

Палеонтологический журнал, 2020, № 5, стр. 38-48
О новых находках челюстей цефалопод в верхнем байосе (средняя юра) Северного Кавказа (Карачаево-Черкесия)
А. А. Мироненко a, *, В. В. Митта b, c, **
a Геологический институт РАН
Москва, Россия
b Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
c Череповецкий государственный университет
Череповец, Россия
* E-mail: paleometro@gmail.com
** E-mail: mitta@paleo.ru
Поступила в редакцию 19.07.2019
После доработки 01.08.2019
Принята к публикации 01.08.2019
Аннотация
Описываются новые находки остатков челюстных аппаратов цефалопод из верхней части зоны Niortense и нижней части зоны Parkinsoni междуречья Кубани и Урупа. Изолированные створки двух аптихов, трактуемых как нижние челюсти аммонитов, отнесены к надсемействам Haploceratoidea и Stephanoceratoidea. Две верхние челюсти предположительно принадлежали также аммонитам с аптиховым типом челюстного аппарата (Ammonitida). Семь челюстей анаптихового типа отнесены к отрядам Phylloceratida и Lytoceratida. Впервые из мезозоя Северного Кавказа описана хорошо сохранившаяся верхняя челюсть колеоидеи.
ВВЕДЕНИЕ
Цефалоподы, в отличие от других моллюсков, обладают хорошо развитым челюстным аппаратом, состоящим из двух челюстей (верхней и нижней) и расположенной между ними радулы. Челюсти большинства современных головоногих моллюсков состоят исключительно из органического вещества, и лишь у доживших до наших дней представителей подкласса Nautiloidea в кончиках челюстей имеются кальцитовые элементы (Saunders et al., 1978; Tanabe et al., 2015a). Однако в прошлом разнообразие вариантов челюстей головоногих моллюсков было существенно выше, в первую очередь за счет представителей подкласса Ammonoidea, у которых исследователи выделяют от четырех до шести различных типов строения челюстного аппарата (Tanabe et al., 2015a; Mitta, Schweigert, 2016). Особенно важным для понимания эволюции челюстного аппарата аммоноидей является юрский период. В конце ранней юры (тоар) у представителей подотряда Ammonitina возник аптиховый тип челюстного аппарата, в котором нижняя челюсть состояла из двух половинок, часто покрытых кальцитовым слоем и соединенных между собой гибкой связкой (Engeser, Keupp, 2002; Tanabe et al., 2015a). Вероятно, в ранней юре появился и ринхаптиховый тип челюстного аппарата, в котором кончики челюстей (как и у наутилид) были усилены кальцитовыми элементами. Находки изолированных кальцитовых элементов челюстей головоногих моллюсков (ринхолитов) известны, начиная с верхнего триаса, но они, скорее всего, принадлежали наутилидам (Riegraf, Luterbacher, 1989; Riegraf, Schmitt-Riegraf, 1995). Древнейшие ринхолиты, предположительно принадлежавшие аммоноидеям, известны, начиная с плинсбаха (Riegraf, Luterbacher, 1989), а самые древние на сегодняшний день челюсти ринхаптихового типа найдены в средней юре (байос) Северного Кавказа (Mironenko, Gulyaev, 2018).
Среднеюрский этап эволюции челюстей цефалопод в течение долгого времени оставался малоизученным. Остатки челюстных аппаратов среднеюрских цефалопод, описанные в литературе, преимущественно представлены классическими двураздельными аптихами отряда Ammonitida (Trauth, 1930; Lehmann, 1972, 1978; Morton, 1973, 1975). Находки среднеюрских анаптихов ограничивались двумя экземплярами (Quilty, 1970; Westermann et al., 1999), причем в первом случае принадлежность находки к челюстным аппаратам цефалопод вызывает сомнения. Однако в последние два десятилетия опубликовано множество новых работ, посвященных среднеюрским аптихам (Рогов, Гуляев, 2003; Рогов, 2004; Mitta, Keupp, 2004; Митта, 2009, 2017б; Dietze et al., 2012; Keupp, Mitta, 2013; Mitta, Schweigert, 2016; Митта, Шерстюков, 2018; Mitta et al., 2018), а также анаптихам и ринхаптихам (Mironenko, Gulyaev, 2018). Также были описаны новые находки челюстей среднеюрских колеоидей (Keupp, Mitta, 2013). Большая часть этих публикаций основана на находках из Центральной России и с Северного Кавказа.
В течение ряда последних лет один из авторов (В.В. Митта) производил полевые работы на байос–батских разрезах Северного Кавказа, в бассейне р. Кубань (Карачаево-Черкесская республика). Были опубликованы две находки нижних челюстей аммонитов из зоны Strenoceras niortense (Mitta, Schweigert, 2016; Митта, 2017б). В последующие годы в верхнем байосе междуречья Кубани и Урупа были найдены новые остатки челюстных аппаратов цефалопод, описываемые ниже.
МЕСТОНАХОЖДЕНИЯ И СТРАТИГРАФИЯ
Верхний байос р. Кяфар представлен средней частью джангурской свиты (байос–нижний бат: Безносов, 1967), в составе которой преобладают глины и аргиллиты с рассеянными в толще и нередко образующими прослои конкрециями различного состава, с подчиненными прослоями алевролитов и известняков. Здесь установлены все три стандартные зоны (хронозоны) верхнего байоса: Strenoceras niortense, Garantiana garantiana и Parkinsonia parkinsoni (Митта, Шерстюков, 2014), последовательно вскрывающиеся вверх по реке от более молодых к более древним. Верхняя часть берегового склона обычно сложена четвертичными галечниками, срезающими юрские отложения по всей долине. Челюсти цефалопод найдены в нескольких местонахождениях в зоне Niortense и в одном случае в зоне Parkinsoni (рис. 1).
Рис. 1.
Расположение местонахождений: а – общая схема, рамкой показан контур детальной карты; б – детальная карта.
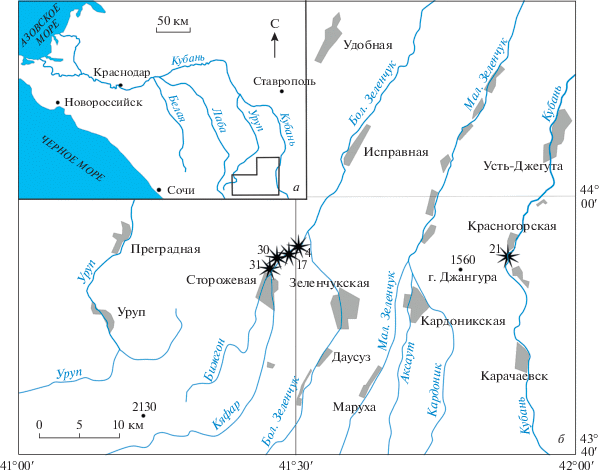
Зона Niortense представлена в бассейне р. Большой Зеленчук глиной темно-серой песчанистой с многочисленными прослоями карбонатных конкреций и маломощными (0.2–0.4 м) конкреционными прослоями и линзами алевролитов. В овраге по левому берегу р. Кяфар у станицы Сторожевая (местонахождение № 31) в этих породах (слои с rostovtsevi) ранее была найдена изолированная створка аптиха (Митта, 2017б, рис. 3). По форме и размерам этот аптих мог принадлежать только Orthogarantiana rostovtsevi (Mitta) (подсем. Garantianinae семейства Stephanoceratidae), многочисленные раковины которого были найдены в этом же слое.
Рис. 2.
Челюсти аммоноидей из местонахождения № 17: а, б – анаптих ПИН, № 5546/207: а – спереди, б – сбоку; в, г – аптих ПИН, № 5546/206: в – экз. ПИН, № 5546/206а, отпечаток аптиха на породе, г – экз. ПИН, № 5546/206б, створка аптиха в породе; д – экз. ПИН, № 5546/205, левая половина анаптиха; е – экз. ПИН, № 5546/209, небольшой анаптих; ж, з – экз. ПИН, № 5546/208, частично сохранившийся анаптих: ж – спереди, з – сверху. Длина масштабных отрезков а, б, д, ж, з – 5 мм, в, г, е – 2.5 мм.

Рис. 3.
Микроструктура органического слоя внешней пластины анаптиха, экз. ПИН, № 5546/207, длина масштабного отрезка: а – 1 мм, б – 300 мкм. Снимки изготовлены на сканирующем электронном микроскопе Tescan Vega, ПИН РАН.

Следующее местонахождение с остатками челюстных аппаратов (№ 30) расположено на левом берегу Кяфара ниже по реке. Из конкреции алевролита серого, с поверхности лимонитизированного красновато-бурого происходят три находки изолированных неполно сохранившихся частей челюстей цефалопод (ПИН, №№ 5546/212, 5546/252, 5546/253). На этом уровне, кроме транзитных видов Phylloceratina и Lytoceratina, найдены многочисленные Garantiana ex gr. baculata (Quenstedt), позволяющие сопоставить вмещающие слои с верхней подзоной Baculata зоны Niortense.
Местонахождение № 17 расположено на правом берегу р. Кяфар немного ниже по течению. Здесь в жилой камере Calliphylloceras ранее были найдены нижние челюсти, описанные как новый тип челюстного аппарата аммоноидей – филлаптих (Mitta, Schweigert, 2016). Находка была сделана в конкреции алевролита серого, с поверхности желтовато-серого, с многочисленными обугленными растительными остатками. Из такой же конкреции в этом обнажении, найденной на том же уровне, были описаны разнообразные гетероморфные аммониты рода Spiroceras (Митта, 2017a).
Большинство конкреций этого прослоя алевролитов не содержит ископаемых. Иногда встречаются конкреции, содержащие растительные остатки, единичные или многочисленные. Еще реже встречаются конкреции, переполненные как остатками растений (вайи сагеноптерис, реже фрагменты хвощевых), так и моллюсков – аммонитов, белемнитов, двустворок и, совсем редко, гастропод. В 2017 г. в результате целенаправленных поисков были найдены три такие конкреции, в одной из которых нашлись остатки челюстных аппаратов цефалопод. В данной работе мы описываем семь находок челюстей цефалопод из этого местонахождения (ПИН, №№ 5546/205, 5546/206, 5546/207, 5546/208, 5546/209, 5546/211, 5546/254). Уточненный перечень таксонов аммонитов из подобных конкреций включает представителей отрядов Ammonitida – родов Oecotraustes (Oppeliidae), Lissoceras (Lissoceratidae), Sphaeroceras (Sphaeroceratidae), Spiroceras (Spiroceratidae), Garantiana и Pseudogarantiana (Stephanoceratidae), Leptosphinctes (Perisphinctidae), а также Phylloceratida – родов Calliphylloceras, Holcophylloceras, Pseudophylloceras, Adabofoloceras, и Lytoceratida – родов Nannolytoceras и Megalytoceras.
Выше по разрезу найдены также характерные для верхней подзоны зоны Niortense таксоны аммонитов. Таким образом, челюстные остатки из всех трех местонахождений (№№ 31, 30, 17) происходят из разных стратиграфических уровней подзоны Baculata зоны Niortense.
Ниже по р. Кяфар обнажаются глины аргиллитоподобные зоны Garantiana garantiana, где найдены редкие аммониты и белемниты (Митта, 2018), но челюстные остатки не встречены. Еще ниже по течению исследованы разрезы подзоны Rarecostites subarietis зоны Parkinsoni, представленной аргиллитами с рассеянными в толще и нередко образующими прослои сидеритовыми и карбонатными конкрециями. В одной из этих конкреций в местонахождении № 4 обнаружена челюсть цефалоподы (ПИН, № 5546/210). В интервале находки (фаунистический горизонт sherstyukovi), кроме многочисленных Rarecostites (сем. Parkinsoniidae) и единичной раковины Patrulia (сем. Stephanoceratidae), найдены Spiroceras, Holcophylloceras, Calliphylloceras, Adabofoloceras, Dinolytoceras и Nannolytoceras (Митта, 2017в; Mitta, Sherstyukov, 2018).
На восточном склоне горы Джангура в разрезе у ст. Красногорская зона Niortense представлена глинами с прослоями и линзами алевролитов и песчаниками, иногда переполненными ископаемыми, общей мощностью около 70 м (Затворницкий, 1914; Кахадзе, Зесашвили, 1956; Безносов, Митта, 1998). В нижней части этой толщи в местонахождении 21 найден фрагмент челюсти (экз. ПИН, № 5546/255).
МАТЕРИАЛ
Авторами было изучено 14 образцов, относящихся к 12 экз. ископаемых челюстей цефалопод. Оригиналы хранятся в Палеонтологическом ин-те им. А.А. Борисяка (ПИН) РАН, колл. № 5546.
Образцы из местонахождения № 17
Экз. ПИН, № 5546/207 (рис. 2, а, б) – небольшой анаптих хорошей сохранности, с остатками органического слоя, но без кальцитовых элементов. Органический слой имеет характерную сетчатую микроструктуру (рис. 3). Похожая микроструктура обнаружена и на поверхности нижних челюстей аммоноидей из зоны Parkinsoni Дагестана (Mironenko, Gulyaev, 2018, рис. 6).
Экз. ПИН, № 5546/206 (рис. 2, в, г) – левая створка аптиха с противоотпечатком. Образец № 5546/206а (рис. 2, в) – отпечаток аптиха на породе, вид сверху; № 5546/206б (рис. 2, г) – аптих в породе, вид снизу. Аптих небольшой, слабовыпуклый и удлиненный, скульптура его поверхности состоит из частых тонких ребер.
Экз. ПИН, № 5546/205 (рис. 2, д) – левая створка анаптиха посредственной сохранности (противоположная правая часть анаптиха скрыта в породе). Хорошо видна ребристая скульптура поверхности анаптиха, кальцитовых элементов нет.
Экз. ПИН, № 5546/209 (рис. 2, е) – небольшой анаптих средней сохранности, следов кальцитового слоя и конхоринха нет. Задние края анаптиха, особенно в его левой части, сохранились плохо.
Экз. ПИН, № 5546/208 (рис. 2, ж, з) – анаптих неполной сохранности во фрагменте конкреции, содержащем мелкие раковины Spiroceras и растительные остатки. Сохранившаяся правая часть анаптиха длиннее, но более узкая, чем левая часть. Наружного кальцитового слоя и конхоринха нет.
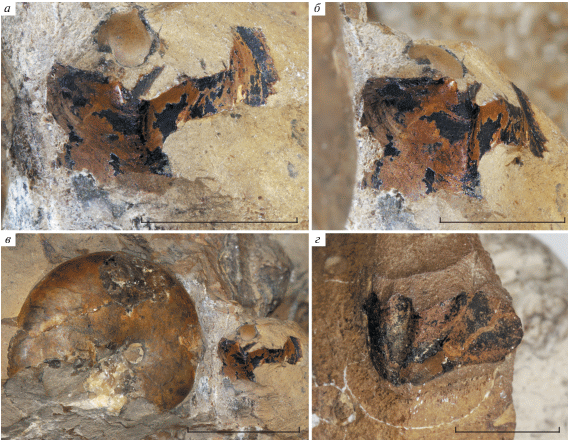
Экз. ПИН, № 5546/211 (рис. 4) – клювообразная верхняя челюсть колеоидеи, узкая и вытянутая, со слегка изогнутым и острым передним концом. Видна внешняя пластина челюсти, внутренняя не сохранилась или скрыта в породе.
Рис. 4.
Верхняя челюсть колеоидеи из местонахождения № 17, экз. ПИН, № 5546/211: а – слева, б – сверху, в – справа. Длина масштабных отрезков 5 мм.

Экз. ПИН, № 5546/254 (рис. 5) – верхняя челюсть хорошей сохранности во фрагменте конкреции, содержащем многочисленные мелкие раковины Spiroceras и растительные остатки. Образец № 5546/254a – наружная пластина, № 5546/254b – внутренняя пластина.
Рис. 5.
Верхняя челюсть аммоноидеи из местонахождения № 17: а – экз. ПИН, № 5546/254a, наружная пластина, снизу; б – экз. ПИН, № 5546/254b – внутренняя пластина, сверху. Длина масштабных отрезков 2.5 мм.

Рис. 6.
Челюсти аммоноидей из местонахождения № 30: а–в – анаптих, экз. ПИН, № 5546/212: а – сбоку, б – сверху, в – левая половина внешней пластины анаптиха; г – фрагмент аптиха, экз. ПИН, № 5546/252. Длина масштабных отрезков 5 мм.

Образцы из местонахождения № 30
Экз. ПИН, № 5546/212 (рис. 6, а, б) – крупный, частично сохранившийся анаптих с остатками органического слоя, без кальцитовых элементов.
Экз. ПИН, № 5546/253 (рис. 6, в) – левая половина внешней пластины нижней челюсти аммоноидеи. Внутренняя пластина не сохранилась, в месте стыка пластин в средней части имеется отверстие.
Экз. ПИН, № 5546/252 (рис. 6, г) – передняя часть аптиха с остатками органического слоя, без кальцитового слоя, задняя часть не сохранилась. Аптих сильно выпуклый, удлиненный и не имеет выраженной скульптуры.
Образец из местонахождения № 4
Экз. ПИН, № 5546/210 (рис. 7, а–в) – анаптих неполной сохранности, расположен в конкреции рядом с раковиной Adabofoloceras ? sp. juv. (рис. 7, в). Левая сторона внешней пластины и центральная часть анаптиха разрушены, за счет этого хорошо видно более короткую внутреннюю пластину. На поверхности наружной пластины, как и на образце № 5546/207, видна сетчатая микроструктура. Кальцитовых элементов нет.
Рис. 7.
Челюсти аммоноидей: а–в – анаптих, экз. ПИН, № 5546/210, местонахождение № 4: а – спереди, б – сбоку, в – общий вид образца с аммонитом; г – экз. ПИН, № 5546/255, передняя часть внутренней пластины верхней челюсти аммоноидеи, местонахождение № 21. Длина масштабных отрезков 5 мм (а, б, г) и 10 мм (в).
Образец из местонахождения № 21
Экз. ПИН, № 5546/255 (рис. 7, г) – передняя часть внутренней пластины верхней челюсти аммоноидеи в небольшом фрагменте конкреции.
ОБСУЖДЕНИЕ
Элементы челюстных аппаратов головоногих моллюсков, описанные выше, принадлежат представителям двух подклассов цефалопод: Ammonoidea и Coleoidea. К настоящему времени из байоса Северного Кавказа были описаны челюсти аммоноидей, относящиеся к четырем различным типам челюстных аппаратов: аптиховому (Митта, 2017б), анаптиховому и ринхаптиховому (Mironenko, Gulyaev, 2018), и известному пока только по одному экземпляру филлаптиховому (Mitta, Schweigert, 2016). Большинство находок, описываемых в данной работе, относятся к нижним челюстям аммоноидей анаптихового типа (анаптихам), которые принадлежали аммоноидеям подотрядов Phylloceratina и Lytoceratina. Анаптихи состоят из двух пластин (более крупной округлой внешней пластины и небольшой внутренней), соединяющихся в середине передней части челюсти. В отличие от двустворчатых аптихов, они не имеют симфиза в центральной части. В некоторых случаях внешние пластины анаптихов могут сохраняться отдельно, вероятно, в результате разрушения более тонких внутренних пластин до захоронения в донном осадке. В таком случае в их передней части наблюдается зияние, как у экз. ПИН, № 5546/253.
Челюсти ринхаптихового типа (ринхаптихи), отличающиеся присутствием кальцитовых элементов (ринхолитов и конхринхов) в заостренной передней части и, в некоторых случаях, наличием внешнего кальцитового слоя, недавно были описаны из зоны Parkinsoni верхнего байоса Дагестана (Mironenko, Gulyaev, 2018). Однако в байосе Карачаево-Черкесии они обнаружены не были. Филлоцератины Adabofoloceras, Holcophylloceras и Pseudophylloceras, которым предположительно принадлежали дагестанские ринхаптихи, встречены и в разрезах верхнего байоса Карачаево-Черкесии, поэтому отсутствие ринхаптихов в этих отложениях нельзя объяснить различием фаунистического состава. Древнейшие кальцитовые элементы аммонитовых челюстей ринхаптихового типа известны, начиная с плинсбаха, встречаются они и в отложениях тоара и аалена (Riegraf, Luterbacher, 1989). Хотя в разных эволюционных линиях аммоноидей ринхаптихи возникали не одновременно на протяжении ранней и средней юры, представляется маловероятным, что на Северном Кавказе они появились в интервале между хронами Niortense и Parkinsoni, хотя такой вариант не может быть полностью исключен. Однако более вероятно, что отсутствие кальцитовых элементов в нижних челюстях аммоноидей из зоны Niortense связано с тафономическими факторами: они либо отделились от органической части челюсти до захоронения, либо растворились несколько позже. Возможность потери кальцитовых элементов до захоронения выглядит тем более вероятной, что почти все изученные образцы челюстей имеют неполную сохранность, их внешние края несут заметные следы эрозии. Таким образом, часть анаптихов, описанных в данной статье, может на самом деле быть ринхаптихами, но при отсутствии кальцитовых элементов различить их не представляется возможным. В любом случае, эти нижние челюсти анаптихового или ринхаптихового типов принадлежали аммоноидеям подотрядов Phylloceratina и Lytoceratina. Точнее связать находки с теми или иными родами не представляется возможным, за исключением обр. ПИН, № 5546/210, расположенного рядом с юным экземпляром Adabofoloceras ? sp.; возможно, что данный анаптих является нижней челюстью именно этого аммонита.
В зоне Niortense найдены три аптиха, один из которых был описан ранее (Митта, 2017б, рис. 3). Кальцитовый слой на этих образцах также отсутствует. Обращает на себя внимание редкость находок аптихов, при том, что обладавшие челюстным аппаратом аптихового типа аммониты подотряда Ammonitina чрезвычайно многочисленны в изученных слоях, и их количество значительно превышает число литоцератин и филлоцератин. Из вышележащих зон верхнего байоса Северного Кавказа, несмотря на многочисленность аммонитов семейства Parkinsoniidae, обладавших аптиховым типом челюстного аппарата (Wetzel, 1911; Lehmann, 1978), находки аптихов пока неизвестны. Вероятно, такая редкость аптихов связана с меньшей толщиной их органического слоя по сравнению с анаптихами/ринхаптихами и, как следствие, худшим фоссилизационным потенциалом.
Вытянутая форма аптиха ПИН, № 5546/206 (рис. 2, в, г) и его ребристая поверхность свидетельствуют о возможной принадлежности этого аптиха к надсемейству Haploceratoidea (роду Lissoceras сем. Lissoceratidae, или роду Oppelia сем. Oppeliidae). Фрагментарно сохранившийся аптих ПИН, № 5546/252 (рис. 6, г), судя по его форме, принадлежал к надсемейству Stephanoceratoidea (подсем. Garantianinae семейства Stephanoceratidae). В пользу этого предположения свидетельствуют и многочисленные раковины Garantiana ex gr. baculata (Quenstedt), захороненные на поверхности плитки алевролита с этим аптихом (рис. 8).
Рис. 8.
Часть плитки алевролита с многочисленными Garantiana ex gr. baculata (Quenstedt) и другими ископаемыми, включая частично сохранившийся аптих (рис. 6, г); экз. ПИН, № 5546/252; местонахождение № 30. Длина масштабного отрезка 20 мм.

Верхняя челюсть ПИН, № 5546/211 (рис. 4), заостренная и сильно сжатая с боков, несомненно, принадлежала головоногому моллюску подкласса Coleoidea. Челюсти колеоидей встречаются в ископаемом состоянии значительно реже, чем челюстные аппараты аммоноидей, и из мезозоя Северного Кавказа они до сих пор описаны не были. В целом форма верхней челюсти из зоны Niortense, узкая и длинная, говорит о ее принадлежности десятируким колеоидеям (Decapodiformes). Верхние челюсти колеоидей очень похожей формы известны из верхнего мела (турон и сантон) Японии (Tanabe et al., 2006), правда, их размер существенно превышает длину нашего экземпляра. Они интерпретированы как челюсти ископаемых теутид (Tanabe et al., 2015b). Из местонахождения № 17 определены ростры белемнитов подотрядов Belemnopseina (роды Dicoelites и Belemnopsis) и Belemnitina (род Megateuthis) (Dzyuba et al., 2019), и вполне вероятно, что челюсть принадлежала одному из этих моллюсков. С другой стороны, нельзя исключать их принадлежность к безростровым колеоидеям, гладиусы которых плохо сохраняются в ископаемом состоянии (Fuchs, Iba, 2015).
Две верхние челюсти (ПИН, №№ 5546/254, рис. 5, и 5546 /255, рис. 7, г) заметно отличаются от вышеописанной челюсти колеоидеи. Они шире и короче, с закругленным передним концом. Ранее подобные челюсти из нижнего келловея реки Унжа были интерпретированы как челюсти цирратных осьминогов (Keupp, Mitta, 2013). Однако их детальное изучение показывает существенные отличия не только от челюстей осьминогов, но и от челюстей колеоидей в целом. Так, их внутренняя и внешняя пластины имеют одинаковую длину, а в центральной части между пластинами находится образование, похожее на вытянутую трубку. Подобная структура не была известна у современных или ископаемых головоногих. Но недавно была описана верхняя челюсть позднеюрского аммонита Kachpurites fulgens (Perisphinctoidea: Craspeditidae), передняя часть которой имеет сходную форму (Мироненко, 2014). Логично предположить, что две находки из зоны Niortense также являются верхними челюстями аммоноидей с аптиховым типом челюстного аппарата. В отличие от нижних челюстей (аптихов) таких аммонитов, которым посвящено множество публикаций, их верхние челюсти до сих пор остаются недостаточно изученными.
ЗАКЛЮЧЕНИЕ
Среди новых находок остатков челюстных аппаратов цефалопод из верхов зоны Niortense и низов зоны Parkinsoni верхнего байоса Северного Кавказа преобладают нижние челюсти анаптихового типа, характерные для аммоноидей подотрядов Phylloceratina и Lytoceratina. Вполне возможно, что часть анаптихов может быть на самом деле ринхаптихами, кальцитовые элементы которых не сохранились по тем или иным причинам. Аптихи аммонитов (принадлежавшие, по всей видимости, Haploceratoidea: Lissoceratidae или Oppeliidae, и Stephanoceratoidea: Stephanoceratidae, Garantianinae) встречаются в зоне Niortense существенно реже, чем анаптихи, несмотря на явное доминирование раковин аммонитов, имевших аптиховый тип челюстного аппарата. Найдены также две верхние челюсти, вероятнее всего, принадлежавшие аммонитам с аптиховым типом челюстного аппарата, и одна верхняя челюсть колеоидеи (возможно, белемнита сем. Belemnopseidae или Megateuthididae).
Имеющиеся в настоящее время сведения об особенностях эволюции челюстного аппарата различных систематических групп головоногих моллюсков далеки от совершенства, даже фрагментарны. И хотя изученный материал представлен изолированными элементами челюстных аппаратов и нередко посредственной сохранности, мы сочли необходимым опубликование этих находок, в связи с их редкостью.
* * *
В полевых работах принимали участие О. Нагель (Радеберг, Германия), Ш. Гребенштайн (Бодельсхаузен, Германия), М.П. Шерстюков (Ставрополь), О.В. Трофимов (Ростов-на-Дону). Фотографии на сканирующем электронном микроскопе изготовлены при содействии Р.А. Ракитова (ПИН). Работа выполнена при частичной поддержке одного из авторов (А.А. Мироненко) грантом РФФИ № 18-05-01070. Авторы искренне благодарны всем способствовавшим подготовке этой работы.
Список литературы
Безносов Н.В. Байосские и батские отложения Северного Кавказа. М.: Недра, 1967. 179 с. (Тр. ВНИИГаз. Вып. 28/36).
Безносов Н.В., Митта В.В. Каталог аммонитид и ключевые разрезы верхнего байоса–нижнего бата Северного Кавказа // Бюлл. КФ ВНИГНИ. 1998. № 1. С. 1–70.
Затворницкий А.Я. Среднеюрские глины по р. Кубани // Изв. Геол. ком. 1914. Т. 33. Вып. 250. С. 525–558.
Кахадзе И.Р., Зесашвили В.И. Байосская фауна долины р. Кубани и некоторых ее притоков // Тр. Геол. ин-та АН Груз. ССР. Сер. геол. 1956. Т. 9(14). Вып. 2. 55 с.
Мироненко А.А. Первая находка челюстного аппарата верхневолжских аммонитов Kachpurites fulgens (Craspeditidae) // Палеонтол. журн. 2014. № 6. С. 12–17.
Митта В.В. Первые находки аптихов в верхнем бате Русской платформы // Современные проблемы изучения головоногих моллюсков: морфология, систематика, эволюция, экология и биостратиграфия. М.: ПИН РАН, 2009. С. 66–68.
Митта В.В. Род Spiroceras (Spiroceratidae, Ammonoidea) в верхнем байосе Северного Кавказа // Палеонтол. журн. 2017а. № 2. С. 26–34.
Митта В.В. Род Keppleritiana gen. nov. (Stephanoceratidae, Ammonoidea) из верхнего байоса Северного Кавказа // Палеонтол. журн. 2017б. № 3. С. 26–35.
Митта В.В. О некоторых Rarecostites (Parkinsoniidae, Ammonoidea) зоны Parkinsoni верхнего байоса Северного Кавказа // Палеонтол. журн. 2017в. № 5. С. 13–26.
Митта В.В. О первых находках Djanaliparkinsonia (Stephanoceratidae, Ammonoidea) в верхнем байосе Северного Кавказа // Палеонтол. журн. 2018. № 4. С. 29–37.
Митта В.В., Шерстюков М.П. О байосе и бате бассейна р. Большой Зеленчук (Северный Кавказ) // Проблемы палеоэкологии и исторической геоэкологии. Саратов: СГТУ, 2014. С. 74–81.
Митта В.В., Шерстюков М.П. О находках остатков челюстного аппарата аммонитов в средней юре Карачаево-Черкесии // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция, экология и биостратиграфия. М.: ПИН РАН, 2018. С. 80–83.
Рогов М.А. Средне- и позднекелловейские аптихи (Ammonoidea) Русской плиты // Палеонтол. журн. 2004. № 1. С. 26–33.
Рогов М.А., Гуляев Д.Б. О первой находке аптихов у представителей подсемейства Proplanulitinae Buckman (Perisphinctidae, Ammonitida) // Палеонтол. журн. 2003. № 4. С. 45–48.
Dietze V., Hillebrandt A. von, Riccardi A., Schweigert G. Ammonites and stratigraphy of a Lower Bajocian (Middle Jurassic) section in Sierra Chacaico (Neuquén Basin Argentina) // Zitteliana. Ser. A. 2012. Bd 52. P. 119–139.
Dzyuba O.S., Mitta V.V., Sherstyukov M.P. On the first appearance of dicoelitid belemnites in the Caucasian margin of Tethys: new data from the Upper Bajocian–Lower Bathonian of Karachay-Cherkessia, southwest Russia // Bull. Geosci. 2019. V. 94. № 4. P. 409–424.
Engeser T., Keupp H. Phylogeny of the aptychi-possessing Neoammonoidea (Aptychophora nov., Cephalopoda) // Lethaia. 2002. V. 34. P. 79–96.
Fuchs D., Iba Y. The gladiuses in coleoid cephalopods: homology, parallelism, or convergence? // Swiss J. Paleontol. 2015. V. 134. № 2. P. 187–197.
Keupp H., Mitta V.V. Cephalopod jaws from the Middle Jurassic of Central Russia // N. Jb. Geol. Paläontol. Abh. 2013. Bd 270. P. 23–54.
Lehmann U. Aptychen als Kieferelemente der Ammoniten // Paläontol. Z. 1972. Bd 46. S. 34–48.
Lehmann U. Über den Kieferapparat von Ammoniten der Gattung Parkinsonia // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1978. Bd 48. S. 79–84.
Mironenko A., Gulyaev D. Middle Jurassic ammonoid jaws (anaptychi and rhynchaptychi) from Dagestan, North Caucasus, Russia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2018. V. 489. P. 117–128.
Mitta V.V., Keupp H. Ammoniten-Kiefer aus dem Callovium Russlands // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 2004. Bd 88. S. 125–134.
Mitta V.V., Schweigert G. A new morphotype of lower jaw associated with Calliphylloceras (Cephalopoda: Ammonoidea) from the Middle Jurassic of the Northern Caucasus // Paläontol. Z. 2016. V. 90. № 2. P. 293–297.
Mitta V.V., Schweigert G., Sherstyukov M.P., Dietze V. First finds of ammonite aptychi of Leioceras and Bredyia (Hildoceratoidea) in the Aalenian of Northern Caucasus, Russia // Paläontol. Z. 2018. V. 92. № 4. P. 605–615.
Mitta V.V., Sherstyukov M.P. First record of Patrulia (Ammonoidea: Stephanoceratidae) in the Upper Bajocian of the Northern Caucasus, Russia // N. Jb. Geol. Paläontol. Abh. 2018. V. 288/3. P. 251–254.
Morton N. The aptychi of Sonninia (Ammonitina) from the Bajocian of Scotland // Palaeontology. 1973. V. 16. P. 195–203.
Morton N. Bajocian Sonniniidae and other ammonites from Western Scotland // Palaeontology. 1975. V. 18. P. 41–91.
Quilty P.G. Jurassic ammonites from Ellsworth Land, Antarctica // J. Paleontol. 1970. V. 44. № 1. P. 110–116.
Riegraf W., Luterbacher H. Jurassic and Cretaceous rhyncholites (cephalopod jaws) from the North Atlantic Ocean (Deep Sea Drilling Project Leg 1–79) and their European counterparts. Evidence for the uniformity of the Western Tethys // Geol. Rundschau. 1989. V. 78. P. 1141–1163.
Riegraf W., Schmitt-Riegraf C. Fossilium Catalogue. Mandibula fossils ammonitorum et nautilorum (Rhyncholithi et rhynchoteuthes, excl. aptychi et anaptychi). Pars 134. Amsterdam–N.Y.: Kugler Publ., 1995. 219 p.
Saunders W.B., Spinosa C., Teichert C., Banks R.C. The jaw apparatus of recent Nautilus and its palaeontological implications // Palaeontology. 1978. V. 21. P. 129–141.
Tanabe K., Hikida Y., Iba Y. Two coleoid jaws from the Upper Cretaceous of Hokkaido, Japan // J. Paleontol. 2006. V. 80. № 1. P. 138–145.
Tanabe K., Kruta I., Landman N.H. Ammonoid buccal mass and jaw apparatus // Ammonoid Paleobiology: from Anatomy to Ecology / Eds. Klug C., Korn D., De Baets K. et al. Dordrecht: Springer, 2015a. P. 439–494 (Topics in Geobiology. V. 43).
Tanabe K., Misaki A., Ubukata T. Late Cretaceous record of large soft-bodied coleoids based on lower jaw remains from Hokkaido, Japan // Acta Palaeontol. Pol. 2015b. V. 60. № 1. P. 27–38.
Trauth F. Aptychenstudien. III–V // Ann. Naturhist. Mus. Wien. 1930. Bd 44. S. 329–411.
Westermann G.E.G., Riccardi A.C., Lehmann U. A new Anaptychus-like jaw apparatus of Jurassic? Lytoceras from Argentina // N. Jb. Geol. Paläontol. Monatsch. 1999. Bd 1. P. 21–28.
Wetzel W. Faunistische und stratigraphische Untersuchung der Parkinsonienschichten des Teutoburger Waldes bei Bielefeld // Palaeontogr. Ser. A. 1911. Bd 58. S. 139–277.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал