на своей медиальной поверхности сочленовную поверхность для акромиона лопатки. Дистально processus procoracoideus продолжается в длинный гребень, ориентированный медиально, идущий вдоль стержня и маркирующий его выдающуюся над медиальной поверхностью кромку вплоть до extremitas sternalis, как у Todidae. У современного T. multicolor этот гребень несколько сильнее выступает медиально, особенно в своей каудальной части. Foramen n. supracoracoidei отсутствует.
В extremitas sternalis хорошо развиты медиальный и латеральные гребни, представляющие собой крыловидные расширения. Медиальный гребень начинается от описанного выше медиального гребня стержня несколько дистальнее отхождения от последнего межмускульной линии, маркирующей краниальный край impressio m. sternocoracoidei. Точные очертания медиального гребня установить не удается, поскольку его наружная часть не сохранилась. Латеральный гребень начинается каудальнее медиального, и его наружный край представляет собой продолжение указанной мускульной линии. Наружная часть гребня также отломана. Дорсальная поверхность extremitas sternalis ровная или немного выпуклая в своей средней части и отчетливо вогнутая в области латерального гребня. Дорсальная поверхность медиального гребня вогнутая только в своей самой каудальной трети, ближе к angulus medialis. В краниальной части impressio m. sternocoracoidei имеется еще одна неясно выраженная мускульная линия, приближенная к краниальному краю отпечатка. Facies articularis sternalis при виде с дорсальной стороны имеет куполовидную форму с несколько зауженной краниальной вершиной, обрамленной отчетливым гребнем (crista dorsalis; Elzanowski et al., 2012) и удлиненными “языками” основания, распространяющимися латерально и медиально практически на весь сохранившийся каудальный край кости. У современного T. multicolor эти каудальные “языки” практически не выражены, и facies с дорсальной стороны имеет более расширенные очертания.
При виде с вентральной стороны extremitas sternalis выраженно выпуклая в своей краниальной части (продолжение выпуклости стержня) и уплощается дистально, медиально и латерально. Каудальный край равномерно вогнутый; facies articularis sternalis не распространяется на вентральную поверхность кости.
Размеры экз. MNHN, № 73.5395 (в мм): максимальная длина вдоль медиального края – 10.6; минимальная медиолатеральная ширина стержня – 0.9; минимальная дорсовентральная высота стержня – 0.6.
* * *
Настоящее исследование выполнено при финансовой поддержке РФФИ и СИТМА в рамках научного проекта №18-54-34004 (“Позднечетвертичные позвоночные Кубы: реликтовые фауны на пороге голоценового вымирания”). Авторы благодарны Э. Перес, Х. Пахон и Р. Рохас Консуегра (Гавана) за консультации и доступ к коллекциям (для первого автора), Е.В. Сыромятниковой (ПИН РАН) за помощь в работе с коллекциями и подготовке карты, а также рецензентам А.В. Пантелееву и А.Б. Савинецкому.
Список литературы
Зеленков Н.В. Зоологические проблемы четвертичной палеорнитологии // Зоол. журн. 2013. Т. 92. С. 1077–1087.
Зеленков Н.В. Этапы формирования современного таксономического разнообразия птиц (по данным палеонтологии) // Зоол. журн. 2014. Т. 93. С. 1173–1185.
Beddard F.E. The structure and classification of birds. L., N.Y., Bombay: Longmans, Green, and Co., 1898. 548 p.
Bernstein L. Fossil birds from the Dominican Republic // Quart. J. Florida Acad. Sci. 1965. V. 28. P. 271–284.
Bond J. Derivation of the Antillean avifauna // Proc. Acad. Natur. Sci. Philad. 1963. V. 115. P. 79–98.
Claramunt S., Cracraft J. A new time tree reveals Earth history’s imprint on the evolution of modern birds // Sci. Advances. 2015. V. 1: e1501005.
Cooke S.B., Dávalos L.M., Mychajliw A.M. et al. Anthropogenic extinction dominates Holocene declines of West Indian mammals // Ann. Rev. Ecol. Evol. Syst. 2017. V. 48. P. 301–327.
Cracraft J. Avian higher-level relationships and classification: Nonpasseriformes // The Howard and Moore complete checklist of the birds of the World. 4th ed. / Eds. Dickinson E.C., Remsen Jr. J.V. Eastbourne: Aves Press, 2013. P. XXI–XLI.
Díaz-Franco S. Estructura dental interna y modificación del diseño oclusal inferior en Boromys offella (Rodentia: Echimyidae) // Rev. Biol. 2001. V. 15. P. 152–157.
Elzanowski A., Bieńkowska-Wasiluk M., Chodyń R., Bogdanowicz W. Anatomy of the coracoid and diversity of the Procellariiformes (Aves) in the Oligocene of Europe // Palaeontology. 2012. V. 55. P. 1199–1221.
Feduccia A. A model for the evolution of perching birds // Syst. Biol. 1977. V. 26. P. 19–31.
Forbes W.A. On some points in the anatomy of the todies (Todidæ), and on the affinities of that group // Proc. Zool. Soc. London. 1882. V. 50. P. 442–450.
González S.F. La fauna mamíferos fósiles fel depósito paleontolígico “El Abrón” (nivel IX), Pinar del Río, Cuba. Unpubl. Thes. Havana: Univ. La Habana, 2015. 69 p.
Kepler A.K. Comparative study of todies (Todidae): with emphasis on the Puerto Rican tody, Todus mexicanus // Publ. Nuttal Ornithol. Club. 1977. V. 16. P. 1–190.
Lowe P.R. What are the Coraciiformes // Ibis. 1948. V. 90. P. 572–582.
Mayr G. New specimens of Hassiavis laticauda (Aves: Cypselomorphae) and Quasisyndactylus longibrachis (Aves: Alcediniformes) from the middle Eocene of Messel, Germany // Cour. Forsch. Senckenb. 2004. Bd 252. S. 23–28.
Mayr G., Knopf C.W. A tody (Alcediniformes: Todidae) from the early Oligocene of Germany // Auk. 2007. V. 124. P. 1294–1304.
Mayr G., Micklich N. New specimens of the avian taxa Eurotrochilus (Trochilidae) and Palaeotodus (Todidae) from the early Oligocene of Germany // Paläontol. Z. 2010. Bd 84. P. 387–395.
Mourer-Chauviré C. Les Todidae (Aves, Coraciiformes) des Phosphorites du Quercy (France) // Proc. Koninkl. Nederl. Akad. Wetensch. Ser. B. 1985. V. 88. P. 407–414.
Murie J. On the skeleton of Todus, with remarks as to its allies // Proc. Zool. Soc. London. 1872. P. 664–680.
Olson S.L. Oligocene fossil bearing on the origins of the Todidae and Momotidae (Aves: Coraciiformes) // Smithson. Contrib. Paleobiol. 1976. № 27. P. 111–119.
Olson S.L., Kurochkin E.N. Fossil evidence of a Tapaculo in the Quaternary of Cuba (Aves: Passeriformes: Scytalopodidae) // Proc. Biol. Soc. Wash. 1987. V. 100. P. 353–357.
Orihuela J. An annotated list of late Quaternary extinct birds of Cuba // Ornitol. Neotrop. 2019. V. 30. P. 57–67.
Overton L.C., Rhoads D. Molecular phylogenetic relationships based on mitochondrial and nuclear gene sequences for the Todies (Todus, Todidae) of the Caribbean // Mol. Phyl. Evol. 2004. V. 32. P. 524–538.
Pregill G.K., Olson S.L. Zoogeography of West Indian vertebrates in relation to Pleistocene climatic cycles // Ann. Rev. Ecol. Syst. 1981. V. 12. P. 75–98.
Prum R.O., Berv J.S., Dornburg A. et al. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing // Nature. 2015. V. 526. P. 569–573.
Sclater P.L. Observations on the systematic position of the genera Peltops, Eurylaemus, and Todus // Ibis. 1872. V. 24. P. 177–180.
Steadman D.W., Albury N.A., Kakuk B. et al. Vertebrate community on an ice-age Caribbean island // Proc. Natl Acad. Sci. 2015. V. 112. P. E5963–E5971.
Steadman D.W., Franklin J. Changes in a West Indian bird community since the late Pleistocene // J. Biogeogr. 2015. V. 42. P. 426–438.
Stresemann E. The status of avian systematics and its unsolved problems // Auk. 1959. V. 76. P. 269–280.
Suárez W. The enigmatic snipe Capella sp. (Aves: Scolopacidae) in the fossil record of Cuba // Carib. J. Sci. 2004. V. 40. P. 155–157.
Suárez W., Díaz-Franco S. A new fossil bat (Chiroptera: Phyllostomidae) from a Quaternary cave in Cuba // Carib. J. Sci. 2003. V. 39. P. 371–377.
Suárez W., Díaz-Franco S. Estudio paleontológico del depósito fosilífero El Abrón, Pinar del Río (Sinopsis de las aves fósiles de Cueva El Abrón, Pinar del Río, Cuba) // Biodiversidad Paleontológica del Archipiélago Cubano: Bases Cartográficas y Conservacionistas. Informe final del proyecto 022 AMA-CITMA; 074 MNHN. Havana: Mus. Natl Hist. Natur., 2011. 13 p.
Syromyatnikova E.V., Aranda E., Gonzalez S.F. The first fossil record of Cadea (Amphisbaenia, Cadeidae) and other amphisbaenian remains from the Upper Pleistocene of Cuba // J. Vertebr. Paleontol. 2020. e1729167. https://doi.org/10.1080/02724634.2019.1729167
Tyrberg T. Holocene avian extinctions // Holocene Extinctions / Ed. S.T. Turvey. N.Y.: Oxford Univ. Press, 2009. P. 63–106.
Upham N.S. Past and present of insular Caribbean mammals: understanding Holocene extinctions to inform modern biodiversity conservation // J. Mammal. 2017. V. 98. P. 913–917.
Warken S.F. Scholz D., Spötl Ch. et al. Caribbean hydroclimate and vegetation history across the last glacial period // Quatern. Sci. Rev. 2019. V. 218. P. 75–90.
Zelenkov N.V. Evolution of bird communities in the Neogene of Central Asia, with a review of the Neogene fossil record of Asian birds // Paleontol. J. 2016. V. 50. № 12. P. 1421–1433.
Zelenkov N.V. Variability in the postcranial skeleton of the European dabbling ducks (Aves, Anatidae): identifying nodes of strong and relaxed selection // J. Morphol. 2019. V. 280. Suppl. 1. P. S243.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал