

Палеонтологический журнал, 2020, № 5, стр. 102-110
Нитчатые цианобактерии из вендских отложений непского горизонта внутренних районов Сибирской платформы
Е. Ю. Голубкова a, *, Б. Б. Кочнев b, c
a Институт геологии и геохронологии докембрия РАН
Санкт-Петербург, Россия
b Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
c Новосибирский государственный университет
Новосибирск, Россия
* E-mail: golubkovaeyu@mail.ru
Поступила в редакцию 01.11.2018
После доработки 23.08.2019
Принята к публикации 03.09.2019
Аннотация
Проанализировано морфологическое разнообразие и характер распространения нитчатых микрофоссилий в вендских отложениях непского горизонта внутренних районов Сибирской платформы. В составе непской биоты обнаружены гормогониевые цианобактерии пяти родов: Nepia gen. nov., Talakania, Oscillatoriopsis, Obruchevella и Siphonophycus. Проведено монографическое изучение и реконструирован цикл размножения новых осцилляториевых водорослей Nepia calycina Golubkova, gen. et sp. nov. Вид Talakania obscura Kolosov, emend. отнесен к политрихоматозным сине-зеленым водорослям, близким к современным родам Schizothrix и Microcoleus.
ВВЕДЕНИЕ
Несмотря на длительную историю изучения органостенных микрофоссилий, большинство обнаруженных в верхнем докембрии таксонов до сих пор не находят свое место в систематике современных организмов. Сложившаяся ситуация связана, прежде всего, с неполной и зачастую плохой сохранностью палеонтологического материала, что существенно затрудняет проведение палеобиологических исследований. В связи с этим, полученные новые данные имеют важное научное значение, так как позволяют определить систематическое положение некоторых вендских микрофоссилий, отнесенных ранее к формальной подгруппе нитчатых водорослей. В настоящей работе проанализировано таксономическое разнообразие, морфологическое строение, а также распространение нитчатых водорослей, выделенных в результате химического растворения терригенных отложений венда Непско-Ботуобинского, Предпатомского и Сюгджерского районов Восточной Сибири (рис. 1).
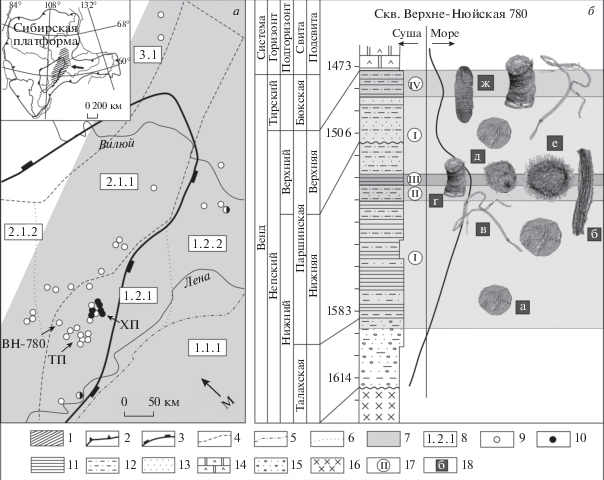
Рис. 1.
Местоположение изученных скважин на схеме структурно-фациального районирования Сибирской платформы (а; Стратиграфия…, 2005) и вертикальное распространение микрофоссилий в разрезе скв. Верхне-Нюйская-780 (б). а: 1 – район исследований; 2 – граница Сибирской платформы; 3 – граница Непско-Ботуобинской антеклизы; 4 – граница фациальных регионов; 5 – граница фациальных районов; 6 – граница фациальных зон; 7 – площадь распространения вендского бассейна; 8 – индексы фациальных регионов, районов, зон: 1 – Байкало-Патомский регион, 1.1 – Патомский район, 1.1.1 – Витимо-Чарская зона, 1.2 – Предпатомский район, 1.2.1 – Нюйско-Пелейдуйская зона, 1.2.2 – Вилючанская зона, 2 – Катангско-Ботуобинский регион, 2.1 – Непско-Ботуобинский район, 2.1.1 – Ботуобинская зона, 2.1.2 – Приленско-Непская зона, 3 – Турухано-Сюгджерский регион, 3.1 – Сюгджерский район; 9 – местонахождения Talakania obscura; 10 – местонахождения Nepia calycina. б: 11 – аргиллиты; 12 – алевролиты; 13 – песчаники; 14 – ангидрито-доломиты; 15 – гравелиты; 16 – фундамент; 17 – ассоциации микрофоссилий: I – с Leiosphaeridia sp., II – c Leiosphaeridia sp.–Talakania obscura, III – c Appendisphaera tenuis, IV –- c Oscillatoriopsis sp.–Siphonophycus sp.; 18 – характерные таксоны: а – Leiosphaeridia sp.; б – Talakania obscura, в – Siphonophycus sp., г – Obruchevella parva, д – Cavaspina acuminata, е – Appendisphaera tenuis, ж – Oscillatoriopsis sp. Принятые сокращения: ВН-780 – скв. Верхне-Нюйская-780, ТП – Талаканская площадь, ХП – Хамакинская площадь, М – направление трансгрессии моря.
Авторы выражают искреннюю признательность К.Е. Наговицину за помощь в фотографировании микрофоссилий. Работа выполнена при финансовой поддержке проекта РФФИ № 17-05-00418 и программы НИР № 0153-2019-0003.
КРАТКИЙ ГЕОЛОГИЧЕСКИЙ ОБЗОР И СТРАТИГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ НИТЧАТЫХ ВОДОРОСЛЕЙ
Благодаря интенсивным микропалеонтологическим исследованиям, в вендских отложениях непского горизонта внутренних районов Сибирской платформы были обнаружены таксономически разнородные ассоциации микрофоссилий, содержащие нитчатые водоросли (Пятилетов, 1980; Колосов, 1983, 1984, 2014; Колосова, 1991; Голубкова и др., 2010; Наговицин, Кочнев, 2015; Станевич и др., 2015). В самом общем приближении верхнедокембрийский осадочный чехол в изученном регионе сложен ?нижне–верхневендскими терригенными последовательностями вилючанского и непского горизонтов, которые выше по разрезу перекрываются верхневендскими карбонатными и эвапоритовыми отложениями тирского и даниловского горизонтов (Стратиграфия …, 2005; Голубкова, Кузнецов, 2014). Граница венда и кембрия проводится в нижней части юряхского подгоризонта даниловского горизонта по появлению мелкораковинных окаменелостей первой зоны томмотского яруса Nochoroicyathus sunnanginicus (Кочнев и др., 2018).
Отложения непского горизонта центральных районов Сибирской платформы накапливались в мелководном морском бассейне, полого погружающемся в юго-восточном направлении. Проведенные палеофациальные реконструкции позволили выделить два трансгрессивно-регрессивных цикла осадконакопления, которые в стратиграфической последовательности отвечают нижнему и верхнему подгоризонтам непского горизонта (рис. 1; Лебедев, Чернова, 1996 и собственные новые данные).
Трансгрессия моря в ранненепское время привела к появлению эврифациальных сфероморфных акритарх Leiosphaeridia crassa (Naumova) emend. Jankauskas, L. jacutica (Timofeev) emend. Mikhailova et Jankauskas, L. minutissima (Naumova) emend. Jankauskas, L. tenuissina Eisenack (рис. 1, ассоциация I), к которым, по мере углубления бассейна, добавились нитчатые водоросли Talakania obscura Kolosov, Obruchevella parva Reitlinger, Siphonophycus Schopf emend. Knoll, Swett et Mark (ассоциация II). Максимуму трансгрессии отвечают наиболее богатые сообщества микрофоссилий (ассоциация III). В разрезах скважин Ботуобинской и Приленско-Непской зон обнаружены разнообразные акритархи (Appendisphaera grandis Moczydłowska, Vidal et Rudavskaya emend. Moczydłowska, A. tenuis Moczydłowska, Vidal et Rudavskaya emend. Moczydłowska, ?A. tabifica Moczydłowska, Vidal et Rudavskaya, Cavaspina acuminata (Kolosova) Moczydłowska, Vidal et Rudavskaya, Ceratosphaeridium Grey, Cucumiforma vanavaria Mikhailova, Leiosphaeridia Eisenack emend. Downie et Sarjeant, Navifusa Combaz, Lange et Pansart, Pterospermopsimorpha insolita Timofeev emend. Mikhailova) и нитчатые водоросли (Obruchevella parva Reitlinger, emend. Yakschin et Luchinina, emend. Burzin, Oscillatoriopsis Schopf, Talakania obscura, Siphonophycus). На западе Нюйско-Пелейдуйской (Хамакинская, Талаканская площади) и в Вилючанской зонах ассоциации микрофоссилий еще более разнообразны. К вышеупомянутым таксонам добавляются акантоморфиты Knollisphaeridium maximum (Yin) Willman et Moczydłowska, Mengeosphaera triangularis Liu, Xiao, Yin, Chen, Zhou et Li, Tanarium conoideum Kolosova emend. Moczydłowska, Vidal et Rudavskaya, T. tuberosum Moczydłowska, Vidal et Rudavskaya, проблематики Hamakinia Rudavskaya, и нитчатые водоросли Nepia calycina gen. et sp. nov. и Obruchevella crassa Kolosov. Такое обилие ископаемых микроорганизмов свидетельствует о благоприятных обстановках, сложившихся в наиболее погруженных частях морского шельфа.
Регрессия и последующая трансгрессия моря в конце ранненепского – начале поздненепского времени способствовали возврату прежних условий, что нашло отражение в последовательной смене комплексов микрофоссилий (рис. 1). Во второй половине непского времени в наиболее глубоководных частях Непско-Ботуобинского (Талаканская, Хамакинская площади) и Предпатомского районов наблюдается постепенная смена комплексов микрофоссилий от моновидового сообщества эврифациальных гладкостенных сфероморфит рода Leiosphaeridia к биотам, содержащим разнообразные орнаментированные акритархи (Appendisphaera, Cavaspina, Ceratosphaeridium, Knollisphaeridium, Tanarium). В составе акантоморфных биот отмечается сокращение количества нитчатых водорослей Talakania obscura вплоть до полного их исчезновения, а их экологическую нишу занимают новые микроорганизмы Nepia calycina gen. et sp. nov. (рис. 1, а). В противоположность этому, в западной части Нюйско-Пелейдуйской и в Приленско-Непской зонах обнаружены лишь эврифациальные микрофоссилии первой ассоциации, что указывает на крайне мелководные условия, сложившиеся здесь в поздненепское время (рис. 1, б). Регрессия моря в конце непского – начале тирского времени, сопровождавшаяся сменой терригенного осадконакопления на карбонатное, привела к исчезновению ассоциаций с акантоморфными акритархами и широкому, но кратковременному развитию микроорганизмов грибной природы Vanavarataenia insolita Pjatiletov и нитчатых водорослей Siphonophycus, Oscillatoriopsis, Obruchevella на всей изученной площади. Последние три таксона, судя по имеющимся палеонтологическим данным, были способны образовывать протяженные корковидные маты. Таким образом, стратиграфическое и латеральное распределение ассоциаций микрофоссилий в отложениях вилючанского, непского и нижней части тирского горизонтов определялось, главным образом, локальными условиями, существовавшими в вендском палеобассейне на каждом этапе его развития.
РЕЗУЛЬТАТЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИЗУЧЕНИЯ
Анализируемые в работе микрофоссилии представлены одиночными и колониальными формами, которые отнесены к пяти родам: Nepia calycina gen. et sp. nov., Obruchevella crassa, O. parva, Oscillatoriopsis sp., Siphonophycus sp., Talakania obscura.
Значительный интерес с точки зрения биологической интерпретации представляют находки новых нитчатых водорослей Nepia calycina. В ископаемом материале присутствуют разнообразные по морфологии нити, которые, как предполагается, отвечают разным стадиям размножения одного организма (табл. XI ; см. вклейку; рис. 2, а). Зрелые нити представляют собой многоклеточные одиночные трихомы, заключенные в плотный чехол – влагалище (табл. XI , фиг. 11 ). Последний редко сохраняется в ископаемом состоянии, что говорит о способности трихомов покидать чехол на определенных этапах своего развития. Предполагается, что при вегетативном размножении нити были способны распадаться на отдельные участки – гормогонии (табл. XI , фиг. 7–9 ). Затем гормогонии отделялись от “материнского” трихома и, совершая колебательные движения, выскальзывали из влагалища, чтобы сформировать новые организмы (табл. XI , фиг. 1–6 ). В ископаемом материале “ювенильные” экземпляры сохраняются в виде коротких бочонковидных или чашевидных трихомов (табл. XI , фиг. 3–6 ), а также сферических тел (табл. XI , фиг. 1, 2). Последние имеют некоторое внешнее сходство с акритархами Leiosphaeridia sp., от которых, однако, отличаются наличием специфических сферических включений и концентрических “складок” вдоль внешнего края этих образований. Форма сохранности “ювенильных” экземпляров определяется их пространственным положением, а также направлением, длительностью и силой воздействия придонного осадка на организм в момент его захоронения (рис. 2, б). Предполагается, что бочонковидные трихомы наиболее приближены к прижизненным формам, в то время как экземпляры с чашевидной или сферической формой были подвержены значительным деформациям. Последние два типа сохранности наиболее часто встречаются в ископаемом материале.
Рис. 2.
Реконструкция цикла вегетативного размножения (а) и формы сохранности (б) гормогониевых цианобактерий Nepia calycina. Oбозначения: т – трихом, к – клетка, ч – чехол, г – гормогония, ц – ?цианофициновые зерна.
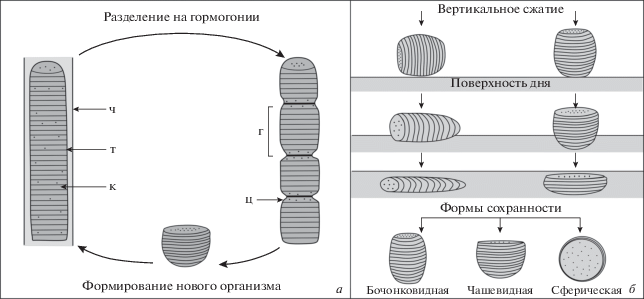
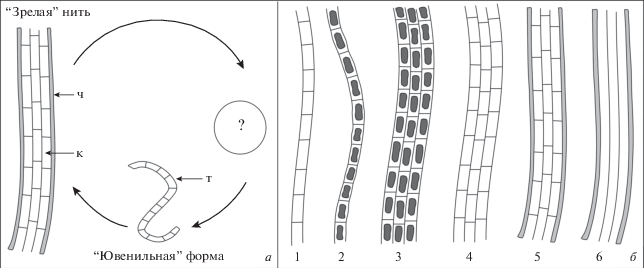
Рис. 3.
Реконструкция цикла размножения (а) и разнообразие форм сохранности (б) цианобактерий Talakania obscura. Oбозначения см. рис. 2.
Представители Nepia calycina содержат мелкие сферические образования (табл. XI , фиг. 1, 9 б). Изучение серии образцов методом электронной микроскопии не выявило присутствия сферических включений на поверхности трихомов (табл. XI , фиг. 12 ), что говорит о нахождении этих форм внутри клеток. Предполагается, что данные образования представляют собой зерна запасных питательных веществ, сопоставимых с цианофициновыми зернами современных сине-зеленых водорослей. На некоторых стратиграфических уровнях водоросли Nepia calycina образуют протяженные маты, составляя до 95% от всего количества выявленных микрофоссилий (табл. XI , фиг. 13 ). По своему морфологическому строению и типу размножения микрофоссилии Nepia calycina сопоставляются с современными осциллаториевыми водорослями, близкими к роду Lyngbia.
Второй анализируемый в статье таксон Talakania был описан П.Н. Колосовым (1983) из вендских отложений непского горизонта внутренних районов Сибирской платформы. На первом этапе изучения в составе рода было выделено два вида – T. diluta Kolosov и T. obscura Kolosov, последний из которых был выбран в качестве типового (Колосов, 1983). Третий вид, T. diversa Kolosov, был описан несколько лет назад из торгинской свиты Березовской впадины юго-востока Сибирской платформы (Колосов, 2014).
Водоросли Talakania демонстрируют высокое морфологическое разнообразие, которое проявляется в плотности прилегания и количестве трихомов в пучке, степени сохранности внутриклеточных образований и клеток вообще, а также в наличии или отсутствии общего чехла (табл. XII , фиг. 1–11 ; см. вклейку). Предполагается, что одиночные многоклеточные трихомы соответствуют “ювенильным” или молодым экземплярам (табл. XII , фиг. 1, 12 б; рис. 3, а, б-1), в то время как пучки многорядных трихомов в чехле отвечают зрелым нитям (табл. XII , фиг. 6 ; рис. 3, а, б-5). Остальные морфотипы представляют собой “переходные” формы между одиночными трихомами и пучками многоклеточных трихомов в чехле (рис. 3, б-2, 3, 4, 6). Их форма сохранности, скорее всего, определялась условиями, существующими в осадке в момент захоронения нитчатых водорослей. Приведенные выше данные позволяют с некоторой долей сомнения сопоставить представителей рода Talakania с политрихоматозными сине-зелеными водорослями, близкими к современным родам Schizothrix и Microcoleus.
Остальные водоросли представлены морфологически просто устроенными одиночными нитями. Многоклеточные гомоцитные трихомы без чехла рода Oscillatoriopsis относят к современным цианобактериям семейства Oscillatoriaceae (Микрофоссилии…, 1989), а свернутые в цилиндрические спирали Obruchevella crassa и O. parva имеют близкое морфологическое строение с гормогониевыми водорослями рода Spirulina (Burzin, 1995). Последний таксон, Siphonophycus sp., представлен нитями бесклеточного строения, которые интерпретируются нами как пустые чехлы или трихомы осцилляториевых водорослей, клетки в которых были разрушены при постмортальном захоронении ископаемых остатков.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ОТДЕЛ CYANOBACTERIA
ПОДКЛАСС NOSTOCOBACTERIDAE
ПОРЯДОК OSCILLATORIALES
Род Nepia Golubkova, gen. nov.
Название рода – по непскому региональному горизонту, в отложениях которого обнаружены описываемые формы.
Типовой вид – Nepia calycina Golubkova, sp. nov.
Диагноз. Однорядные, неветвящиеся многоклеточные гомоцитные трихомы в чехле. Клетки однотипные, короткие, цилиндричеcкой формы. Гормогонии чашевидной, реже бочонковидной формы, короткие. В трихомах содержатся мелкие сферические включения, сконцентрированные, преимущественно, в области сочленения соседних гормогониев. Размножение – вегетативное.
Diagnosis. Single row, unbranced multicellular trichomata, enclosed in a dense sheath. Cells are of similar type, short and cylindrical. Hormogonia are cup-shaped, rarely barrel-shaped, short. Trichomata contain small spherical inclusions, concentrated mainly in the joint area of neighboring hormogonia. Reproduction vegetative.
Видовой состав. Типовой вид.
Сравнение. От представителей рода Oscillatoriopsis Schopf emend. Butterfield отличаются наличием чехла и характерных сферических образований, более короткими клетками, чашевидной формой гормогониев, а также значительно более крупными размерами трихома. От моновидового рода Striatella Assejeva описываемые нитчатые водоросли отличаются формой гормогониев, присутствием сферических образований внутри трихомов, а также более короткими клетками.
Nepia calycina Golubkova, sp. nov.
Табл. XI , фиг. 1–14
Название вида от calycinus лат. – чашевидный.
Голотип – ИГГД РАН, № II-7; Россия, Восточная Сибирь, Байкало-Патомского регион, Предпатомский р-н, Нюйско-Пелейдуйская зона, скв. Нижнехамакинская-21302, гл. 1590 м, обр. 13, пр. 3792/2; ?нижний–верхний венд, непский горизонт, верхняя подсвита паршинской свиты; обозначен здесь, табл. XI , фиг. 8 .
Diagnosis. Single row, unbranched, multicellular trichomata in a sheath. The length of filament fragments is up to 0.8 mm. The width of the filaments are ranges from 48 to 110 μm. Cells are short, uniform in shape and size, up to 2–3 μm in length. Reproduction vegetative. Hormogonia are cup-shaped or barrel-shaped, 30–52 μm in length. Trichomata contain small (about 1 μm in diameter) spherical formations, concentrated mainly in the joint area of neighboring hormogonia.
Описание. Одиночные, неветвящиеся, прямые или слабоизогнутые, многоклеточные трихомы в чехле. Максимальная длина фрагментов нитей в изученном материале составила 0.8 мм. Чехол плотный, редко сохраняется в ископаемом состоянии. Ширина нитей выдерживается на всем протяжении трихома и для разных экземпляров составляет от 48 до 110 мкм. Клетки короткие, одинаковые по форме и размерам, до 2–3 мкм длиной. Поперечные перегородки клеток четкие, сохраняются не всегда. Размножение – вегетативное. Нити распадаются на отдельные участки – гормогонии, которые затем отделяются от трихома и формируют новые организмы. Гормогонии длиной 30–52 мкм, чашевидной или бочонковидной формы. Трихомы содержат многочисленные сферические образования диаметром около 1 мкм, которые располагаются, главным образом, в области сочленения соседних гормогониев, реже вдоль поперечных перегородок клеток. В ископаемом материале встречаются как отдельные, изолированные трихомы, так и фрагменты колоний, сложенные спутанными, незакономерно переплетенными нитями.
Распространение. Восточная Сибирь: паршинская свита Нюйско-Пелейдуйской зоны и харыстанская свита Вилючанской зоны Предпатомского р-на Байкало-Патомского региона; непский горизонт, ?нижний–нижняя часть верхнего венда.
Материал. 126 экз. хорошей и средней сохранности из паршинской свиты, скв. Чайкинская-367 (гл. 1419.7 м), Нижнехамакинская-843 (инт. 1508.7–1499.4 м), -844 (гл. 1706 м), -845 (инт. 1680–1557 м), -849 (инт. 1605–1596 м), ‑21301 (гл. 1592 м), -21302 (гл. 1590 м), и харыстанской свиты, скв. Вилюйско-Джербинская-642 (инт. 2221.6–2169.9 м).
Род Talakania Kolosov, 1983, emend. Golubkova
Типовой вид – Talakania obscura Kolosov, 1983.
Диагноз. Многорядные, однотипные, неветвящиеся трихомы в плотном чехле. Трихомы многоклеточные, перегородки клеток четкие. Клетки одинаковые по морфологии, прямоугольной формы, вытянуты в длину. Внутри клеток часто содержатся темные бесструктурные уплотнения, повторяющие их форму.
Diagnosis emended. Multiseriate, unbranched, multicellular trichomata, enclosed in a dense sheath. Trichomata are multicellular, cell partitions are clearly defined. Cells are of the same morphology, in rectangular shape, elongated. Dark structureless compactions are often observed within the cells.
Видовой состав. Типовой вид.
Сравнение. От многорядных докембрийских водорослей рода Polytrichoides Hermann отличаются клеточным строением трихомов и наличием общего чехла.
Talakania obscura Kolosov, 1983, emend. Golubkova
Табл. XII , фиг. 1–13
Talakania obscura: Колосов, 1983, с. 945, рис. 1, фиг. 1а, б, в; 1984, с. 65, табл. XX , фиг. 4 , табл. XXI , фиг. 1, табл. XXII , фиг. 1; Голубкова и др., 2010, с. 12, табл. II , фиг. 11, 12 ; 2012, с. 593, фиг. 3; Голубкова, Кузнецов, 2014, с. 139, табл. IV , фиг. 6 ; Наговицин, Кочнев, 2015, с. 754, рис. 4 , фиг. I –11.
Talakania diluta: Колосов, 1983, с. 945, рис. 1, фиг. 2; 1984, с. 66, табл. XXII , фиг. 3.
Talakania sp.: Колосова, 1990, табл. 5 , фиг. 2.
Talakania diversa: Колосов, 2014, с. 109, табл. XVIII , фиг. 1
Arctacellularia doliformis: Станевич и др., 2015, с. 129, рис. 3, фиг. 12 .
Arctacellularia sp.: Станевич и др., 2015, с. 129, рис. 3, фиг. 13 .
Голотип – ИГАБМ СО РАН, № 87-82; Россия, Восточная Сибирь, Байкало-Патомский регион, Предпатомский р-н, Нюйско-Пелейдуйская зона, скв. Талаканская-809, инт. 1364.0–1371.2 м, пр. 427-81(б); ?нижний–верхний венд, непский горизонт, паршинская свита (Колосов, 1983, с. 945, рис. 1, фиг. 1а, б, в).
Diagnosis emended. Multiseriate, unbranched trichomata, enclosed in a dense cover. The maximum length of the filament fragments is 4 mm. Trichomata are multicellular. Identical in shape and size rectangular cells are in length up to 2 times of the width or equal to it. The cell wall is clearly defined. The width of the cells is 2.5–7 μm, the length is 3.5–12 μm. Dark, structureless compactions of a unknown origin locate inside the cells. The number of filaments in the bundle is not constant, up to 6–10 in averaged. Type of reproduction is unknown.
Описание. Многорядные, однотипные, прямые или изогнутые, неветвящиеся трихомы, заключенные в плотный чехол, редко сохраняющийся в ископаемом состоянии. Максимальная наблюдаемая длина фрагментов нитей составила 4 мм. Трихомы многоклеточные. Клетки одинаковые по форме и близкие по размерам, прямоугольной формы. Длина клеток в 2 раза превышает ширину, в единичных случаях равна ей. Перегородки клеток четкие. Длина клеток у разных экземпляров изменяется в пределах от 3.5 до 15.0 мкм, ширина – от 2.5 до 7.0 мкм. Внутри клеток часто наблюдаются темные бесструктурные уплотнения неясного генезиса, повторяющие их форму. Количество трихомов в пучке непостоянно, в среднем составляет 6–10 экземпляров. В ископаемом состоянии водоросли сохраняются в виде одиночных трихомов, пучков нитей или скоплений.
Замечания. В составе рода Talakania было выделено три вида: T. obscura, T. diluta и T. diversa (Колосов, 1983, 2014). Отнесенные к этим таксонам водоросли имели близкое морфологическое строение и размеры. Так, длина и ширина клеток у представителей вида T. obscura определялась в интервалах 6.72–18.40 и 2.3–5.6 мкм соответственно. Те же самые параметры для T. diluta составили 7.6–10.4 мкм, 2.40–3.36 мкм и для T. diversa – 5–15 и 1.5–3.1 мкм. Число трихомов в пучке для всех трех видов не превышало восьми. В отличие от T. diluta, типовой вид T. obscura был установлен по наличию темных внутриклеточных уплотнений (Колосов, 1983). Описанный позднее вид T. diversa также содержал темные образования внутри клеток, но отличался “разобщенностью (раздвинутостью, расположением на некотором расстоянии друг от друга) клеток в рядах; различной длиной и толщиной клеток в пределах пучка; не соприкасающимися друг с другом клеточными рядами” (Колосов, 2014, с. 109). Анализ ископаемого материала из типовых местонахождений юго-востока Сибирской платформы позволяет предполагать, что темные внутриклеточные образования, характерные для видов T. obscura и T. diversa, являются “мумифицированными” остатками содержимого клеток, а их присутствие или отсутствие в трихомах определялось особенностями захоронения и последующей консервации ископаемых в толще осадка. С особенностями тафономии также связано неплотное прилегание трихомов друг другу (табл. XII , фиг. 3, 4, 11 ), отличающее представителей вида T. diversa. В случае, если пучок нити в процессе захоронения был “зафиксирован” на растительной пленке (табл. XII , фиг. 11 ), очертание трихома не удается распознать и создается ложное представление о том, что “клетки расположены на некотором расстоянии друг от друга” (Колосов, 2014, с. 109). К такому типу сохранности, по нашему мнению, относится фрагмент нити, предложенный Колосовым в качестве голотипа вида T. diversa (Колосов, 2014, табл. XVIII , фиг. 1). Клетки в пучке несколько отличаются по длине друг от друга не только у описанного Колосовым (2014) вида T. diversa, но и у нитчатых водорослей, отнесенных к T. obscura и T. diluta (напр., табл. XII , фиг. 7 ). Наблюдаемая разница в размерах клеток отражает естественную биологическую изменчивость в пределах вида и не выходит за пределы параметров, установленных для этих нитчатых водорослей. Сказанное выше позволяет объединить выделенные ранее таксоны в один общий вид T. obscura.
Распространение. Восточная Сибирь: харыстанская свита непского горизонта, бюкская свита тирского горизонта Сюгджерского р-на Турухано-Сюгджерского региона; хоронохская, харыстанская свиты непского горизонта Вилючанской зоны, талахская, паршинская свиты непского горизонта Нюйско-Пелейдуйской зоны, торгинская свита Березовской зоны Предпатомского р-на Байкало-Патомского региона; курсовская/паршинская свиты непского горизонта Ботуобинской и Приленско-Непской зон Непско-Ботуобинского р-на Катангско-Ботуобинского региона; ?кессюсинская свита Лена-Анабарского р-на; ?нижний–верхний венд.
Материал. Сотни экземпляров хорошей и средней сохранности из скважин: Вилюйско-Джербинская-642, (инт. 2221.6–2179.8 м, харыстанская свита), -643 (инт. 2483–2136 м, хоронохская, харыстанская свиты), Верхне-Вилючанская-611 (инт. 2204–2184.6 м, харыстанская свита), Буягинская-664 (гл. 1801 м, харыстанская свита), Озерная-761 (инт. 1879.5–1859 м, паршинская свита), Чайкинская-367 (гл. 1419.7 м, паршинская свита), Нижнехамакинская-848 (гл. 1571.5 м, паршинская свита), -844 (гл. 1706 м, паршинская свита), -841 (инт. 1572.9–1570.5 м, паршинская свита), Талаканская-823 (инт. 1542–1536 м, паршинская свита), -824 (гл. 1623 м, паршинская свита), -826 (гл. 1560.8 м, паршинская свита), -809 (инт. 1371.2–1364.1 м, паршинская свита), -815 (инт. 1513.8–1430 м, паршинская свита), -806 (инт. 1590.3–1467 м, паршинская свита), -808 (инт. 1627–1620 м, паршинская свита), -804 (инт. 1676–1669.3 м, паршинская свита), Верхненюйская-780 (инт. 1525.8–1523.8 м, паршинская свита), Верхнечонская-96 (инт. 1666–1645 м, талахская, паршинская свиты), -98 (инт. 1600.5–1587 м, паршинская свита), -20613 (инт. 1619.9–1607 м, паршинская свита), Западная-741 (инт. 1820–1796 м, паршинская свита), -742 (инт. 1886.5–1877.8 м, паршинская свита), Пелейдуйская-750 (гл. 1836 м, паршинская свита), Северо-Юрегинская-1591 (инт. 1962.3–1958.1 м, курсовская свита), Накынская-2950 (инт. 3066.6–3062.5 м, харыстанская свита), Дюданская-2910 (инт. 3415.3–3414.8 м, нижняя подсвита бюкской свиты), Усть-Бирюкская-1 (инт. 3101–3120 м, нижняя подсвита торгинской свиты).
ЗАКЛЮЧЕНИЕ
Проведенные палеонтологические исследования показали, что подавляющая часть нитчатых водорослей (Talakania, Oscillatoriopsis, Obruchevella, Siphonophycus) имеют широкое вертикальное и латеральное распространение в вендских отложениях непского горизонта внутренних районов Сибирской платформы. Они обнаружены во всех обстановках, за исключением крайнего мелководья, где фиксируются лишь редкие планктонные эврифациальные акритархи Leiosphaeridia. Ограниченное площадное распространение имеют водоросли Nepia calycina, выявленные в наиболее глубоководных частях непского бассейна.
Анализируемые в работе микрофоссилии сопоставляются с наиболее консервативной группой сине-зеленых водорослей (цианобактерий). Для новых нитчатых водорослей Nepia calycina проведено монографическое изучение и реконструирован цикл размножения, что позволило отнести их к современным осцилляториевым водорослям, близким к роду Lyngbia. Ископаемые микроорганизмы, выделенные ранее в формальный род Talakania, сопоставляются с политрихоматозными водорослями, морфологически близкими современным родам Schizothrix и Microcoleus. Полученные палеонтологические данные могут быть использованы в дальнейшем для проведения палеоэкологических и палеофациальных исследований.
Список литературы
Голубкова Е.Ю., Кузнецов А.Б. Таксономическое разнообразие и корреляция эдиакарских комплексов акантоморфной палинофлоры мира // Водоросли в эволюции биосферы / Ред. Рожнов С.В. М.: ПИН РАН, 2014. С. 119–140.
Голубкова Е.Ю., Раевская Е.Г., Ивановская А.В. Закономерности латерального распространения микрофоссилий в вендских нефтегазоносных отложениях Непского свода Непско-Ботуобинской антеклизы Восточной Сибири // Проблемы зарождения и эволюции биосферы. Ч. 2 / Pед. Э.М. Галимов. М.: УРСС, 2012. С. 591–595.
Голубкова Е.Ю., Раевская Е.Г., Кузнецов А.Б. Нижневендские комплексы микрофоссилиий Восточной Сибири в решении стратиграфических проблем региона // Стратигр. Геол. корреляция. 2010. Т. 18. № 4. С. 3–27.
Колосов П.Н. Нитчатые микрофоссилии в курсовской свите венда Якутии // Докл. Акад. наук. 1983. Т. 269. № 4. С. 944–946.
Колосов П.Н. Позднедокембрийские микроорганизмы востока Сибирской платформы. Якутск: ЯФ СО АН СССР, 1984. 84 с.
Колосов П.Н. Несколько видов микрофоссилий из торгинской свиты неопротерозоя Березовской впадины // Палеонтол. журн. 2014. № 3. С. 105–112.
Колосова С.П. Древнейшие акантоморфиты востока Сибирской платформы // Органический мир и стратиграфия отложений нефтегазоносных и рудоносных районов Якутии. Якутск: Якутский ин-т геол. наук СО АН СССР, 1990. С. 2–45.
Кочнев Б.Б., Покровский Б.Г., Кузнецов А.Б., Марусин В.В. C- и Sr-хемостратиграфия карбонатных отложений венда-нижнего кембрия центральных районов Сибирской платформы // Геол. и геофизика. 2018. Т. 59. № 6. С. 731–755.
Лебедев М.В., Чернова Л.С. Фациальные модели терригенных отложений венда северо-востока Непско-Ботуобинской антеклизы (Сибирская платформа) // Геол. и геофизика.1996. Т. 37. № 10. С. 51–64.
Микрофоссилии докембрия СССР / Ред. Янкаускас Т.В. Л.: Наука, 1989. 190 с.
Наговицин К.Е., Кочнев Б.Б. Микрофоссилии и биофации вендской ископаемой биоты юга Сибирской платформы // Геол. и геофизика. 2015. Т. 56. № 4. С. 748–760.
Пятилетов В.Г. Юдомский комплекс микрофитофоссилий Южной Якутии // Геол. и геофизика. 1980. № 7. С. 8–20.
Станевич А.М., Корнилова Т.А., Мазукабзов А.М., Вахромеев А.Г. Корреляция и микрофоссилии венд–кембрийских отложений, вскрытых Чайкинской параметрической скважиной 279 // Изв. Иркутск. гос. ун-та. Сер. Науки о Земле. 2015. Т. 12. С. 124–135.
Стратиграфия нефтегазоносных бассейнов Сибири. Рифей и венд Сибирской платформы и ее складчатого обрамления / Ред. Конторович А.Е. Новосибирск: Гео, 2005. 432 с.
Burzin M.B. Late Vendian helicoids filamentous microfossils // Paleontol. J. 1995. V. 29. № 1A. P. 1–34.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал