

Палеонтологический журнал, 2020, № 5, стр. 3-19
Морфология, палеобиология и систематическое положение кембрийских зоопроблематик – мобергеллид
Ю. Е. Демиденко a, *, П. Ю. Пархаев a
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: juliad@paleo.ru
Поступила в редакцию 11.11.2019
После доработки 25.02.2020
Принята к публикации 25.02.2020
Аннотация
Проанализированы возможные варианты систематического положения кембрийских зоопроблематик сем. Mobergellidae. Впервые обсуждается предполагаемое родство склеритов мобергеллид с сипункулидами (Sipuncula) – типом морских червеобразных бентосных лофотрохозой, вероятно, родственных кольчатым червям. Предлагается интерпретация склеритов мобергелл как каудальных щитков сипункулид.
ВВЕДЕНИЕ
Палеонтологическая летопись раннего кембрия наряду с широко известными ископаемыми – археоциатами, трилобитами, брахиоподами – представлена множеством морфологически разнообразных, преимущественно мелких склеритов (отдельные элементы скелета), реже – склеритомами (комплект склеритов, принадлежащих одному организму), а также мелкими цельными скелетами (раковины, трубки). Все они описываются в литературе под общим названием – мелкораковинные ископаемые, или SSF (small shelly fossils) (Matthews, Missarzhevsky, 1975; Bengtson et al., 1990). Комплекс этих таксономически разнородных фоссилий – результат широко используемой методики препарирования кембрийских микроископаемых – извлечения скелетных элементов из образцов карбонатных пород путем растворения пород в уксусной кислоте, промывки нерастворенного осадка от глинистой фракции и последующего отбора из него интересующего материала. Значительная часть кембрийских мелкораковинных ископаемых – проблематики (Розанов и др., 1969; Matthews, Missarzhevsky, 1975; Rozanov, Zhuravlev, 1992; Dzik, 1994), т.е., таксоны неясного систематического положения, в силу отсутствия морфологически близких организмов среди современных беспозвоночных животных.
В последние годы наблюдается заметный прогресс в решении вопросов функциональной морфологии и интерпретации систематического положения многих групп кембрийских зоопроблематик, которые долгое время являлись загадочными. Например, после находок сочлененных склеритов (склеритомов) томмотиид родов Paterimitra и Eccentrotheca и изучения их морфологии и микроструктуры стенки склеритов, стало возможным поместить эти таксоны в основание ствола брахиопод. Также было показано, что ископаемые Microdyction могут рассматриваться как базальные представители стволовой группы онихофор (Skovsted et al., 2008, 2009, 2011, 2014; Balthasar et al., 2009; Murdock et al., 2012, 2014).
Однако, систематическое положение многих кембрийских таксонов, представленных в ископаемой летописи только склеритами, остается невыясненным. К таким группам относятся склериты представителей семейства Mobergellidae Missarzhevsky, 1989. Это семейство, выделенное В.В. Миссаржевским (1989) и условно отнесенное к типу Mollusca, первоначально включало в себя три рода – Mobergella Hedström, 1923, Brastadella Missarzhevsky, 1989 и Discinella Hall, 1872. К данной группе Миссаржевский (1989) отнес мелкие многослойные фосфатные “раковины” блюдцевидной или низкоконической формы с концентрическими линиями нарастания на наружной поверхности и парными мускульными отпечатками (далее – м.о.) – на внутренней (рис. 1; табл. I, II , см. вклейку). Миссаржевский (1989, с. 187) отмечал, что “раковины” мобергеллид по форме и характеру мускульных отпечатков сходны с раковинами некоторых представителей моллюсков класса моноплакофор, но отличаются от них химическим составом. Кроме Миссаржевского, мобергеллид к моллюскам относили еще некоторые палеонтологи (Hedström, 1923, 1930; Poulsen, 1932; Nyers, 1987; Conway Morris, Chapman, 1997); реже эти проблематики рассматривались как створки фосфатных брахиопод (Hall, 1872; Moberg, 1892; Dzik, 2010). Большинство же авторов склонялись к оперкулярной гипотезе, полагая что склериты мобергеллид являются крышечками хиолительминтов (Billings, 1871; Walcott, 1886, 1890; Knight, 1941; Lochman, 1956; Fisher, 1962) или неизвестного трубчатого организма (Bengtson, 1968; Rozanov, Zhuravlev, 1992; Topper, Skovsted, 2017; Skovsted, Topper, 2018). В данной статье впервые обсуждается возможность отнесения мобергеллид к сипункулидам и предлагается интерпретация их склеритов как щитков сипункулид.
Рис. 1.
Морфология склеритов видов мобергеллид: а–в – Mobergella radiolata: а – снаружи, б – изнутри, в – сбоку; г–е – M. holsti: г – снаружи, д – изнутри, е – сбоку; ж–и – M. turgida: ж – снаружи, з – изнутри, и – сбоку; к–м – Discinella micans: к – снаружи, л – изнутри, м – сбоку; н–п – Aktugaia triangula: н – снаружи, о – изнутри, п – сбоку; р–т – A. duodecifoliata: р – снаружи, с – изнутри, т – сбоку; у–х – Tateltella ranoculata: у – снаружи, ф – изнутри, х – сбоку; ц–ш – Hippoklosma mongolica: ц – снаружи, ч – изнутри, ш – сбоку.
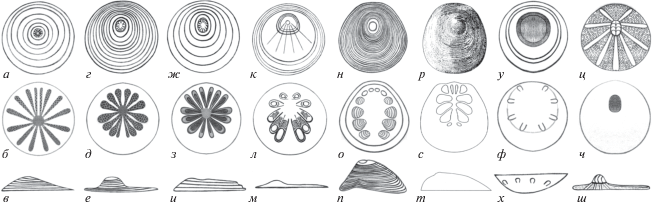
МОРФОЛОГИЯ МОБЕРГЕЛЛИД
Вслед за Миссаржевским (1989), мы рассматриваем мобергеллид в ранге семейства Mobergellidae, включающего следующие роды – Mobergella, Discinella, Aktugaia Missarzhevsky, 1976, Hippoklosma Conway Morris et Chapman, 1997 и Tateltella Streng et Skovsted, 2006. Род Brastadella является младшим объективным синонимом рода Discinella, так как его типовой вид B. brastadi (Poulsen, 1932) оказался (Skovsted, 2003) синонимом типового вида рода Discinella – D. micans (Billings, 1871). Род Thorslundella Nyers, 1984 является младшим субъективным синонимом рода Aktugaia, так как его типовой вид T. duodecifoliata Nyers, 1984 помещен в состав рода Aktugaia (Skovsted, 2003). Кратко остановимся на характеристиках валидных родов мобергеллид.
Род Mobergella Hedström, 1923 (рис. 1, а–и) характеризуется дисковидными (уплощенными, вогнутыми или выпуклыми) склеритами миллиметровой размерности. Наружная поверхность склерита несет отчетливые концентрические линии нарастания. Макушечное поле и сама макушка склерита сдвинуты к краю склерита, который назван апикальным. Внутренняя поверхность склерита несет 10–16 радиально расходящихся и билатерально расположенных м.о. с пористой поверхностью (табл. II ), сгруппированных попарно (Bengtson, 1968; Миссаржевский, 1989; Демиденко, 2016).
Род Discinella Hall, 1872 (рис. 1, к–м) представлен толстостенными уплощенными, либо сильно выпуклыми, либо сильно вогнутыми склеритами с радиальными ребрами на наружной поверхности; на внутренней поверхности присутствуют 8‒10 непористых м.о. в виде углублений грушевидной формы с дуговидными линиями роста (Миссаржевский, 1989; Skovsted, 2003; Демиденко, 2016).
Род Aktugaia Missarzhevsky, 1976 (рис. 1, н–т) представлен массивными сильно выпуклыми колпачковидными склеритами, на внутренней поверхности которых расположены 12 непористых м.о. в виде неглубоких каплевидных углублений с дуговидными линиями роста (Миссаржевский, 1976; Skovsted, 2003).
Род Tateltella Streng et Skovsted, 2006 (рис. 1, у–х) включает в себя вогнутые склериты с нечеткой радиальной ребристостью на наружной поверхности и 4–8 выпуклыми м.о. на внутренней поверхности, дистально приподнятыми, гладкими или с нечеткими дуговидными линиями роста (Streng, Skovsted, 2006).
Род Hippoklosma Conway Morris et Chapman, 1997 (рис. 1, ц–ш) представлен тонкими вогнуто-выпуклыми склеритами, на наружной поверхности которых отчетливо видны пять пар радиально расходящихся зон, несущих многочисленные фиброплакоды – дисковидные углубления в стенке склерита, заполненные тонкими плотно упакованными фибрами и закрытые гладкой крышечкой (Conway Morris, Chapman, 1997). Подобные структуры могут проявляться и на внутренней поверхности склеритов (Skovsted, 2003).
Ювенильная часть склеритов Mobergella по морфологическим признакам напоминает (Демиденко, 2016; Skovsted, Topper, 2018) личиночные раковины некоторых представителей лофотрохозой – брахиопод и хиолитов (крышечки), и, возможно, представляет собой раннюю минерализованную стадию роста склерита, по времени соответствовавшую метаморфозу личинки и ее переходу от планктонного к бентосному образу жизни (Skovsted, Topper, 2018). Изучение вида Discinella micans (Billings, 1871) из нижнего кембрия Сев. Гренландии показало наличие шапочковидной личиночной стадии с внутренними м.о. (Skovsted, Topper, 2018).
МОРФОГЕНЕЗ МОБЕРГЕЛЛИД
Проанализировав время существования родов и видов мобергеллид, можно наметить ряд эволюционных преобразований в морфологии их склеритов. Самый древний представитель группы – вид Mobergella radiolata Bengtson, 1968 – имеет дисковидные (уплощенные, вогнутые, выпуклые или вогнуто-выпуклые) тонкостенные склериты диаметром от 0.4 до 1.7 мм с 10–14 пористыми узкими м.о. (Bengtson, 1968; Демиденко, 2016). Он появляется на Сибирской платформе на рубеже томмотского–атдабанского веков и продолжает существовать до середины атдабана в Сибирском регионе и в Европе (Пархаев и др., 2020). В первой половине атдабана происходит расселение вида на территории Казахстана и Европы (Швеция, Норвегия, Дания, Польша, Великобритания).
Вместе с M. radiolata в раннем кембрии Европы встречаются еще два вида мобергелл. Один из них – это появившийся в середине атдабана M. holsti (Moberg, 1892), известный из Швеции (Moberg, 1892), Норвегии (Åhman, Martinsson, 1965; Brasier, 1989), Польши (Lendzion, 1983). Также предполагаются находки этого вида в ботомских? толщах Центрального Казахстана (Конева, 1983). M. holsti имеет более крупные склериты (диаметр до 4.4 мм) с сильно выпуклой макушкой, мускульное поле сильно сдвинуто к апикальному краю, отчетливые м.о. (как правило 14–16 шт.) пористые, расширяются дистально. Второй вид – M. turgida Bengtson, 1968 – известен из атдабана Швеции (Bengtson, 1968), Сев. Польши (Lendzion, 1983) и Англии (Bengtson, 1977). Он отличается от M. radiolata толстостенными склеритами от 0.96 до 2.1 мм в диаметре и большим количеством м.о. (14–16 шт.). Мускульные отпечатки вытянутые, сильно утолщенные в дистальных частях, непористые.
Далее развитие мобергеллид идет по пути уменьшения числа м.о. и одновременного увеличения их относительного размера и площади. В позднем атдабане Казахстана появляется новый вид Aktugaia triangula Missarzhevsky, 1976. Это форма с сильно выпуклой раковиной, 12 м. о. в виде гладких неглубоких углублений с аркообразными линиями роста (Skovsted, 2003). Несомненно близкий вид A. duodecifoliata (Nyers, 1984), немного отличающийся от типового вида рода Aktugaia конфигурацией мускульных отпечатков, известен из ранней ботомы Центральной Швеции (Nyers, 1984, 1987; Skovsted, 2003).
Во второй половине ботомы востока Лаврентии (cеверо-восток США, Гренландия, Квебек, Лабрадор и запад Ньюфаундленда Канады) появляется вид Discinella micans (Billings, 1871). Это различной степени выпуклости склериты (от сильно выпуклой через уплощенную до сильно вогнутой) с 8–10 м. о. лепестковидной формы в виде гладких неглубоких углублений с дуговидными линиями роста (Skovsted, 2003).
В ранней ботоме Монголии появляется вид Hippoklosma mongolica (Missarzhevsky, 1989). Он представлен тонкими вогнуто-выпуклыми склеритами, наружная поверхность которых несет пять пар радиальных зон со сложно устроенной микроструктурой. Предполагается (Conway Morris, Chapman, 1997), что эти зоны гомологичны м.о. мобергелл.
На рубеже раннего–среднего кембрия (поздний тойон–ранняя амга) в Западной Гондване (Марокко) появляется последний вид мобергеллид – Tateltella ranoculata Streng et Skovsted, 2006. Склериты вогнутые, с нечеткой радиальной ребристостью, несут 4–8 выпуклых м.о., их поверхность гладкая или с нечеткими дуговидными линиями роста.
Таким образом, можно обозначить три основных морфогенетических тренда в эволюции мобергеллид на протяжении раннего – начала среднего кембрия. Во-первых, отчетливо наблюдается редукция числа м. о. от 12–16 у видов рода Mobergella, встречающихся в первой половине раннего кембрия (поздний томмот – ранний атдабан) до 12 у Aktugaia (поздний атдабан – ранняя ботома), 10 у Hippoklosma mongolica (ранняя ботома), 8–10 у Discinella micans (поздняя ботома) и 4–8 у Tateltella ranoculata (поздний тойон – ранняя амга). Вместе с уменьшением числа м. о. растет их относительная ширина (рис. 1): у M. radiolata – м. о. в виде узких полосок, у M. holsti и M. turgida м.о. заметно расширяются к периферии склерита, у Aktugaia м.о. субокруглые, у Hippoklosma – очень широкие, у Discinella м. о. лепестковидные, сильно расширенные дистально, у Tateltella м. о. заметно расширяются к периферии склерита и располагаются на выпуклых валиках. Также следует отметить изменение в характере микроскульптуры поверхности м. о.: у M. radiolata и M. holsti она ноздреватая; у M. turgida, Aktugaia triangula, Discinella micans и Tateltella ranoculata – гладкая, иногда несущая лишь дуговидные следы роста; y Hippoklosma mongolica состоящая из сложно устроенных округлых структур – фиброплакод. Пока преждевременно говорить о причинах подобных морфогенетических изменений у мобергеллид, возможно, это следует связывать с адаптациями к различным условиям обитания конкретных видов.
ПАЛЕОЭКОЛОГИЯ МОБЕРГЕЛЛИД
Для реконструкции образа жизни мобергеллид важно проанализировать фациальную приуроченность находок (табл. 1). Наиболее репрезентативную информацию мы имеем по виду Mobergella radiolata, который встречается в десятках разрезов различных структурно-фациальных регионов обширного палеобассейна Сибирской платформы, а также широко распространен на территории Северной и Восточной Европы и, возможно, Казахстана (Пархаев и др., 2020, рис. 1, 4, 6 ).
Таблица 1.
Литологическая и фациальная приуроченность видов мобергеллид
| Вид | Стратон | Литология | Обстановки осадконакопления | Источник |
|---|---|---|---|---|
| Mobergella radiolata | Средняя часть пестроцветной свиты (Сибирская платформа) | Известняки различно-глинистые (до мергелей), вишнево-красные, розовые, желтые, сиреневые, зеленовато-серые | Зона перехода от внешнего карбонатного шельфа к склону открытого бассейна | Кембрий Сибири, 1992; Сухов и др., 2016 |
| Средняя часть краснопорожской свиты (Сибирская платформа) | Пестроцветные, неравномерно глинистые и детритистые известняки | Зона перехода от внешнего карбонатного шельфа к склону открытого бассейна | Кембрий Сибири, 1992; Сухов и др., 2016 |
|
| Тюсэрская свита, средняя подсвита (Сибирская платформа) | Пестрые алевро-аргиллиты, известковистые алевролиты и сильноглинистые известняки | Склон мелководного бассейна | Кембрий Сибири, 1992; Сухов и др., 2016 |
|
| Средняя часть эмяксинской свиты (Сибирская платформа) | Известняки глинистые, красноцветные | Внешний карбонатный шельф | Кембрий Сибири, 1992; Сухов и др., 2016 |
|
| Сыгдахская свита (Сибирская платформа) | Пестроцветные доломиты, глинстые до мергелистых, доломиты и известняковые песчаники | Зона перехода от внешнего карбонатного шельфа к склону открытого бассейна | Сухов и др., 2016 | |
| Низы формации Норреторп (Швеция) | Алевролиты с прослоями песчанников и глауконитом | Мелководные обстановки | Bengtson, 1968; Bergström, 1981; Nielsen, Schovsbo, 2007, 2011 | |
| Низы формации Филе-Хайдар, песчаники Mickwitzia (Швеция) | Тонкослоистые песчаники и алевролиты с глинистыми прослоями (темпеститы) | Мелководные обстановки на удалении от береговой линии обширного эпиконтинентального бассейна | Jensen, 1997 | |
| Формация Филе-Хайдар, конгломераты Вененяс (Швеция) | Среднезернистый песчаник, с хорошо окатанными зернами кварца, карбонатным цементом и кварцитовой галькой | Мелководные обстановки с активной гидродинамикой и переотложением осадка | Ahman, Martinsson, 1965; Bengston, 1968; Brasier, 1989; Topper, Skovsted, 2017 |
|
| Люкатиские слои (Эстония) | Переслаивание зеленовато-серых песачнистых аргиллитов и светло-серых алевролитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Менс, Пиррус, 1977; Mens et al., 1990; Nielsen, Schovsbo, 2011 | |
| Низы тискреских слоев (Эстония) | Грубозернистые алевролиты с линзами разнозернистых песчанников и конгломератов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Менс, Пиррус, 1977; Mens et al., 1990; Nielsen, Schovsbo, 2011 | |
| Завишинские слои (Польша) | Серые и серо-зеленые мелко- и среднезернистые кварц-глауконитовые песчаники с редкими прослоями и линзами песчанистых аргиллитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974; Dzik, Lendzion, 1988; Mens et al., 1990; Nielsen, Schovsbo, 2011 | |
| Мобергелловые слои (Польша) | Мелко- и среднезернистые песчанники с прослоями аргиллитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974; Dzik, Lendzion, 1988; Nielsen, Schovsbo, 2011 | |
| Глебовские слои (Московская синеклиза) | Светло-зеленые и зелено-серые аргиллитоподобные глины с редкими прослоями и линзами кварцевых алевролитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Розанов, 1973; Кирсанов, 1974; Rozanov, Zhuravlev, 1992; Nielsen, Schovsbo, 2011 | |
| Мобергелловые слои, керимбайская свита (Казахстан) | Алевролиты и известковистые песчаники | Склоновые терригенные отложения в зоне распространения островных вулканических дуг | Конева, 1983; Дергунов и др., 1986; Гридина, 1990; Ившин и др., 1993; Коробкин, Смирнов, 2006; Дегтярев, 2012 | |
| Mobergella turgida | Формация Филе-Хайдар, конгломераты Вененяс (Швеция) | Среднезернистый песчаник, с хорошо окатанными зернами кварца, карбонатным цементом и кварцитовой галькой | Мелководные обстановки с активной гидродинамикой и переотложением осадка | Ahman, Martinsson, 1965; Bengston, 1968; Brasier, 1989; Topper, Skovsted, 2017 |
| Мобергелловые слои (Польша) | мелко- и среднезернистые песчанники с прослоями аргиллитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974; Dzik, Lendzion, 1988; Nielsen, Schovsbo, 2011 | |
| Низы песчанников Комли (Англия) | Карбонатные песчаники с глауконитом и биокластами | Мелководный шельф | Rushton, 1974, 2000; Bengtson, 1977; Brasier, 1989; Brasier et al., 1992; Rushton, Molyneux, 2011 | |
| Mobergella holsti | Формация Филе-Хайдар, конгломераты Вененяс (Швеция) | Среднезернистый песчаник, с хорошо окатанными зернами кварца, карбонатным цементом и кварцитовой галькой | Мелководные обстановки с активной гидродинамикой и переотложением осадка | Ahman, Martinsson, 1965; Bengston, 1968; Brasier, 1989; Topper, Skovsted, 2017 |
| Мобергелловые слои (Польша) | Мелко- и среднезернистые песчанники с прослоями аргиллитов | Мелководные обстановки на фоне крупной доминопольской трансгрессии | Lendzion, 1972, 1975, 1983; Nielsen, Schovsbo, 2011 |
|
| Мобергелловые слои, керимбайская свита (Казахстан) | Алевролиты и известковистые песчаники | Склоновые терригенные отложения в зоне распространения островных вулканических дуг | Конева, 1983; Дергунов и др., 1986; Гридина, 1990; Ившин и др., 1993; Коробкин, Смирнов, 2006; Дегтярев, 2012 | |
| Discinella micans | Средняя сланцева пачка, формация Форти (Лабрадор, Канада) | Органогенно-обломочные известняки, межбиогермовые фации | Шельфовые мелководные обстановки с активной гидродинамикой | Spencer, 1980; Skovsted, Topper, 2018 |
| Пачка Дэвилз-Коув, формация Форти (Зап. Ньюфаундленд, Канада) | Розовые, зеленовато-серые, известняки с прослоями пурпурных песчанников | Мелководные трансгрессивные обстановки внешнего шельфа с археоциатовыми патч-рифами | Skovsted et al., 2017; Skovsted, Topper, 2018 |
|
| Верхняя часть формации Бастион (СВ Гренландия) | Темноцветные карбонатные сланцы с прослоями известняков | Мелководные обстановки внешнего шельфа ниже уровня волнового воздействия | Skovsted, 2003, 2004, 2006 | |
| Формация Элла-Айленд (СВ Гренладния) | Известняки, местами с оолитами | Мелководные обстановки внешнего шельфа с активной гидродинамикой | Skovsted, 2003, 2004, 2006 | |
| формация Ансе-Моранда (Квебек, Канада) | Серые сланцы и известняки | Турбидиты на верхней части континентального склона | Landing et al., 2002 | |
| формация Браунз-Понд (шт. Нью-Йорк, США) | Темно серые алевролиты с прослоями органогенно-облмочных известняков | Проксимальные фации турбидитовых потоков на верхней части континентального склона | Landing, Bartowski, 1996 | |
| Aktugaia triangula |
Гересская пачка, шабактинская свита (Казахстан) | Слоистые органогенно-обломочные доломитизированные известняки с прослоями ракушняков | Шельфовые мелководные обстановки с активной гидродинамикой | Миссаржевский, 1976; Миссаржевский, Мамбетов, 1981; Skovsted, 2003 |
| Aktugaia duodecifoliata | Пачка Ломвикен, формация Вассбо (Швеция) | Базальные конгломераты | Мелководные осадки | Nyers, 1984, 1987; Nielsen, Schovsbo, 2011 |
| Tateltella ranoculata |
Пачка Breche a Micmacca, формация Джбель-Ваврмаст (Марокко) | Органогенно-обломочные известняки слоистые и нодулярные c прослоями песчанников и ракушняков | Мелководные осадки со следами перемыва и осушения, формировавшиеся в ходе трансгрессивно-регрессивных событий | Streng, Skovsted, 2006; Geyer, Landing, 2006 |
| Hippoklosma mongolica |
Хаирханская свита (Западная Монголия) | Песчаники и сланцы с линзами и прослоями пасчанистых с глауконитом известняков | Моласса – мелководно-морские и затем континентальные отложения, заполняющие краевой прогиб на завершающей стадии равития морского бассейна | Коробов, Миссаржевский, 1977; Воронин и др., 1982; Conway Morris, Chapman, 1997; Smith et al., 2016 |
В сибирских разрезах M. radiolata встречается в отложениях пестроцветной свиты (Лено-Алданский, Учуро-Майский р-ны), краснопорожской свиты (Игарский р-н), средней подсвиты тюсэрской свиты (хр. Хараулах), эмяксинской и сыгдахской свит (внутренние р-ны). Эти толщи представлены преимущественно известняками с различным содержанием терригенной примеси. Предполагается (Сухов и др., 2016), что данные осадки формировались в мелководных условиях внешнего карбонатного шельфа платформы (пестроцветная, краснопорожская, эмяксинская свиты), реже – в зоне перехода от шельфа к склону (сыгдахская свита) или в верхней части континентального склона (средняя подсвита тюсэрской свиты) (табл. 1). Можно сделать вывод, что представители вида на территории Сибирского палеобассейна были приурочены к карбонатным илам, накапливавшимся в различных мелководных обстановках открытого моря. Следует отметить отсутствие находок M. radiolata в лагунных частях бассейна запада платформы, а также в межбиогермных фациях как на юге (наши полевые наблюдения), так и на севере платформы (Репина и др., 1974; Пархаев и др., 2020). В связи с последним обстоятельством, данные Т. Топпера и К. Сковстеда, указывающие на распространение мобергелл в межбиогермных слоях нижнего кембрия Сибири (Topper, Skovsted, 2017, табл. 1), не вполне точны. Исключительно хорошая сохранность тончайших склеритов M. radiolata в изученных образцах с территории Сибирской платформы исключает сколь-нибудь значимый перенос или переотложение ископаемых, поэтому их находки можно считать автохтонными.
На территории Балтийского палеобассейна находки M. radiolata приурочены к разнообразным терригенным фациям – от глин до конгломератов (табл. 1). Накопление большей части этих формаций происходило в крайне мелководных прибрежно-морских условиях, часто в зоне активной гидродинамики: отмечаются перемывы и переотложение материала. Можно предположить, что представители вида на территории Балтии обитали на песчано-глинистых грунтах мелководья. Находки в крупно-обломочных литологических разностях (конгломераты, гравелиты и крупно-зернистые песчаники) являются, по-видимому, аллохтонными, что подтверждается значительно худшей сохранностью материала в этих толщах (Bengtson, 1968).
Таким образом, вид M. radiolata следует рассматривать как обитателя разнообразных по составу мягких грунтов мелководных участков морских бассейнов как в низких широтах (Сибирская платформа, Палеонезия), так и в более высоких широтах (Балтия). Учитывая крайне широкое географическое распространение и пестроту литологического состава вмещающих отложений, данный вид мобергеллид, вероятно, являлся наиболее эврибионтным по сравнению с остальными представителями группы.
Другие виды мобергелл – M. turgida и M. holsti – встречаются совместно с M. radiolata в терригенных отложениях Балтии. M. turgida также отмечен в Авалонии (Англия), а M. holsti – на территории Палеонезии (Казахстанский террейн) (табл. 1). Везде данные виды приурочены к мелководным терригенным формациям (песчаники, алевролиты, аргиллиты), только лишь для находок M. holsti в Казахстане можно предположить чуть более глубоководные условия обитания (или захоронения?), так как накопление керимбайской свиты, в которой отмечены находки этого вида, связывается со склоновыми условиями в зоне распространения островных вулканических дуг.
Discinella micans широко распространен в разрезах востока Лаврентии (табл. 1) и приурочен, в основном, к карбонатным фациям (известняки, карбонатные сланцы). Условия осадконакопления этих формаций интерпретируются как мелководные, располагавшиеся на внешнем шельфе платформы (Лабрадор и Гренландия) или чуть более глубоководные, располагавшиеся в верхней части континентального склона в проксимальной зоне разгрузки турбидитовых потоков (Нью-Йорк, США; Квебек, Канада). Скорее всего, дисцинеллы в отложениях турбидитов аллохтонны. Следует особо отметить находки D. micans в межбиогермных органогенно-обломочных известняках формации Форти (Spencer, 1980; Skovsted et al., 2017; Topper, Skovsted, 2017).
Оставшиеся четыре вида мобергеллид – Aktugaia triangula, A. duodecifoliata, Tateltella ranoculata и Hippoklosma mongolica – имеют локальное распространение (табл. 1), и информации по их фациальной приуроченности у нас не много. Но можно утверждать, что три вида из четырех приурочены к карбонатным, преимущественно органогенно-обломочным толщам, формирование которых происходило в мелководных обстановках низких широт (кроме вида A. duodecifoliata, известного из терригенных толщ Центральной Швеции).
Подводя итог анализа фациальной приуроченности мобергеллид, можно сделать следующий вывод. Виды рода Mobergella, вероятно, были широко распространены на мягких грунтах верхней сублиторали, возможно, иногда проникали в нижние горизонты сублиторали и даже чуть ниже – в батиаль, на верхнюю часть континентального склона. Discinella также связаны с мелководными участками сублиторали, где могли селиться как на мягких грунтах, так и в водорослево-археоциатовых постройках типа патч-риф.
ОПЕРКУЛЯРНАЯ ПРИРОДА МОБЕРГЕЛЛИД
Гипотеза о том, что мобергеллиды – это крышечки организмов, обитающих в трубчатых “домиках”, обсуждается очень давно – со времен описания Э. Биллингсом склеритов Discinella micans (Billings, 1871) как крышечек хиолительминтов Hyоlithellus micans Billings, 1871 (Billings, 1871; Walcott, 1886, 1890; Knight, 1941; Lochman, 1956; Fisher, 1962). Однако до сих пор не найдено убедительных доказательств в пользу этого предположения, и не установлены подходящие по размеру “домики–трубки”.
C. Бенгтсон указывал на важные замечания Дж. Моберга (Moberg, 1892) о том, что в тех породах, где есть Mobergella, находки форм, близких к Hyolithellus micans, крайне редки и, наоборот, там, где обычны трубки хиолительминтов, Mobergella отсутствует (Bengtson, 1968, с. 347–348). Только в одном образце из местонахождения Стора-Рер Швеции, наряду со склеритами мобергелл, найдено как минимум три хиолителлоидных трубки (Moberg, 1892). Единственным местонахождением, где в значительном количестве совместно найдены склериты мобергелл и трубки хиолительминтов, является Аспелунд (Ӓspelund) в Норвегии (Åhman, Martinsson, 1965). С.П. Конева (1983) при изучении склеритов рода Mobergella из мобергелловых слоев нижнего кембрия Селетинского синклинория подчеркивает их многочисленность, указывая при этом на довольно редкие находки хиолительминтов Hyolithellus sp.
Тестируя оперкулярную гипотезу, Топпер и Сковстед провели анализ процентного соотношения участков, занимаемых м.о. у Mobergella holsti и Discinella micans, по отношению к остальной части склерита, и сравнили полученные данные с другими беспозвоночными (Topper, Skovsted, 2017). На долю м.о. мобергеллид приходится около 20%, это значение довольно высоко по сравнению с остальными группами и соизмеримо лишь с крышечками брюхоногих моллюсков сем. Neritidae. Ссылаясь на это, авторы считают оперкулярную гипотезу наиболее подходящей для склеритов мобергеллид, хотя отмечают, что сложно подобрать подходящие по размеру и форме сечения трубчатые окаменелости из встречающихся совместно таксонов. Так, несмотря на совстречаемость склеритов D. micans с фосфатными трубками Hyolithellus, хиолитами и различными известковыми трубками в Северо-Восточной Гренландии и на Лабрадоре (Skovsted, 2006; Skovsted, Peel, 2007, 2010; Skovsted, Topper, 2018), размер и морфология этих трубок не соответствуют размерам и морфологии D. micans (Skovsted, Topper, 2018).
Мы проанализировали совстречаемость склеритов мобергеллид со всеми известными видами хиолительминтов – ископаемыми, представленными фосфатными трубками с округлым сечением, стратиграфическое и географическое распространение которых приведено на рис. 2. Анализируя временнýю совстречаемость мобергеллид и хиолителлусов, легко заметить, что стратиграфический диапазон распространения мобергеллид существенно уже, чем у хиолительминтов. Как отмечалось выше, самые древние мобергеллы известны из базального атдабана, тогда как первые хиолительминты появляются на полтора века раньше – в середине немакит-далдынского времени, и в интервале немакит-далдын – поздний томмот известно уже более 10 видов рода Hyolithellus. Аналогичная, но обратная картина наблюдается и с исчезновением мобергеллид из геологической летописи – самые поздние их представители известны с рубежа раннего–среднего кембрия, тогда как несколько видов хиолительминтов продолжают существовать в среднем и даже позднем кембрии (рис. 2).
Рис. 2.
Географическое и стратиграфическое распространение видов мобергеллид (①–⑧) и видов рода Hyolithellus ($\boxed1$‒$\boxed{22}$); интервал распространения мобергеллид залит светло серым. Обозначения: ① – Mobergella radiolata Bengtson, 1968: нижний – средний атдабан, Сибирская платформа; нижний атдабан, Московская синеклиза, средний атдабан, Швеция, Норвегия, Польша, Эстония; ? нижний атдабан, Юж. Китай; ? ботома, Казахстан (Пархаев и др., 2020); ② – M. turgida Bengtson, 1968: средний атдабан, Швеция (Bengtson, 1968, 1977), Польша (Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974), Центр. Англия (Bengtson, 1977; Brasier, 1989; Brasier et al., 1992); ③ – M. holsti (Moberg, 1892): средний атдабан, Швеция (Bengtson, 1968), Польша (Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974); ? ботома, Казахстан (Гридина, 1990); ④ – Discinella micans (Billings, 1871): средняя – верхняя ботома, С.-В. США (Landing, Bartowski, 1996), Квебек, Лабрадор, Зап. Ньюфаундленд, Канада (Spencer, 1980; Landing et al., 2002; Skovsted, Topper, 2018), Вост. Гренландия (Skovsted, 2003, 2004, 2006); ⑤ – Aktugaia triangula Missarzhevsky, 1976: верхний атдабан, слои с Adyshevitheca, хр. Малый Каратау, Казахстан (Миссаржевсктй, Мамбетов, 1981); ⑥ – A. duodecifoliata Nyers, 1987: нижняя ботома, Швеция (Nyers, 1984, 1987; Nielsen, Schovsbo, 2011); ⑦ – Tateltella ranoculata Streng et Skovsted, 2006: тойон – амга, Марокко (Streng, Skovsted, 2006); ⑧ – Hippoklosma mongolica (Missarzhevsky, 1989): нижняя ботома, Зап. Монголия (Коробов, Миссаржевский, 1977; Воронин и др., 1982; Conway Morris, Chapman, 1997); $\boxed1$ – Hyolithellus tenuis Missarzhevsky, 1966: томмот, Магаданская обл.; томмот – тойон, Красноярский край (Миссаржевский, 1966; Хоментовский, Карлова, 1989); томмот, слои с Tianzhushania tetramera; ботома, зона Mongolitubulus squamifer, Зап. Монголия; ботома, С.‑З. Монголия (Воронин и др., 1982; Есакова, Жегалло, 1996); нижний – верхний кембрий, Казахстан (Есакова, Жегалло, 1996); мейшучунь – чунчуси, Юж. Китай (Есакова, Жегалло, 1996); ср. кембрий, серия Дамао, о. Хайнань, Китай (Jiang, Huang, 1986); средний атдабан, формация Лоуер Тэл, Гималаи, Индия (Brasier, Singh, 1987); $\boxed2$ – H. vladimirovae Missarzhevsky, 1966: томмот, слои с Tianzhushania tetramera и ботома, зона Mongolitubulus squamifer, Зап. Монголия (Есакова, Жегалло, 1996); томмот и низы атдабана, ср. течение р. Лены, Сибирская платформа (Миссаржевский, 1966); нижний томмот, зона Pseudorthotheca costata – верхний атдабан, слои с Adyshevitheca, Казахстан (Миссаржевский, Мамбетов, 1981); томмот, зона Protohertzina anabarica – ботома, низы зоны Bonnia–Olenellus, Юкон и Северо-Западные Территории, Канада (Есакова, 1987); мейшучунь, пров. Юньнань, Хубей, Китай (Qian, 1989); томмот, средняя доломитовая пачка, Сев. Иран (Hamdi et al., 1989); средний атдабан, формация Лоуер Тэл, Гималаи, Индия (Brasier, Singh, 1987); атдабан, пачка Хом Фам, Англия (Brasier, 1984); $\boxed3$ – H. insolitus Grigorieva, 1982: томмот, слои с Halkieria amphora и Tianzhushania tetramera, ботома?, Зап. Монголия, С.-З. Монголия (Воронин и др., 1982; Есакова, Жегалло, 1996); томмот, зона D. regularis и низы атдабана, Сибирская платформа (Ярусное расчленение.., 1983; Есакова, Жегалло, 1996); амга, Узбекистан (Есакова, Жегалло, 1996); амга и сакс, Казахстан (Конева и др., 1990); средний атдабан, формация Лоуер Тэл, Гималаи, Индия (Brasier, Singh, 1987; Есакова, Жегалло, 1996); чунчуси, зона Microcornus – Rhombocorniculum, пров. Хубей, Китай (Qian, 1989); амга, Туркестанский хр., Киргизия (Мешкова, 1985); $\boxed4$ – H. isiticus Missarzhevsky, 1969: томмот, зона N. sunnaginicus, Сибирская платформа (Розанов и др., 1969; Ярусное расчленение…, 1983); мейшучунь, формация Майдипин, пров. Сычуань, Китай (Qian, 1989); средний атдабан, формация Лоуер Тэл, Гималаи, Индия (Brasier, Singh, 1987); ботома, слои с Strenuella, Англия (Hinz, 1987); ботома, эрратические валуны, о-в Кинг-Джордж, Антарктида (Wrona, 1989); $\boxed5$ – H. micans Billings, 1871: ботома, слои с Elliptocephala asaphoides, США (Lochman, 1956); ботома, Зап. Монголия; чунчуси, пров. Сычуань, Китай (Yin et al., 1980); средний – верхний атдабан, зоны Mobergella и Holmia, Польша (Lendzion, 1972); ботома, слои с Serrodiscus, Германия (Есакова, Жегалло, 1996); атдабан, зоны “Obolella” groomi, Coleoloides typicalis и Callavia, ботома, зона Protolenus–Strenuellida, Центр. Англия (Cobbold, Pocock, 1934; Brasier, 1984); атдабан, зона Holmia kjerulfi, Ю.-З. Финляндия (Есакова, Жегалло, 1996), атдабан, зона Holmia kjerulfi – ботома, зона Ornamentaspis linnarssoni, о. Борнхольм, Дания (Poulsen, 1964); ботома, сланцы с Serrodiscus, Callodiscus и Olenellus, юг Шпицбергена (Major, Winsnes, 1955); атдабан, зона Fallotaspis, пров. Северо-Западные Территории, Канада (Есакова, 1987); атдабан, формация Веймауз, Массачусетс, США (Landing, 1988); атдабан и ботома, известняки Кулпара, Парара, Аякс, Юж. Австралия (Bengtson et al., 1990; Gravestock et al., 2001); $\boxed6$ – H. annulatus Meshkova, 1969: томмот, ср. течение р. Лены, Сибирская платформа (Мешкова, 1969); $\boxed7$ – H. grandis Missarzhevsky, 1969: томмот, зоны N. sunnaginicus – D. lenaicus, ср. течение р. Лены, Сибирская платформа (Розанов и др., 1969); томмот, зона D. regularis, хр. Малый Каратау, Казахстан (Миссаржевский, 1989); $\boxed8$ – H. filiformis Bengtson, 1990: атдабан и ботома, известняки Парара, Аякс, Юж. Австралия (Bengtson et al., 1990; Gravestock et al., 2001); $\boxed9$ – H. limbatus Yue in Xing et al., 1984: мейшучунь, формация Куаньчуаньпу, пров. Шаньси, Китай (Xing et al., 1984); $\boxed{10}$ – H. rectus Mambetov, 1981: нижний–средний томмот, зона Pseudorthotheca costata, чулактауская свита, каратауская пачка, хр. Малый Каратау, Казахстан (Миссаржевский, Мамбетов, 1981); $\boxed{11}$ – H. vitricus Mambetov, 1981: верхний томмот, зона Bercutia cristata, р. Коксу; верхний атдабан, слои с Adyshevitheca, хр. Малый Каратау, Казахстан (Миссаржевский, Мамбетов, 1981); $\boxed{12}$ – H.? infundibuliformis Meshkova, 1969: томмот, хр. Хараулах, Сибирская платформа (Мешкова, 1969); $\boxed{13}$ – H. sinuosus Cobbold, 1921: атдабан, зона “Obolella” groomi, гр. Шропшир, Англия (Cobbold, 1921); $\boxed{14}$ – H. acutus Meshkova, 1985: амга, ур. Араван, Алайский хр., Киргизия (Мешкова, 1985); $\boxed{15}$ – H. longulus Meshkova, 1985: амга, ур. Мадыген, Туркестанский хр., Киргизия (Мешкова, 1985); $\boxed{16}$ – H. multifarious Meshkova, 1985: амга, ур. Араван, Алайский хр., Киргизия (Мешкова, 1985); $\boxed{17}$ – H. singularis Meshkova, 1985: амга, р-н Сулюкты, Туркестанский хр., Киргизия (Мешкова, 1985): $\boxed{18}$ – H. aravanicus Meshkova, 1985: амга, ур. Араван, Алайский хр., Киргизия (Мешкова, 1985); $\boxed{19}$ – H. aequalis Meshkova, 1985: амга, ур. Араван, Алайский хр. и р-н Сулюкты, Туркестанский хр., Киргизия (Мешкова, 1985); $\boxed{20}$ – H. botomaensis Meshkova, 1985: амга, еланская свита, ср. течение р. Лены и р. Ботома, Сибирская платформа (Мешкова, 1985); $\boxed{21}$ – H. minutus (Qian et Jiang, 1980): мейшучунь, пров. Юньнань, Китай (Qian, Jiang, 1980); $\boxed{22}$ – H. tschushunensis Valkov, 1968: томмот, слои с Allatheca cana, Оленекское поднятие, Сибирская платформа (Вальков, 1968); ВЕП – Восточно-Европейская платформа.
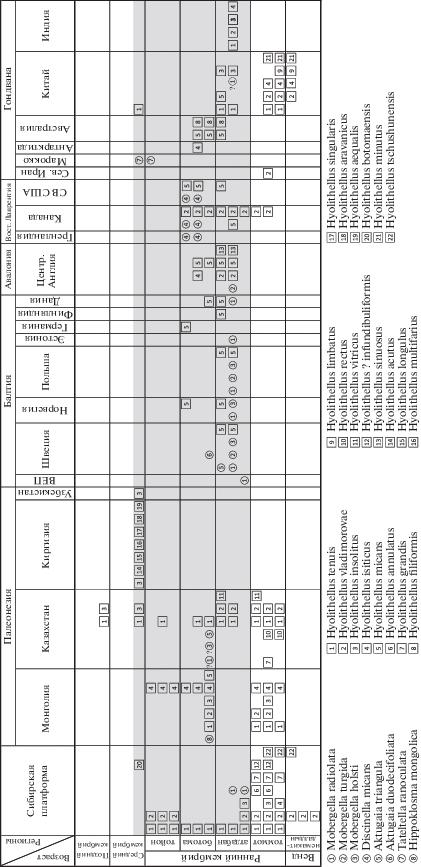
Рассматривая совстречаемость мобергеллид и хиолителлусов в географическом аспекте, также отмечается несопоставимость широты распространения данных групп. Во многих регионах, где обнаружены хиолителлусы, мобергеллиды не встречаются: Австралия, Антарктида, Индия, Иран, Германия, Финляндия, Дания, Киргизия.
Таким образом, проведенный нами анализ также не подтверждает связь между представителями сем. Mobergellidae и хиолительминтами, что ослабляет позиции сторонников оперкулярной природы мобергеллид и заставляет искать другую функциональную и систематическую гипотезу данным зоопроблематикам. Внешнее сходство со склеритами мобергеллид наблюдается у щитков современных сипункулид, морфологию, биологию и систематическое положение которых мы кратко рассмотрим ниже.
СОВРЕМЕННЫЕ СИПУНКУЛИДЫ
Сипункулиды – группа морских червеобразных бентосных лофотрохозой, характеризующихся билатеральной симметрией, отсутствием сегментации, наличием интроверта и венчика щупалец вокруг терминально расположенного рта (рис. 3, 4). Размер тела – от 1–3 мм до 500–700 мм, большая часть – 15–50 мм длиной. Большинство современных сипункулид населяет мягкие грунты на различных глубинах (от литорали до абиссали), где зарываются в осадок, либо обитают под камнями, в расщелинах скал и ризоидах водорослей, заселяют пустые раковины моллюсков, домики седентарных полихет; некоторые виды сверлят кораллы (Pancucci-Papadopoulou et al., 1999). По типу питания сипункулиды – микрофаги, потребляющие либо детрит, заглатывая осадок, либо сестон, улавливая его из толщи воды щупальцами; также в качестве пищи выступают мейобентосные животные и диатомовые (Вестхайде, Ригер, 2008).
Рис. 3.
План внутреннего строения сипункулид на примере рода Aspidosiphon Diesing, 1851 (по: Rice, 1993, рис. 15 ). Обозначения: a – анус, as – анальный щиток, cs – каудальный щиток, e – пищевод, i – спирально-завитый кишечник, in – основание интроверта, n– нефридий, r – прямая кишка, rm – мускул-ретрактор интроверта, sm – веретеновидный мускул, t – туловище.
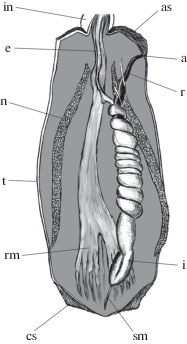
Рис. 4.
Современные сипункулиды рода Aspidosiphon Diesing, 1851 (сем. Aspidosiphonidae Quatrefages, 1865) с каудальными и анальными щитками: а – Aspidosiphon laevis Quatrefages, 1865; современный, Красное море, Египет (© Arthur Anker, печатается с разрешения автора фото); б, в – Aspidosiphon sp.; современный, Тихий океан, Муреа, Французская Полинезия: б – (© 2010 Moorea Biocode, печатается с разрешения Biocode MINV Team); в – (© 2012 Moorea Biocode, печатается с разрешения Biocode MINV Team). Обозначения: as – анальный щиток, cs – каудальный щиток, in – интроверт.
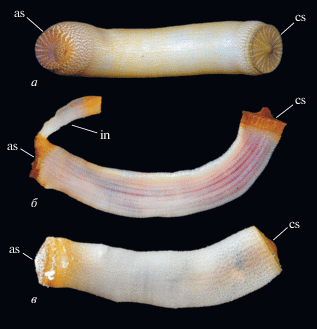
У современных представителей группы покровы тела обычно не минерализованы, но эластичная покровная кутикула часто несет различной формы протеиновые (но не хитиновые) крючья, шипики и папиллы (Voss-Foucart et al., 1977), используемые для фиксации в субстрате. Некоторые формы (представители сем. Aspidosiphonidae) имеют особые эпидермальные утолщения на туловище – каудальный (хвостовой) и анальный (передний) щитки (рис. 4). Округлый в плане каудальный щиток, уплощенный или конический, расположен на заднем конце туловища. Анальный щиток обычно колпачковидной формы, располагается дорсально в верхней части туловища, вблизи его границы с интровертом. У видов рода Aspidosiphon анальный щиток роговой, у Lithacrosiphon – минерализован карбонатом кальция. Анальный щиток Cloeosiphon состоит из отдельных пластинок, окружающих основание интроверта (Cutler, 1994; Schulze et al., 2005). Функция щитков различна: анальный щиток блокирует вход в норку животного после того, как интроверт втягивается в туловище; каудальный щиток используется для заякоривания в субстрате и/или участвует в процессе засверливания в субстрат (Rice, 1969, 1970). При этом радиальные ребра, гранулы или острые шипы на щитке действуют как фреза. Для некоторых видов рода Aspidosiphon отмечено (Rice, 1993; Pancucci-Papadopoulou et al., 1999), что парные мускулы-ретракторы интроверта крепятся вблизи каудального щитка (рис. 3), поэтому можно предположить, что каудальный щиток выполняет также и опорную функцию для мощной ретракторной мускулатуры. Морфология щитков имеет важное систематическое значение (Мурина, 1977).
Развитие у сипункулид прямое, либо со стадией планктонной личинки – лецитотрофной трохофоры, затем – лецитотрофной или планктотрофной пелагосферы, которая трансформируется в ювенильную особь (Вестхайде, Ригер, 2008). Личиночная стадия довольно продолжительная, что позволяет расселяться сипункулидам на значительные расстояния, как результат – многие современные виды – космополиты.
Систематическое положение сипункулид дискуссионно. В XIX в. их сближали с голотуриями, затем они были обособлены в “промежуточную” группу между первичноротыми “червями” и вторичноротыми иглокожими – Gephyrea. В конце XX в. стало очевидно, что сипункулиды – представители клады Lophotrochozoa и близкие родственники аннелид и, возможно, моллюсков (Schulze et al., 2005). С аннелидами их сближает схожая по микроструктуре кутикула и наличие трохофорной личинки, с моллюсками – наличие трохофорной личинки и характер дробления яйца с формированием так называемого “моллюскового креста” из особых микромеров в апикальной части яйца (Scheltema, 1993). Однако отсутствие сегментации и щетинок, характерных для кольчецов, мантии и раковины в каком-либо выражении, заставляют рассматривать сипункулид в качестве отдельного типа Sipuncula Rafinesque, 1814. В его пределах выделяют 2 класса, 4 отряда, 6 семейств и 17 родов (Schulze et al., 2005). Число видов современных сипункулид оценивается от 147 (Schulze et al., 2005) до 162 (Cutler, 1994). Отметим, что по последним данным молекулярной (анализ рибосомальных протеинов) и генетической (анализ митохондриальной ДНК) филогенетики (Shen et al., 2009; Struck et al., 2011), сипункулиды являются аннелидами, при кладистическом анализе признаков попадая внутрь различных клад кольчецов.
ИСКОПАЕМЫЕ СИПУНКУЛИДЫ
Из-за долгого отсутствия надежных ископаемых палеонтологических находок сипункулид интерпретация их происхождения и эволюции была затруднительна (Мурина, 1977). На рубеже XX–XXI вв. в ходе активного изучения раннекембрийских лагерштеттов Китая были описаны многочисленные новые таксоны, представленные мягкотелыми или слабоминерализованными ископаемыми. В знаменитых нижнекембрийских сланцах Маотяньшань (лагерштетт Ченьцзянь, уезд Аньнин, пров. Юньнань) были обнаружены червеобразные организмы, интерпретированные как ископаемые сипункулиды (Huang et al., 2004). Находки приурочены к толще сланцев, сформированной в ходе схода микротурбидита из близлежащего мелководья. О близости описанных форм к сипункулидам свидетельствует как внешняя морфология обнаруженных отпечатков (колбасообразное тело с тонким удлиненным интровертом и более широким туловищем, периоральная коронка щупалец, крючки, сосочки и складки на поверхности тела), так и отдельные элементы внутреннего строения (U-образный кишечник, положение анального отверстия вблизи зоны соединения интроверта и туловища). Описанные ископаемые таксоны Archaeogolfingia caudata Huang, Chen, Vannier et Salinas, 2004 и Cambrosipunculus tentaculatus Huang, Chen, Vannier et Salinas, 2004 имеют поразительное сходство с современными представителями Golfingiidae. Авторы статьи делают вывод, что основные морфологические черты сипункулид сформировались уже в раннем кембрии, и с тех пор группа находится в эволюционном стазисе. U-образный кишечник описанных форм свидетельствует об инфаунном образе жизни кембрийских сипункулид.
Другая кембрийская находка сипункулид происходит из среднекембрийской формации Стэфен, Британская Колумбия, Канада (Caron et al., 2010), где в местонахождении бёрджеского типа (Burgess Shale-Type, BST) обнаружены разнообразные организмы с неминерализованными покровами, в том числе не получившая еще названия сипункулида – Sipunculan A worm (Caron et al., 2010, рис. 3H, DR4-G, DR4-H), представленная пятью экземплярами различной сохранности. Авторы воздержались от описания нового таксона, отметив лишь, что морфология ископаемого в целом близка к материалу из нижнего кембрия Китая.
В более молодых палеозойских отложениях известны еще две находки ископаемых, интерпретируемые как сипункулиды – Lecthaylus gregarius Weller, 1925 из силура Иллинойса, США (Weller, 1925; Roy, Croneis, 1931; Fisher, 1962) и L. briggsi Muir et Botting, 2007 из нижнего карбона окрестностей Эдинбурга, Шотландия (Muir, Botting, 2007). В мезозое отмечены три находки: Valvasoria camiolica Kolar-Jurkovsek et Jurkovsek, 1997 из триаса Словении (Kolar-Jurkovsek, Jurkovsek, 1997), Epitrachys rugosus Ehlers, 1869 из известных позднеюрских литографских сланцев Зольнгофен Германии (Ehlers, 1869) и Rondeletia scutata Alessandrello, Bracchi et Riou, 2004 из среднеюрского лагерштетта Ла-Вульт-сюр-Рон Франции (Alessandrello et al., 2004). Отличительными чертами всех этих находок являются червеобразная форма тела с более толстым (туловище) и более тонким (интроверт) отделами и характерная продольная складчатость переднего отдела, которая формируется при сжатии и последующей фоссилизации мощной мускулатуры интроверта.
Как справедливо отмечают Л. Мюир и Дж. Боттинг (Muir, Botting, 2007), скорее всего палеонтологическая история сипункулид сильно “недооценена”, так как сипункулиды, как и многие “черви”, не обладают минерализованными покровами, но в добавок, именно у этой группы “червей” присутствует наименьшее число внешних диагностических признаков, по которым можно было бы определять ископаемые находки. Поэтому не исключено, что значительная часть ископаемых сипункулид пропущена исследователями.
ЭВОЛЮЦИЯ СИПУНКУЛИД
Вопросы эволюции группы поднимались еще с конца XIX–начала XX вв. (Selenka et al., 1883). Важно понять, какие признаки являются исходными (архаичными). По данным многих исследователей, наиболее древние сипункулиды могли иметь сплошной слой мускулатуры, несколько пар ретракторов интроверта, простой венчик щупалец, в один ряд окружающих центральное ротовое отверстие. Потом эволюция сипункулид могла идти по двум различным направлениям, именуемым катаморфозом (редукция ряда органов и упрощение организации) и алломорфозом (усовершенствование и усложнение организации) (Мурина, 1977; Догель, 1981). Например, упрощение организации проявляется в редукции щупалец с уменьшением числа лопастей, или их полной редукции; в редукции ретракторов и нефридиев путем уменьшения их количества. Усложнение организации проявляется в развитии сосудистой системы как сложного щупальцевого аппарата с дихотомическим ветвлением (щупальцевый венчик), так и сложного сократительного сосуда с длинными развилками или развилками, закрученными в спираль. Разветвленный щупальцевый венчик частично берет на себя функцию дыхания. Еще один алломорфоз в развитии сипункулид – усиление, а именно, утолщение мускулатуры и ее разделение на продольные ленты. Приобретение щитков на одном или обоих концах туловища некоторых сипункулид – еще один крупный алломорфоз в их развитии. Твердые протеиновые или известковые щитки служат для разрушения субстрата при бурении мертвых коралловых рифов, рыхлых и твердых известковых пород, тем самым способствуя усилению активности сипункулид и коренному изменению их образа жизни; главным образом, это относится к представителям класса Phascolosomatidea.
КЕМБРИЙСКИЕ МОБЕРГЕЛЛИДЫ – ДРЕВНЕЙШИЕ СИПУНКУЛИДЫ?
Неубедительность оперкулярной гипотезы для мобергеллид (см. выше) заставляет нас искать другие варианты возможного расположения склеритов этой группы на теле организмов. Наиболее близкими к склеритам мобергеллид морфологически оказались щитки на туловище современных сипункулид. Действительно, уплощенный каудальный и выпуклый анальный щитки рода Aspidosiphon напоминают склериты данной группы (рис. 4). Выше отмечалось, что у сипункулид каудальные щитки могут выполнять опорную функцию для мощной ретракторной мускулатуры, парные тяжи которой крепятся как раз в хвостовой части туловища. Тогда парные мускульные отпечатки на внутренней поверхности склеритов мобергеллид можно интерпретировать как место крепления мускулатуры данного организма. У современных сипункулид основных ретракторов одна-две пары, но нельзя исключать, что это результат олигомеризации мускулатуры – обычного явления в эволюции различных групп беспозвоночных, например, моллюсков. Важно отметить, что число м. о. на склеритах мобергеллид оставалось неизменным на протяжении его роста (т.е. неизвестны ювенильные склериты с меньшим числом м. о., чем на взрослых экземплярах того же вида), что соответствует модели прикрепления мускулов-ретракторов, число которых также было постоянно в ходе всего онтогенеза организма. Число м. о. лишь варьирует у разных таксонов этих зоопроблематик.
Отсутствие развитой ретракторной мускулатуры в районе анального щитка у современных сипункулид заставляет рассматривать все склериты мобергеллид с развитыми м.о. в качестве именно каудальных щитков, несмотря на тот факт, что некоторые склериты (напр., Aktugaia) довольно выпуклые и внешне близки к анальным щиткам сипункулид.
Если мы сопоставляем склериты мобергеллид с каудальными щитками сипункулид, то функция склеритов может быть реконструирована соответственно: (1) крепление мускулатуры ретракторов интроверта; (2) приспособление для заякоривания в рыхлом сусбстрате; (3) приспособление для сверления в уплотненном сусбстрате.
Таким образом, по сравнению с оперкулярной гипотезой, склерит занимает обратное положение – не сверху тела, а снизу. Отсутствие находок следов каких-либо эпибионтов на склеритах мобергеллид косвенно подтверждает идею о положении склерита внутри осадка. Предполагаемые следы сверления, обнаруженные на некоторых склеритах M. holsti (Bengtson, 1968; Conway Morris, Bengtson, 1994), не могут свидетельствовать в пользу какого-либо образа жизни жертвы, так как нам ничего не известно о самом хищнике, который мог оставлять следы сверления на склеритах – был ли он инфаунным и нападал на жертву внутри осадка, либо он был эпифаунным и охотился в придонном слое воды или на поверхности осадка.
Образ жизни, экология и характер географического распространения современных сипункулид не противоречат данным, которые мы реконструируем для кембрийских мобергеллид. Сипункулиды – обитатели мягких грунтов морского мелководья, но есть и более глубоководные представители. То же самое мы предполагаем и для кембрийских мобергеллид на основе анализа их фациальной приуроченности (см. выше). Некоторые современные сипункулиды – обитатели коралловых рифов, где они засверливаются в твердый субстрат. Сходный биотоп мы предполагаем для Discinella micans, находки которого отмечены в фациях, прилегающих к водорослево-археоциатовым постройкам. В связи с длительной личиночной стадией, географическое распространение некоторых современных сипункулид практически космополитное, то же можно сказать по крайней мере для одного кембрийского вида – Mobergella radiolata, который встречается как в низких широтах Сибирского палеобассейна, так и в умеренно высоких широтах Балтии и Авалонии. Такое сходство в экологии кембрийских и современных форм лишь укрепляет аргументы в пользу эволюционного стазиса, в котором находится группа последние 520–530 млн лет (Huang et al., 2004).
Основными аргументами против сипункулидной природы мобергеллид могут быть следующие наблюдения. Во-первых, считается, что первичный химический состав склеритов мобергеллид – фосфат кальция, тогда как щитки у современных сипункулид органические или из карбоната кальция. Здесь можно только лишь сделать допущение, что на протяжении длительной эволюционной истории у сипункулид, как и у некоторых других лофотрохозой, например, брахиопод, возникала способность не только к карбонатной, но и к фосфатной биоминерализации. Кроме того, некоторые склериты мобергелл, например, M. radiolata, настолько изменчивы по степени выпуклости (см. Демиденко, 2016), что нельзя исключить предположение, что при жизни они были органическими и пластичными, а их фосфатизация проходила уже посмертно.
Во вторых, следует отметить, что ни анальный, ни каудальный щитки пока не были обнаружены ни у одного из известных видов ископаемых сипункулид. Действительно, сложно объяснить, почему некоторые кембрийские сипункулиды обладали каудальными щитками, а затем палеозойские и мезозойские представители их утратили, и гомологичные структуры появились вновь лишь у рецентных форм. Возможно, это результат слабо известной палеонтологической летописи группы, и в будущем “щитковые” сипункулиды будут распознаны в ископаемом состоянии.
Таким образом, для кембрийских зоопроблематик – мобергеллид мы предлагаем новую гипотезу, альтернативную традиционной – оперкулярной. Мы полагаем, что склериты мобергеллид – это каудальные щитки сипункулид, и кембрийское семейство Mobergellidae Missarzhevsky, 1989 следует относить к типу Sipuncula, классу Phascolosomatidea и, возможно, отряду Aspidosiphonida.
* * *
Авторы благодарны к. б. н. Р.А. Ракитову за помощь при работе на сканирующем электронном микроскопе TESCAN VEGA II кабинета приборной аналитики ПИН РАН.
Коллекция хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН, № 5279. Работа выполнена при поддержке гранта РФФИ № 19-04-01027\19 и по Программе Президиума РАН № 17 “Эволюция органического мира. Роль и влияние планетарных процессов” (подпрограмма I “Развитие жизненных и биосферных процессов”).
Список литературы
Арень Б., Лендзён К. Распространение и стратиграфия климонтовского яруса нижнего кембрия на платформе в Польше // Биостратиграфия и палеонтология нижнего кембрия Европы и Северной Азии. М.: Наука, 1974. С. 30–35.
Вальков А.К. К фауне кессюсинской свиты нижнего кембрия Оленекского поднятия // Тектоника, стратиграфия и литология осадочных формаций Якутии. Якутск, 1968. С. 115–119.
Варламов А.И., Розанов А.Ю., Хоментовский В.В. и др. Кембрий Сибирской платформы. Книга 1: Алдано-Ленский регион. М.: ПИН РАН, 2008. 300 с.
Вестхайде В., Ригер Р. (ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод. М.: КМК, 2008. С. 344–349.
Воронин Ю.И., Воронова Л.Г., Григорьева Н.В. и др. Граница докембрия и кембрия в геосинклинальных областях (опорный разрез Салааны-Гол, МНР). М.: Наука, 1982. 152 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 18).
Гридина Н.М. Фосфатные проблематики в отложениях нижнего кембрия реки Селеты (Центральный Казахстан) // Тез. докл. 3-го Междунар. симпозиума по кембрийской системе. Новосибирск: ИГиГ СО АН СССР, 1990. С. 25.
Дегтярёв К.Е. Тектоническая эволюция раннепалеозойских островодужных систем и формирование континентальной коры каледонид Казахстана. М.: ГЕОС, 2012. 289 с. (Тр. Геол. ин-та РАН. Вып. 602).
Демиденко Ю.Е. Морфология, систематическое положение и стратиграфическое распространение раннекембрийских скелетных проблематик Mobergella radiolata Bengtson, 1968 // Палеонтол. журн. 2016. № 5. С. 10–22.
Дергунов А.Б., Моссаковский А.А., Самыгин С.Г., Хераскова Т.Н. Закономерности формирования палеозоид Центральной Азии (Алтае-Саянская область, Западная Монголия) и Казахстана // Закономерности формирования структур континентов в неогее / Ред. Пущаровский Ю.М. М.: Наука, 1986. С. 53–67.
Догель В.А. Зоология беспозвоночных: Учебник для ун-тов. 7-е изд. / Ред. Ю.И. Полянский. М.: Высш. школа, 1981. 606 с.
Есакова Н.В. Мелкие раковинчатые проблематики // Ископаемые нижнего кембрия гор Маккензи (Канада). М.: Наука, 1987. С. 52–56.
Есакова Н.В., Жегалло Е.А. Фауна и биостратиграфия нижнего кембрия Монголии. М.: Наука, 1996. 216 с. (Тр. Совм. Росс.-Монгол. палеонтол. экспед. Вып. 46).
Ившин Н.К., Булыго Л.В., Гридина Н.М. и др. Стратиграфия и фаунистическое обоснование возраста осадочно-вулканогенных толщ нижнего кембрия северо-востока Центрального Казахстана // Проблемы геологии и металлогении Центрального Казахстана. М.: Наука, 1993. С. 33–38.
Кембрий Сибири / Ред. Репина Л.Н., Розанов А.Ю. Новосибирск: Наука, 1992. 135 с.
Кирсанов В.В. К вопросу о стратиграфии пограничных слоев венда и кембрия в центральных районах Восточно-Европейской платформы // Биостратиграфия и палеонтология нижнего кембрия Европы и Северной Азии. М.: Наука, 1974. С. 5–21.
Конева С.П. Mobergella из нижнего кембрия Селетинского синклинория // Стратиграфия и палеонтология нижнего палеозоя Казахстана. Алма-Ата: Наука, 1983. С. 110–112.
Конева С.П., Попов Л.Е., Ушатинская Г.Т., Есакова Н.В. Беззамковые брахиоподы (акротретиды) и микропроблематики из верхнего кембрия Северо-Восточного Казахстана // Биостратиграфия и палеонтология кембрия Северной Азии. Новосибирск: Наука, 1990. С. 166–170 (Тр. Ин-та геол. и геофиз. СО АН СССР. Вып. 765).
Коpобкин В.В., Cмиpнов А.В. Палеозойcкая тектоника и геодинамика вулканичеcкиx дуг Севеpного Казаxcтана // Геол. и геофиз. 2006. Т. 47. № 4. С. 462–474.
Коробов М.Н., Миссаржевский В.В. О пограничных слоях кембрия и докембрия Западной Монголии (хребет Хасаrт-Хайрхан) // Беспозвоночные палеозоя Монголии. М.: Наука, 1977. С. 7–9.
Менс К., Пиррус Э. Стратотипические разрезы кембрия Эстонии. Таллин: Валгус, 1977. 68 с.
Мешкова Н.П. К вопросу о палеонтологической характеристике нижнекембрийских отложений Сибирской платформы // Биостратиграфия и палеонтология нижнего кембрия Сибири и Дальнего Востока. М.: Наука, 1969. С. 158–174.
Мешкова Н.П. Новые трубчатые проблематики среднего кембрия Сибири и Средней Азии // Проблематики позднего докембрия и палеозоя. М.: Наука, 1985. С. 127–133 (Тр. Ин-та геол. и геофиз. СО АН СССР. Вып. 632).
Миссаржевский В.В. Гастроподы, хиолиты и группы неясного систематического положения нижних горизонтов кембрия // Биостратиграфия и фауна нижних горизонтов кембрия. М.: Наука, 1966. С. 81–111 (Тр. Геол. ин-та АН СССР. Вып. 148).
Миссаржевский В.В. Новые данные о раннекембрийских моноплакофорах // Палеонтол. журн. 1976. № 2. С. 129–131.
Миссаржевский В.В. Древнейшие скелетные окаменелости и стратиграфия пограничных толщ докембрия и кембрия. М.: Наука, 1989. 237 с. (Тр. Геол. ин-та АН СССР. Вып. 443).
Миссаржевский В.В., Мамбетов А.М. Стратиграфия и фауна пограничных слоев кембрия и докембрия Малого Каратау. М.: Наука, 1981. 92 с. (Тр. Геол. ин-та АН СССР. Вып. 326).
Мурина Г.-В.В. Морские черви сипункулиды арктических и бореальных вод Евразии. Л.: Изд-во Наука, 1977. 283 с. (Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР. Т. 111).
Пархаев П.Ю., Демиденко Ю.Е., Кульша М.А. Зоопроблематики Mobergella radiolata как вид-индекс ярусных подразделений нижнего кембрия // Стратигр. Геол. корреляция. 2020. Т. 28. № 2. С. 33–54.
Репина Л.Н., Лазаренко Н.П., Мешкова Н.П. и др. Биостратиграфия и фауна нижнего кембрия Хараулаха. М.: Наука, 1974. 300 с. (Тр. Ин-та геол. и геофиз. СО АН СССР. Вып. 235).
Розанов А.Ю. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука, 1973. 164 с. (Тр. Геол. ин-та АН СССР. Вып. 241).
Розанов А.Ю., Миссаржевский В.В., Волкова Н.А. и др. Томмотский ярус и проблема нижней границы кембрия. М.: Наука, 1969. 380 с. (Тр. Геол. ин-та АН СССР. Вып. 206).
Сухов С.С., Шабанов Ю.Я., Пегель Т.В. и др. Стратиграфия нефтегазоносных бассейнов Сибири. Кембрий Сибирской платформы. Т. 1. Стратиграфия. Новосибирск: ИНГГ СО РАН, 2016. 498 с.
Хоментовский В.В., Карлова Г.А. Венд-кембрийские слои р. Джанды и их аналоги в опорных разрезах Восточной Сибири // Поздний докембрий и ранний палеозой Сибири: Актуальные вопросы стратиграфии. Новосибирск: ИГиГ СО АН СССР, 1989. С. 23–61.
Ярусное расчленение нижнего кембрия. Атлас окаменелостей / Ред. Соколов Б.С., Журавлева И.Т. М.: Наука, 1983. 216 с.
Ярусное расчленение нижнего кембрия. Стратиграфия / Ред. Розанов А.Ю., Соколов Б.С. М.: Наука, 1984. 184 с.
Åhman E., Martinsson A. Fossiliferous Lower Cambrian at Äspelund on the Skäggenäs Peninsula // Geol. Fören. Stockholm Förhandl. 1965. V. 87. P. 139–151.
Alessandrello A., Bracchi G., Riou B. Polychaete, sipunculan and enteropneust worms from the Lower Callovian (Middle Jurassic) of La Voulte-sur-Rhone (Ardeche, France) // Mem. Soc. Ital. Sci. Natur. Museo Civ. Stor. Natur. Milano. 2004. V. 32. № 1. P. 1–16.
Balthasar U., Skovsted C.B., Holmer L.E., Brock G.A. Homologous skeletal secretion in tommotiids and brachiopods // Geology. 2009. V. 37. P. 1143–1146.
Bengtson S. The problematic genus Mobergella from the Lower Cambrian of the Baltic area // Lethaia. 1968. V. 1. P. 325–351.
Bengtson S. Aspects of problematic fossils in the early Palaeozoic // Acta Univ. Upsal. Abstr. Uppsala Dissert. Faculty of Sci. 1977. V. 415. P. 1–71.
Bengtson S., Conway Morris S., Cooper B.J. et al. Early Cambrian fossils from South Australia // Mem. Assoc. Australas. Palaeontol. 1990. № 9. P. 1–364.
Bergström J. Lower Cambrian shelly faunas and biostratigraphy in Scandinavia // Short Pap. 2nd Int. Sympos. Cambrian system. Golden: Colorado School of Mines, 1981. P. 22–25.
Billings E. On some new species of Paleozoic fossils // Cambrian Naturalist. 1871. № 6. P. 213–223, 240.
Brasier M.D. Microfossils and small shelly fossils from the Lower Cambrian Hyolithes Limestone at Nuneaton, English Midlands // Geol. Mag. 1984. V. 121. № 3. P. 229–253.
Brasier M.D. Towards a biostratigraphy of the earliest skeletal biotas // The Precambrian-Cambrian Boundary / Eds. Cowie J.W., Brasier M.D. Oxford: Clarendon Press, 1989. P. 117–165.
Brasier M.D., Anderson M.M., Corfield R.M. Oxygen and carbon isotope stratigraphy of Early Cambrian carbonates in southeastern Newfoundland and England // Geol. Mag. 1992. V. 129. P. 265–279.
Brasier M.D., Singh P. Microfossils and Precambrian–Cambrian boundary stratigraphy at Maldeota, Lesser Himalaya // Geol. Mag. 1987. V. 124. № 4. P. 323–345.
Caron J.-B., Gains R.R., Mángano G.M. et al. A new Burgess Shale–type assemblage from the “thin” Stephen Formation of the southern Canadian Rockies // Geology. 2010. V. 38. № 9. P. 811–814.
Cobbold E.S. The Cambrian horizonts of Comley (Shropshire) and their Brachiopoda, Pteropoda, Gastropoda, etc. // Quart. J. Geol. Soc. Lond. 1921. V. 76. № 304. P. 325–386.
Cobbold E.S., Pocock R.W. The Cambrian area of Rushton (Shropshire) // Phil. Trans. Roy. Soc. Lond. B. 1934. V. 223. P. 305–411.
Conway Morris S., Bengtson S. Cambrian predators: possible evidence from boreholes // J. Paleontol. 1994. V. 68. № 1. P. 1–23.
Conway Morris S., Chapman A.J. Mobergellans from the lower Cambrian of Mongolia, Sweden, and the United States: molluscs or opercula of incertae sedis // J. Paleontol. 1997. V. 71. № 6. P. 968–985.
Cutler E.B. The Sipuncula: their biology, systematics and evolution. Ithaca: Cornell Univ. Press, 1994. 453 p.
Dzik J. Evolution of ‘small shelly fossils’ assemblages of the Early Paleozoic // Acta Palaeontol. Pol. 1994. V. 39. № 3. P. 247–313.
Dzik J. Brachiopod identity of the alleged monoplacophoran ancestors of cephalopods // Malacol. 2010. V. 52. P. 97–113.
Dzik J., Lendzion K. The oldest arthropods of the East European Platform // Lethaia. 1988. V. 21. P. 29–38.
Ehlers E. Ueber fossile Wurmer aus dem lithographischen Schiefer in Bayern // Palaeontogr. 1869. Bd 17. S. 145–175.
Fisher D.W. Small conoidal shells of uncertain affinities // Treatise on Invertebrate Paleontology. Pt W. Miscellanea. Lawrence: Univ. Kansas Press, 1962. P. 98–143.
Geyer G., Landing E. Morocco-2006: Ediacaran–Cambrian depositional environments and stratigraphy of the western Atlas regions // Beringeria. 2006. Spec. № 6. P. 1–121.
Gravestock D.I., Alexander E.M., Demidenko Yu.E. et al. The Cambrian Biostratigraphy of the Stansbury Basin, South Australia. Moscow: MAIK Nauka/Interperiodica, 2001. 344 p.
Hall J. Notes on some new or imperfectly known forms among the Brachiopoda // Ann. Rep. Regents of the Univ. State of New York. 1872. Rep. 23. Appendix G. P. 244–247.
Hamdi B., Brasier M.D., Jiang Zhiwen. Earliest skeletal fossils from Precambrian–Cambrian boundary strata, Elburz Mountains, Iran // Geol. Mag. 1989. V. 126. № 3. P. 283–289.
Hedström H. On “Discinella holsti Mbg” and Scapha & Archaeophiala (some questions on nomenclature) // Sver. Geol. Unders. 1923. V. 314. P. 1–26.
Hedström H. Mobergella versus Discinella; Paterella versus Scapha & Archaeophiala (some questions on nomenclature) // Sver. Geol. Unders. 1930. V. 362. P. 1–8.
Hinz I. The Lower Cambrian microfauna of Comley and Rushton, Shropshire, England // Palaeontogr. A. 1987. Bd 198. № 1/2. S. 41–100.
Huang D.-Y., Chen J.-Y., Vannier J., Saiz Salinas J.I. Early Cambrian sipunculan worms from southwest China // Proc. Roy. Soc. Lond. B. 2004. V. 271. P. 1671–1676.
Jensen S. Trace fossils from the Lower Cambrian Mickwitzia sandstone // Fossil and Strata. 1997. V. 42. P. 1–111.
Jiang Zhiwen, Huang Zhaobi. Middle Cambrian small shelly fauna in Yaxian county, Hainan island // Geol. Rev. 1986. V. 32. № 4. P. 317–324.
Knight J.B. Paleozoic gastropod genotypes // Geol. Surv. Amer. 1941. Spec. pap. 32. P. 1–510.
Kolar-Jurkovsek T., Jurkovsek B. Valvasoria carniolica n. gen. n. sp., a Triassic worm from Slovenia // Geol. Croatica. 1997. V. 50. P. 1–5.
Landing E. Lower Cambrian of Eastern Massachusetts: stratigraphy and small shelly fossils // J. Paleontol. 1988. V. 62. № 5. P. 661–695.
Landing E., Bartowski K.E. Oldest shelly fossils from the Taconic allochthon and Late Early Cambrian sea-levels in Eastern Laurentia // J. Paleontol. 1996. V. 70. № 5. P. 741–761.
Landing E., Geyer G., Bartowski K.E. Latest Early Cambrian small shelly fossils, trilobites, and Hatch Hill dysaerobic interval on the Quebec continental slope // J. Paleontol. 2002. V. 76. № 2. P. 287–305.
Lendzion K. Kambr subholmiowy w polnockowschodniej Polsee // Kwart. Geol. 1972. W. 16. № 13. S. 557–568.
Lendzion K. Fauna of the Mobergella zone in the Polish Lower Cambrian // Kwart. Geol. 1975. T. 19. № 2. S. 237–244.
Lendzion K. Mobergella // Upper Precambrian and Cambrian Palaeontology of the East-European Platform / Eds. Urbanek A., Rozanov A.Yu. Warszawa: Publ. House Wydawnictwa Geol., 1983. P. 121–123.
Lochman Ch. Stratigraphy, paleontology and paleogeography of the Elliptocephala asahoides strata in Cambridge and Hoosick quadrangles, New York // Bull. Geol. Soc. Amer. 1956. V. 64. № 10. P. 1331–1396.
Major H., Winsnes T.S. Cambrian and Ordovician fossils from Sorkapp Land, Spitsbergen // Norsk Polarinst. Skr. 1955. № 106. P. 1–47.
Matthews S.C., Missarzhevsky V.V. Small shelly fossils of late Precambrian and early Cambrian age: A review of recent work // J. Geol. Soc. 1975. V. 131. P. 289–304.
Mens K., Bergström J., Lendzion K. The Cambrian System on the East European Platform. Correlation chart and explanatory notes // Int. Union Geol. Sci. Spec. Publ. 1990. № 25. P. 1–74.
Moberg J.C. Om en nyupptäckt fauna i block af kambrisk sandsten, insamlade av Dr. N.O. Holst // Geol. Fören. Stockholm Förhandl. 1892. V. 14. P. 103–120.
Muir L.A., Botting J.P. A Lower Carboniferous sipunculan from the Granton Shrimp Bed, Edinburgh // Scottish J. Geol. 2007. V. 43. P. 51–56.
Murdock D.J., Bengtson S., Marone F. et al. Evaluating scenarios for the evolutionary assembly of the brachiopod body plan // Evol. & Developm. 2014. V. 16. P. 13–24.
Murdock D.J., Donoghue P.C.J., Bengtson S., Marone F. Ontogeny and microstructure of the enigmatic Cambrian tommotiid Sunnaginia Missarzhevsky, 1969 // Palaeontol. 2012. V. 55. P. 661–676.
Nielsen A.T., Schovsbo N.H. Cambrian to basal Ordovician lithostratigraphy in southern Scandinavia // Bull. Geol. Soc. Denmark. 2007. V. 53. P. 47–92.
Nielsen A.T., Schovsbo N.H. The Lower Cambrian of Scandinavia: depositional environment, sequence stratigraphy and palaeogeography // Earth Sci. Rev. 2011. V. 107. P. 207–310.
Nyers A. Fauna of the basal conglomerate of the Vassbo lead mine (L. Cambrian; NW Dalecarlia, Sweden) // N. Jb. Geol. Paläontol. Monats. 1984. P. 291–299.
Nyers A. Thorslundella: a proposed Early Cambrian protogastropod that secreted a phosphatic shell due to environmental constraints // N. Jb. Geol. Paläontol. Abh. 1987. V. 174. P. 171–192.
Pancucci-Papadopoulou M.A., Murina G.-V.V., Zenetos A. The Phylum Sipuncula in the Mediterranean Sea // Monogr. Mar. Sci. 1999. № 2. P. 1–109.
Poulsen C. The lower Cambrian faunas of East Greenland // Medd. Grønland. 1932. V. 87. P. 1–66.
Poulsen C. Fossils from the Lower Cambrian of Bornholm // Mat. Fys. Medd. Dan. Vidensk. Selsk. 1964. V. 36. № 2. P. 5–48.
Qian Yi. Early Cambrian small shelly fossils of China with special reference to the Precambrian-Cambrian boundary // Stratigraphy and Palaeontology of Systemic Boundaries in China: Precambrian-Cambrian Boundary. Nanjing: Univ. Publ. House, 1989. P. 27–278.
Qian Yi, Jiang Zhiwen. The small shelly fossils from Meshucun stage and stratigraphy of the Lower Cambrian in Yunnan // Acta geol. sin. 1980. V. 54. № 2. P. 150–158.
Rice M.E. Possible boring structures of sipunculids // Amer. zool. 1969. V. 9. P. 803–812.
Rice M.E. A sexual reproduction in a sipunculan worm // Science. 1970. V. 167. P. 1618–1620.
Rice M.E. Chapter 7 – Sipuncula // Microscopic Anatomy of Invertebrates. Vol. 12: Onychophora, Chilopoda, and Lesser Protostomata / Eds. Harrison F.M., Rice M.E. Hoboken: Wiley, 1993. P. 237–325.
Roy S.K., Croneis C. A Silurian worm and associated fauna // Manual of Natural History: Geology IV. Chicago: Field Museum Press, 1931. P. 229–242.
Rozanov A.Yu., Zhuravlev A.Yu. Lower Cambrian fossil records of the Soviet Union // Origin and Early Evolution of the Metazoa / Eds. Lipps J.H., Signor P.W. N.Y.: Plenum Press, 1992. P. 205–281.
Rushton A.W.A. The Cambrian of Wales and England // Cambrian of the British Isles, Norden and Spitsbergen / Ed. C.H. Holland. L.: Wiley, 1974. P. 42–122.
Rushton A.W.A. Chapter 1. General introduction // British Cambrian to Ordovician Stratigraphy / Eds A.W.A. Rushton, A.W. Owen, R.M. Owens, J.K. Prigmore. Peterborough: Joint Nature Conservation Committee, 2000. P. 3–12.
Rushton A.W.A., Molyneux S.G. 7. Welsh Basin // A Revised Correlation of the Cambrian Rocks in the British Isles / Eds A.W.A. Rushton, P.M. Brück, S.G. Molyneux et al. Geol. Soc. L. Spec. Rep. 2011. V. 25. P. 21–27.
Scheltema A.H. Aplacophora as progenetic aculiferans and the coelomate origin of molluscs as the sister taxon of Sipuncula // Biol. Bull. 1993. V. 184. P. 57–78.
Schulze A., Cutler E.B., Giribet G. Reconstructing the phylogeny of the Sipuncula // Hydrobiologia. 2005. V. 535. № 277. https://doi.org/10.1007/s10750-004-4404-3
Selenka E., Man J.G. de, Bulow C. Die Sipunculiden., eine systematische Monographie 2. Wiesbaden, 1883. P. 1–131 (Reisen in Archipel der Philippinen von Dr. C. Semper. V. 4).
Shen X., Ma X., Ren J., Zhao F. A close phylogenetic relationship between Sipuncula and Annelida evidenced from the complete mitochondrial genome sequence of Phascolosoma esculenta // BMC Genomics. 2009. V. 10. № 136. https://doi.org/10.1186/1471-2164-10-136
Skovsted C.B. Mobergellans (Problematica) from the Cambrian of Greenland, Siberia and Kazakhstan // Paläontol. Z. 2003. V. 77. № 2. P. 429–443.
Skovsted C.B. Mollusc fauna of the Early Cambrian Bastion Formation of North-East Greenland // Bull. Geol. Soc. Denmark. 2004. V. 51. P. 11–37.
Skovsted C.B. Small shelly fauna from the Upper Lower Cambrian Bastion and Ella Island formations, North-East Greenland // J. Paleontol. 2006. V. 80. № 6. P. 1087–1112.
Skovsted C.B., Brock G.A., Paterson J.R. et al. The scleritome of Eccentrotheca from the lower Cambrian of South Australia: Lophophorate affinities and implications for tommotiid phylogeny // Geology. 2008. V. 36. P. 171–174.
Skovsted C.B., Brock G.A., Topper T.P. et al. Scleritome construction, biofacies, biostratigraphy and systematics of the tommotiid Eccentrotheca helenia sp. nov. from the early Cambrian of South Australia // Palaeontol. 2011. V. 54. P. 253–286.
Skovsted C.B., Clausen S., Álvaro J.J., Ponlevé D. Tommotiids from the early Cambrian (Series 2, Stage 3) of Morocco and the evolution of the tannuolinid scleritome and setigerous shell structures in stem group brachiopods // Palaeontol. 2014. V. 57. P. 171–192.
Skovsted C.B., Holmer L.E., Larsson C.M. et al. The scleritome of Paterimitra: An early Cambrian stem group brachiopod from South Australia // Proc. Roy. Soc. Lond. B. 2009. V. 276. P. 1651–1656.
Skovsted C.B., Knight I., Balthasar U., Boyce W.D. Depth related brachiopod faunas from the lower Cambrian Forteau Formation of southern Labrador and western Newfoundland, Canada // Palaeontol. Electron. 2017. 20.3.54A. P. 1–52. https://doi.org/10.26879/775
Skovsted C.B., Peel J.S. Small shelly fossils from the argillaceous facies of the Lower Cambrian Forteau Formation of western Newfoundland // Acta Palaeontol. Pol. 2007. V. 52. № 4. P. 729–748.
Skovsted C.B., Peel J.S. Early Cambrian brachiopods and other shelly fossils from the basal Kinzers Formation of Pennsylvania // J. Paleontol. 2010. V. 84. P. 754–762.
Skovsted C.B., Topper T.P. Mobergellans from the early Cambrian of Greenland and Labrador: new morphological details and implications for the functional morphology of mobergellans // J. Paleontol. 2018. V. 92. № 1. P. 71–79.
Smith E.F., Macdonald F.A., Petach T.A. et al. Integrated stratigraphic, geochemical, and paleontological late Ediacaran to early Cambrian records from southwestern Mongolia // Bull. Geol. Soc. Amer. 2016. V. 128. № 3–4. P. 442–468.
Spencer L.M. Paleoecology of a Lower Cambrian archaeocyathid interreef fauna from Southern Labrador. M.Sc. thesis. N.Y.: Univ. N.Y. Stony Brook, 1980. 163 p.
Streng M., Skovsted C.B. A new mobergellan (small shelly fossils) from the early Middle Cambrian of Morocco and its significance // Paläontol. Z. 2006. V. 80. № 3. P. 209–220.
Struck T.H., Paul Ch., Hill N. et al. Phylogenomic analyses unravel annelid evolution // Nature. 2011. V. 471. P. 1–6. https://doi.org/10.1038/nature09864
Topper T.P., Skovsted C.B. Keeping a lid on it: Muscle scars and the mystery of the Mobergellidae // Zool. J. Linn. Soc. 2017. V. 180. P. 717–731.https://doi.org/10.1093/zoolinnean/zlw011
Voss-Foucart M.F., Barzin S., Jeuniaux C., Bussers J.C. Étude comparée de la composition chimique des régions souples et durcies de la cuticule de quatre espèces de sipunculiens // Cahiers Biol. Marine. 1977. V. 18. P. 135–145.
Walcott C.D. Second Contribution to the Studies on the Cambrian Faunas of North America // Bull. US Geol. Surv. 1886. V. 30. P. 1–369.
Walcott C.D. The fauna of the Lower Cambrian or Olenellus Zone // 10th Ann. Rep. US Geol. Surv. 1890. Pt 1. P. 509–774.
Weller S. A new type of Silurian worm // J. Geol. 1925. V. 33. № 5. P. 540–544.
Wrona R. Cambrian Limestone erratics in the Tertiary glacio-marine sediments of King George Island, West Antarctica // Pol. Polar Res. 1989. V. 10. № 4. P. 533–553.
Xing Yusheng, Ding Qixiu, Luo Huilin et al. The Sinian-Cambrian boundary of China and its related problems // Geol. Mag. 1984. V. 121. № 3. P. 155–170.
Yin Jicheng, Ding Lianfang, He Tinggui et al. The Paleontology and Sedimentary Environment of the Sinian System in Emei-Ganluo Area, Sichuan, Yunnan. Sichuan: People’s Publ. House, 1980. 230 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал