

Палеонтологический журнал, 2020, № 5, стр. 90-101
Ревизия Plioperdix (Aves: Phasianidae) из плио-плейстоцена Украины
Н. В. Зеленков a, *, Л. В. Горобец b
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Национальный научно-природоведческий музей НАН Украины
Киев, Украина
* E-mail: nikita.zelenkov@gmail.com
Поступила в редакцию 12.11.2019
После доработки 18.11.2019
Принята к публикации 18.11.2019
Аннотация
К Plioperdix pontica (Tugarinov, 1940) традиционно на основании сходных размеров относят остатки мелких фазановых из ряда местонахождений плиоцена и плейстоцена Северного Причерноморья и Восточной Европы (Украина, Молдова, Польша, Чехия, Венгрия). Изучение украинских материалов показало, что в действительности они относятся к нескольким таксонам Galliformes. Материалы из местонахождений кучурганского фаунистического комплекса (MN 14) могут быть отнесены к Eurobambusicola turolicus Zelenkov, 2016 и предположительно “Plioperdix” hungarica (Jánossy, 1991). Plioperdix pontica характеризуется морфологическим сходством с современными Coturnix и представлен материалами преимущественно из Одесских катакомб (верхи MN 15) и нескольких местонахождений зоны MN 16 (в том числе, Рембелицы Крулевски 2; Польша). В местонахождениях среднего виллафранка (напр., Котловина; а также Этулия-3, Молдова; MN 17) доминируют остатки другой неназванной формы, надежно отличающейся от P. pontica по большинству скелетных элементов. Предыдущие описания и диагнозы Plioperdix pontica, выполненные на смешанном материале, здесь переработаны. Обсуждаются проблемы таксономии мелких фазановых неогена–плейстоцена Северного Причерноморья и Восточной Европы.
ВВЕДЕНИЕ
Фазановые (семейство Phasianidae отряда Galliformes) – одна из характерных групп птиц в палеонтологической летописи позднего кайнозоя Европы. Несмотря на относительную многочисленность их остатков, эволюционная история фазановых в неогене и четвертичном периоде не может считаться хорошо изученной – главным образом, из-за запутанности таксономии большого числа описанных форм (напр., Göhlich, Mourer-Chauviré, 2005). Не являются исключением и очень мелкие представители семейства, известные из ряда местонахождений Северного Причерноморья. История их изучения начинается с работы А.Я. Тугаринова (1940), описавшего два таксона из нижнего плиоцена Одесских катакомб: ископаемый вид пустынной куропатки Ammoperdix ponticus Tugarinov, 1940 и похожий на современного перепела Pliogallus coturnoides Tugarinov, 1940. Вскоре П.В. Серебровский (1941) отнес хорошо сохранившийся коракоид из того же местонахождения к P. coturnoides и указал на возможную синонимию Pliogallus и современного рода перепелов Coturnix. Позднее М. Кретцой (Kretzoi, 1955) отметил, что родовое название Pliogallus Tugarinov, 1940 преоккупировано и предложил новое – Plioperdix Kretzoi, 1955. В каталоге ископаемых птиц П. Бродкорба (Brodkorb, 1964) фигурируют оба таксона: Ammoperdix ponticus и Plioperdix coturnoides. Бродкорб включил в род Plioperdix еще три мелких вида фазановых из среднего миоцена Франции, которые в настоящее время относятся к ископаемым родам Palaeortyx и Palaeocryptonyx (Göhlich, Mourer-Chauviré, 2005).
М.А. Воинственский (1967) определил “Pliogallus coturnoides” из ряда плиоценовых и раннеплейстоценовых (по современным представлениям) местонахождений Украины (Кучурган, Одесские катакомбы, Крыжановка, Соколовский карьер и др.), однако не описал их морфологию и не указал коллекционные номера. Морфологическая ревизия голотипа ponticus и описание переданных в ПИН материалов (не тех, что были упомянуты Воинственским) из нескольких местонахождений юга Украины (исключая типовое!) и Молдовы были проведены Е.Н. Курочкиным (1985; Bocheński, Kurochkin, 1987), установившим, что Ammoperdix ponticus и Plioperdix coturnoides представляют один вид, который должен называться Plioperdix ponticus (Tugarinov, 1940). Курочкин (1985) отметил некоторое сходство P. ponticus и современных Coturnix, но посчитал, что у этой ископаемой формы нет близких родственников в современной фауне; он также отнес к Plioperdix ponticus остатки из верхнего плиоцена Забайкалья; позднее P. ponticus был найден и в верхнем плиоцене северной Монголии (Зеленков, Курочкин, 2009).
Следующий этап эволюции представлений о родственных связях, распространении, разнообразии и таксономии Plioperdix связан с работами Й. Мликовского (Mlíkovský, 1995, 2002). Первоначально он (Mlíkovský, 1995; см. также Bocheński et al., 2012) отнес к P. ponticus мелких фазановых из нижнего и верхнего плиоцена (MN 15, 16) Польши, до этого обозначавшихся как Francolinus minor Jánossy, 1974, и также определил P. ponticus по материалам из нижнего плейстоцена (MNQ20-21; MQ1b) местонахождения Странска Скала в Чехии (Mlíkovský, 1995). Синонимизация Plioperdix ponticus и Francolinus minor была основана на морфологическом сходстве обеих форм с Coturnix, при этом непосредственное сравнение материалов, по-видимому, не проводилось. В род Plioperdix Мликовский отнес и другие, заметно более крупные ископаемые виды Francolinus (Mlíkovský, 1995). Позднее он (Mlíkovský, 2002) свел P. ponticus в младшие синонимы Palaeocryptonyx donnezani Depéret, 1892 из плиоцен–плейстоцена Западной Европы, которого, в свою очередь, включил в современный род Alectoris. Необоснованность этого и других таксономических решений Мликовского не раз отмечалась в литературе (Mourer-Chauviré, 2004; Зеленков, Курочкин, 2009, 2015).
Позднее к роду Plioperdix были отнесены еще два вида, существенно расширившие его географию и возраст. Из нижнего плейстоцена (MN 17) Марокко был описан P. africana Mourer-Chauviré et Geraads, 2010, заметно отличающийся от P. ponticus морфологически и по размерам (Mourer-Chauviré, Geraads, 2010). Н.В. Зеленков и А.В. Пантелеев (Zelenkov, Panteleyev, 2015; Зеленков, 2016) переместили в род Plioperdix еще один вид – Palaeocryptonyx hungaricus Jánossy, 1991 из верхнего миоцена (MN 13) Венгрии, к которому они также отнесли ряд материалов из верхнего миоцена (MN 11–12) местонахождения Морская-2 в Приазовье (Zelenkov, Panteleyev, 2015). Эти же авторы (см. также Зеленков, Курочкин, 2015) изменили видовой эпитет на pontica для грамматического согласования с родовым названием по аналогии с названием женского рода Perdix (Dickinson, Remsen, 2013).
Работая с материалами из Центральной Азии и Восточной Европы, один из авторов настоящего сообщения (Н.В. Зеленков) неоднократно сталкивался с морфологической неоднородностью материалов, традиционно относимых к P. pontica (см. Зеленков, 2015). Эта морфологическая неоднородность отчетливо контрастирует с общей слабо выраженной морфологической изменчивостью у фазановых и обуславливает необходимость таксономической ревизии материалов по Plioperdix pontica, чему, в первую очередь, и посвящена настоящая статья. В настоящей работе описаны материалы из ряда неогеновых местонахождений преимущественно Украины, хранящиеся в Национальном научно-природоведческом музее в Киеве (NMNHU) и Палеонтологическом ин-те им. А.А. Борисяка РАН в Москве (ПИН РАН). Поскольку в предыдущих работах (Курочкин, 1985) морфологическая характеристика P. pontica была в значительной степени основана на материалах из местонахождений Этулия (Молдова) и Береговая (Россия, Забайкалье), эти материалы также учитываются в этой работе. Изучение ранее описанных и новых материалов, традиционно определявшихся как P. pontica, позволило выявить присутствие в позднем кайнозое Северного Причерноморья нескольких таксонов мелких фазановых, относящихся к Plioperdix и еще двум–трем родам. Таким образом, предыдущие морфологические характеристики P. pontica оказываются основанными на смешанных материалах по нескольким таксонам и поэтому здесь исправлены.
Возраст местонахождений приведен в соответствии с рядом источников (Nesin, Nadachowski, 2001; Тесаков, 2004; Tesakov et al., 2007). Исследование выполнено за счет гранта Российского научного фонда (проект № 18-74-10081). Авторы благодарны А.Б. Савинецкому и Н.В. Мартыновичу за замечания по рукописи статьи.
МОРФОЛОГИЧЕСКАЯ ИДЕНТИФИКАЦИЯ PLIOPERDIX PONTICA
Тарсометатарсус. Голотипом Ammoperdix ponticus Tugarinov, 1940 послужил практически целый тарсометатарсус № 6489 (рис. 1, б) без обозначения принадлежности к какому-то учреждению (Тугаринов, 1940). Затем он был обнаружен Е.Н. Курочкиным в Зоологическом ин-те AH СССР в Ленинграде и описан (Курочкин, 1985) как принадлежащий коллекции Ин-та зоологии АН УССР (Киев), куда он был возвращен в 2019 г. Характерные черты, надежно отличающие тарсометатарсус pontica от Ammoperdix и указывающие на близость этой формы к Coturnix – сильно сближенные trochleae metatarsorum (в особенности, приближенный к продольной оси кости trochlea metatarsi II) и сдвинутый плантарно trochlea metatarsi IV (Курочкин, 1985). Каких-либо значительных структурных отличий в строении дистального эпифиза тарсометатарсуса и его стержня от современных Coturnix выявить не удается; в то же время ряд мелких морфологических особенностей прослеживается (см. ниже).
Рис. 1.
Тарсометатарсусы мелких фазановых из плиоцена и нижнего плейстоцена Северного Причерноморья: а, б, з, к – Plioperdix pontica (Tugarinov, 1940): а, з – экз. NMNHU-P Av-674; б, к – голотип NMNHU-P Av-6489; местонахождение Одесские катакомбы; нижний плиоцен; в, ж, и, л – Phasianidae indet. 2: в, и, л – экз. ПИН, № 2614/64 (отражен); местонахождение Этулия-3, Молдова; нижний плейстоцен; ж – экз. NMNHU-P Av-708; местонахождение Котловина, Украина; предположительно нижний плейстоцен; г, е, м – Coturnix coturnix, современный; д – Eurobambusicola turolicus Zelenkov, 2016, экз. NMNHU-P Av-486; местонахождение Кучурган, Украина; нижний плиоцен (отражен); а–д – с дорсальной стороны; е–и – с проксимальной стороны; к–м – с дистальной стороны. Обозначения: cl1 – выступающий дорсально cotyla lateralis (апоморфия Coturnix); cl2 – не выступающий дорсально cotyla lateralis; iil1 – узкая incisura intertrochlearis lateralis; iil2 – широкая incisura intertrochlearis lateralis. Длина масштабной линейки – 0.5 см.
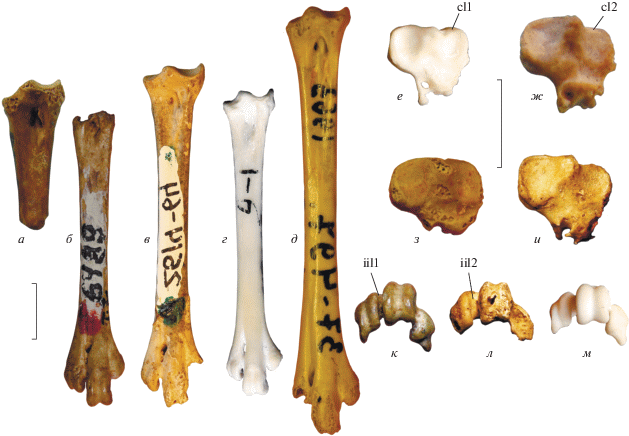
Все признаки проксимального эпифиза тарсометатарсуса P. pontica из описания Курочкина (1985) и последующих работ, опирающихся на эту публикацию (Bocheński, Kurochkin, 1987; Зеленков, Курочкин, 2009, 2015), основаны на цевках от других таксонов. Судя по приведенному Курочкиным (1985) описанию, признаки проксимальной цевки были основаны на экз. ПИН, № 2614/64 из местонахождения Этулия-3 (см. ниже) – у этого экземпляра частично сохранился дистальный эпифиз, что позволяет сравнивать его с голотипом. Но экз. ПИН, № 2614/64 отличается от голотипа P. pontica большей длиной (при сходной толщине кости расстояние между дистальным питательным отверстием и дистальным краем гипотарсуса у экз. из Этулии больше), не сдвинутой плантарно trochlea metatarsi IV относительно латероплантарного угла trochlea metatarsi III, несколько более широкой incisura intertrochlearis lateralis, заметно меньше опущенной плантарно trochlea metatarsi II, а также тем, что канал гипотарсуса выходит в центральной части гипотарсуса (как у Palaeocryptonyx edwardsi из среднего миоцена Франции – наши данные). У голотипа P. pontica канал открывается на медиальной стороне гипотарсуса, а trochlea metatarsi IV сдвинута плантарно – как у современных Coturnix. Кроме того, у экз. № 2614/64 cotyla lateralis не выступает дорсально относительно eminentia intercotylaris (как у Coturnix), а fossa infracotylaris dorsalis имеет пологий медиальный край (у Coturnix ямка доходит до медиального края кости и имеет крутую стенку). При виде с дорсальной стороны cotyla lateralis располагается примерно на одном уровне с cotyla medialis (формирует неглубокую вырезку в проксимальном профиле кости), в то время как у Coturnix s.l. cotyla lateralis сдвинут дистально, вследствие чего формирует глубокую вырезку в проксимальном крае кости. Таким образом, экз. № 2614/64 из Этулии не показывает продвинутой морфологии Coturnix s.l. и не может быть отнесен к P. pontica, поскольку отличается от голотипа рядом существенных морфологических деталей.
В то же время, несколько характерных признаков Coturnix имеет проксимальный фрагмент тарсометатарсуса экз. NMNHU-P Av-674 из Одесских катакомб: глубокую и медиально расширенную fossa infracotylaris dorsalis с крутой медиальной стенкой, выступающий дорсально cotyla lateralis, формирующий глубокую вырезку в проксимальном крае кости, а также сильно сдвинутый плантарно канал гипотарсуса. Кроме того, для этого экземпляра характерен вытянутый дистально гипотарсус, как у современных Coturnix (но в отличие, в частности, от Ammoperdix и Chauvireria). Сходство с современными Coturnix позволяет относить этот проксимальный фрагмент к P. pontica и свидетельствует о том, что проксимальный эпифиз цевки у этого ископаемого вида также был устроен сходным образом с таковым у современных Coturnix.
Дистальный конец тарсометатарсуса из местонахождения Котловина (экз. NMNHU-P Av707) также, по-видимому, принадлежит P. pontica, поскольку имеет прижатый к trochlea metatarsi III и нескольку опущенный плантарно trochlea metatarsi IV. Так же как и у голотипа, у этого экземпляра прослеживается отчетливая вырезка в медиальном крае профиля trochlea metatarsi III при виде с дистальной стороны. Trochlea metatarsi II умеренно опущен плантарно – отчасти сходно с состоянием у голотипа P. pontica, но выражено слабее, чем у “Francolinus” minor. По сравнению с голотипом Plioperdix pontica trochlea metatarsi III выглядит дорсоплантарно менее высоким при виде с дистальной стороны, но эта особенность может объясняться индивидуальной или временной изменчивостью.
Практически полный правый тарсометатарсус (экз. ПИН, № 2614/146) из нижнего плиоцена местонахождения Войничево (Украина; Одесская обл.; MN 14) демонстрирует характерные для Coturnix s.l. признаки: наличие глубокой смещенной медиально fossa infracotylaris dorsalis, удлиненного hypotarsus, некоторым образом скрученного trochlea metatarsi III и узкого в целом дистального эпифиза с прижатыми trocheae metatarsorum (особенно trochlea metatarsi II). При этом экз. ПИН, № 2614/146 отличается от голотипа P. pontica медиолатерально широким trochlea metatarsi III и сдвинутым дорсально trochlea metatarsi IV при виде с дистальной стороны.
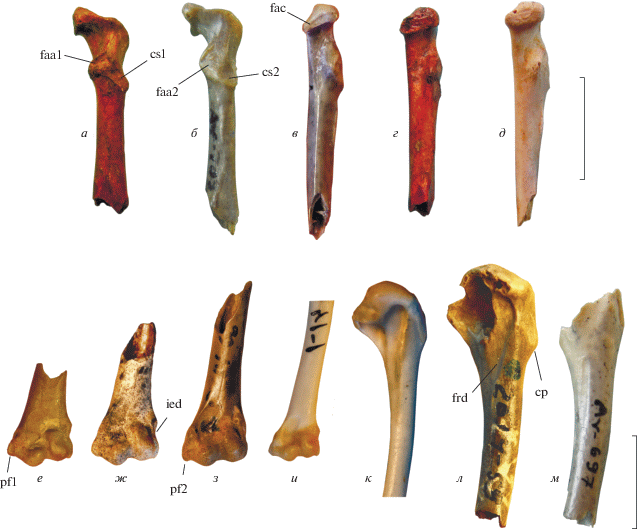
Коракоид. Тугаринов (1940) отнес к Pliogallus coturnoides фрагментарный правый коракоид (“№ 6478”; ныне экз. NMNHU-P Av-674), но не привел его изображения или описания. Позднее Серебровский (1941) опубликовал практически полный коракоид из типового местонахождения и отнес его к Pliogallus coturnoides. Экземпляр, описанный Серебровским, по-видимому, не сохранился, так как в коллекции NMNHU имеются только фрагментарные экземпляры. Курочкин (1985; Bocheński, Kurochkin, 1987) при подготовке описания Plioperdix pontica использовал только коракоиды из Забайкалья (местонахождение Береговая), которые, по-видимому, не были сравнены с материалами из Одесских катакомб. Описание коракоидов, выполненное Курочкиным (1985), было позднее использовано Зеленковым и Курочкиным (2009) для отнесения дополнительных материалов из местонахождения Шамар в Монголии к P. pontica.
Однако коракоиды из Одесских катакомб могут быть достоверно отличимы от коракоидов, описанных как P. pontica из ряда других местонахождений. У коракоидов из Одесских катакомб вентромедиальный край стержня краниальнее уровня processus procoracoideus изгибается несколько вентрально – таким образом, вентральная поверхность processus acrocoracoideus оказывается ориентированной не строго параллельно плоскости грудинного расширения, но под некоторым углом к ней (аналогичное состояние встречается у некоторых утиных – например, Anserini). В результате такого поворота краниальной части кости при виде с вентральной стороны медиальный край стержня оказывается в значительной степени параллелен длинной оси кости и сильнее приближен к labrum glenoidale, чем у многих других мелких фазановых. Отчасти сходная морфология опять-таки характеризует современных Coturnix – значительное сходство коракоидов из Одесских катакомб и современных перепелов уже отмечалось Серебровским (1941). У коракоидов из Забайкалья и Шамара подобный поворот краниальной части коракоида не выражен. Примечательно, что коракоид из более молодого местонахождения Попово-1 (экз. NMNHU Av 684) морфологически крайне сходен с таковым из Одесских катакомб – это свидетельствует о стабильности морфологического типа во времени и позволяет диагностировать P. pontica на основании данного элемента скелета.
Коракоиды из Одесских катакомб также сходны с таковыми современных Coturnix морфологией processus procoracoideus и cotyla scapularis. У Coturnix при виде с дорсальной стороны cotyla scapularis сужен краниокаудально за счет того, что ориентирован несколько более краниально, чем дорсально. При плезиоморфном для фазановых состоянии cotyla ориентирован практически строго дорсально и поэтому при виде с дорсальной стороны выглядит заметно более расширенным краниокаудально. Кроме того, у фазановых на дорсальной поверхности processus procoracoideus располагается обширная сочленовная поверхность для соединения с акромионом лопатки (pис. 2, faa1, faa2) – у Coturnix и коракоидов из Одесских катакомб эта поверхность заметно редуцирована и занимает только краниальную половину дорсальной поверхности processus procoracoideus. По морфологии данной (лопаточной) области коракоида экземпляры из местонахождения Котловина заметно отличаются от Coturnix и коракоидов из Одесских катакомб, которые мы относим к Plioperdix. Помимо этого, facies articularis clavicularis при виде с медиальной стороны заметно суживается у коракоидов из Котловины, в отличие от Plioperdix из Одесских катакомб и Попово. Также, в отличие от Plioperdix, при виде с медиальной стороны весь extremitas omalis у коракоидов из Котловины несколько отклоняется вентрально, а при виде с дорсальной стороны впадина между cotyla scapulais и выступающей латерально частью facies articularis humeralis выражена очень слабо.
Плечевая кость. Плечевые кости не упоминались в первоначальных публикациях по Plioperdix и впервые были перечислены (но также не описаны) Воинственским (1967) из Одесских катакомб и, вероятно, плиоценовых слоев Соколовского карьера (Кривой Рог, Украина; основная фауна этого местонахождения относится к миоцену). Курочкин (1985) использовал для характеризации морфологии плечевой кости P. pontica экземпляры из Этулии-3 (экз. ПИН, №№ 2614/51, 59, 62, 66) и Забайкалья. Впоследствии эти данные были использованы в работах Зеленкова (Зеленков, Курочкин, 2009, 2015; Zelenkov, Panteleyev, 2015; Зеленков, 2016). Однако, по результатам настоящего обзора в Этулии доминируют остатки другой фазановой птицы (не P. pontica); судя по коракоидам, мелкая фазановая птица из плиоцена Забайкалья также не может быть отнесена к P. pontica (см. выше). Таким образом, морфология плечевой кости P. pontica до сих пор остается неопределенной.
Среди материалов из Одесских катакомб в коллекции NMNHU имеются два дистальных фрагмента плечевой кости (экз. NMNHU Av 669, 5242, 5243), морфологически схожие с плечевыми костями современных Coturnix – это позволяет относить их к P. pontica. На экз. NMNHU Av 669 хорошо прослеживается характерная для Coturnix выпрямленность стержня. Кроме того, оба экземпляра характеризуются сильно выраженной вырезкой в дорсальном крае epicondylus dorsalis (при виде с краниальной стороны), при этом сам epicondylus развит умеренно (при виде с краниальной стороны его ширина составляет около трети ширины condylus dorsalis). У экз. NMNHU Av 669 также прослеживаются сильно выдающийся краниально tuberculum supracondylare ventrale и слабо выставленный дистально processus flexorius при виде с краниальной стороны. Дистальный эпифиз в целом существенно дорсовентрально сжат (узкий). Все эти признаки характеризуют и современных Coturnix. Плечевая кость из Соколовского карьера (NMNHU-P Av-88), а также ряд экземпляров из местонахождения Котловина (см. ниже) характеризуются той же морфологией.
Отмеченный выше комплекс морфологических признаков характерен для дистального фрагмента плечевой кости из Этулии (экз. ПИН, № 2614/51), в то время как у другого образца из Этулии (экз. ПИН, № 2614/66) вырезка в epicondylus dorsalis мелкая, сам epicondylus тонкий (его толщина при виде с краниальной стороны составляет около ¼ толщины condylus dorsalis), processus flexorius заметно менее выдвинут дистально, и весь дистальный эпифиз в целом кажется несколько расширенным по сравнению с прилегающей дистальной частью стержня. Отмеченные отличия по всем прослеживаемым признакам свидетельствуют о том, что экз. ПИН, № 2614/66, по-видимому, принадлежал другому таксону мелких фазановых птиц. У экз. ПИН, № 2614/145 из нижнего плиоцена местонахождения Войничево processus flexorius выдвинут дистально – таким образом, этот экземпляр тоже не может быть отнесен к P. pontica. Присутствие двух сходных по размеру фазановых птиц в местонахождении Этулия не позволяет с уверенностью отнести к P. pontica проксимальную плечевую кость из этого же местонахождения (экз. ПИН, № 2614/59), для которой все же характерны некоторые черты, сближающие ее с Coturnix: отчетливая выставленность дорсально дистального угла crista pectoralis (см. Zelenkov, Panteleyev, 2015), наличие четко огороженного желоба, простирающегося дистально от fossa tricipitalis dorsalis.
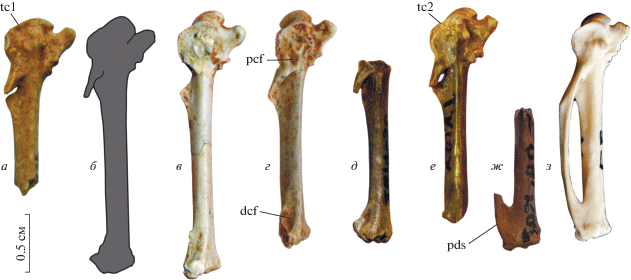
Карпометакарпусы из Одесских катакомб также морфологически сходны с таковыми современных Coturnix – уже Тугаринов (1940), описывая их, не отметил никаких структурных отличий, кроме большей длины. Однако Курочкин (1985, с. 100) в своем сравнительном анализе не использовал материалы из Одесских катакомб, но основывался на пряжках из других местонахождений – в частности, из Котловины, которые имеют отличную морфологию и должны представлять другой таксон. Выделенный им ряд отличий Plioperdix от современного Coturnix основан именно на этих материалах из Котловины. В то же время, для карпометакарпусов из Одесских катакомб Тугаринов (1940) отмечал наличие маленького processus intermetacarpalis, выпрямленной metacarpalia minor и, как следствие, узкого spatium intercarpale, что является характерными чертами современных Coturnix. Маленький отросток, действительно, сближает карпометакарпусы из Одесских катакомб с Coturnix. Признаки metacarpalia minor напрямую не прослеживаются у двух описанных Тугариновым экземпляров (современные номера: NMNHU-P Av-340, 341), поскольку у них малая метакарпалия, в целом, не сохранилась, однако они легко реконструируются по углу ее отхождения от большой метакарпалии. Но на экз. NMNHU-P Av-340 (голотип Pliogallus coturnoides) четко прослеживается другая апоморфия Coturnix – характерное строение вентральной части trochlea carpalis. У всех фазановых вентральный блок trochlea carpalis разделен вырезкой на две части, при этом каудальная (дистальная) часть у подавляющего большинства заметно сдвинута дистально относительно проксимальной. По этому признаку род Coturnix заметно отличается от своих ближайших родственников (в том числе, от филогенетически близкого Ammoperdix): у Coturnix вырезка в вентральном крае trochlea carpalis практически не выражена и каудальная (дистальная) часть сдвинута проксимально, так что весь вентральный блок имеет округлые очертания – как, например, у Anatidae. Эта черта сходства с Coturnix, хорошо выраженная на экз. NMNHU-P Av-340, подтверждает отнесение этого экземпляра P. pontica и синонимию этого вида с Pliogallus coturnoides. Еще одна характерная черта Coturnix, прослеживаемая на карпометакарпусах из Одесских катакомб – очень слабая изогнутость metacarpalia major при виде с краниальной стороны (кость выглядит практически прямой).
В целом устройство вентральной части trochlea carpalis, отмеченное выше для Coturnix, характерно и для примитивных фазановых Rollulinae, а также еще более примитивного семейства Odontophorinae. Однако Odontophorinae и Rollulinae отличаются от современных Coturnix и экз. NMNHU-P Av-340 морфологией малой метакарпалии, которая у них сильно изогнута, и поэтому в своем основании отходит от большой метакарпалии под заметно большим углом. Кроме того, плоскость малой метакарпалии (в ее проксимальной части) у Coturnix ориентирована преимущественно каудально, в то время как у Odontophorinae и Rollulinae она развернута вентрокаудально (поэтому метакарпалия заметно более широкая при виде с вентральной стороны). Этот признак представляет дополнительную черту сходства NMNHU-P Av-340 с родом Coturnix.
Курочкин (1985) отмечал, что Plioperdix отличается от Coturnix наличием двух ямок на вентральной поверхности малой метакарпалии: в ее проксимальной и дистальной частях. Примечательно, что эти ямки не прослеживаются на материалах из Одесских катакомб, что опять-таки подтверждает близость Plioperdix к Coturnix. Эти ямки есть у карпометакарпусов из Котловины, которые мы относим к другому таксону. В дистальной части карпометакарпуса характерными чертами Coturnix являются очень слабо выступающая дистально суставная поверхность для малого пальца крыла и длинный симфиз. Важно, что у обоих экземпляров из Одесских катакомб каудальный край симфиза ориентирован параллельно длинной оси кости – в точности, как у Coturnix, что указывает на то, что малая метакарпалия у Plioperdix была ориентирована продольно и была не выгнута каудально [что предполагал Тугаринов (1940) на проксимальных фрагментах]. Экз. ПИН, № 2614/137 из местонахождения Котловина, отнесенный Курочкиным (1985) к P. pontica, отличается от материалов из Одесских катакомб и современного Coturnix тем, что дистальная часть вентрального блока trochlea carpalis сдвинута дистально (напоминает состояние у современного Perdix) и, кроме того, присутствует отмеченная ямка в основании вентральной поверхности малой метакарпалии. Такое же строение имеет и экз. NMNHU-P Av-686 из Котловины. Metacarpalia major у карпометакарпусов из Котловины заметно изогнута дугой при виде с краниальной стороны.
Особая отличительная черта карпометакарпуса P. pontica, отмеченная еще Тугариновым (1940) – удлиненные пропорции кости в сравнении с современными Coturnix. В настоящее время у экз. NMNHU-P Av-340 (голотип Pliogallus coturnoides) дистальная часть обломана, но иллюстрация в первоописании уверенно свидетельствует о пропорциях, отличных от таковых современных Coturnix. Важно заметить, что такие же удлиненные пропорции карпометакарпуса при сходном размере проксимального эпифиза обнаружены нами у экз. без номера в колл. Ин-та систематики и эволюции животных в Кракове (Польша), первоначально включенного в материалы по “Francolinus” minor (Jánossy, 1974). Этот экз. происходит из местонахождения Рембелицы Крулевски 2 (Польша, 3.4–3.1 млн л.; Тесаков, 2004) и отличается от голотипа “Francolinus” minor из геологически более молодого местонахождения Рембелицы Крулевски 1 (3.15–2.7 млн л.; Тесаков, 2004) не только длиной, но и рядом морфологических деталей. Экз. из Рембелицы Крулевски 2 сходен с современным Coturnix и указанными выше материалами из Одесских катакомб (3.8–3.7 млн л.; Tesakov et al., 2007) отсутствием вырезки в trochlea carpalis (см. выше) и ямок на вентральной поверхности кости (дистальная неявно выражена). Таким образом, наличие сходного по пропорциям и морфологии карпометакарпуса в достаточно удаленных местонахождениях свидетельствует о стабильности морфотипа, характеризующего P. pontica. В то же время, голотип “Francolinus” minor отчетливо демонстрирует морфологию, соответствующую дифференциации на родовом уровне в современном разнообразии – таким образом, этот вид не только не может считаться младшим синонимом pontica (Mlíkovský, 1995 и последующие работы), но и должен быть исключен из рода Plioperdix.
С учетом полученных данных, свидетельствующих о присутствии в плиоцене–нижнем плейстоцене Северного Причерноморья нескольких форм мелких фазановых, ниже приведены уточненные морфологические характеристики P. pontica.
СИСТЕМАТИЧЕСКИЙ РАЗДЕЛ
ОТРЯД GALLIFORMES
СЕМЕЙСТВО PHASIANIDAE HORSFIELD, 1821
Триба Coturnicini Reichenbach, 1848
Род Plioperdix Kretzoi, 1955
Ammoperdix: Тугаринов, 1940, с. 311; Brodkorb, 1964, с. 318.
Pliogallus: Тугаринов, 1940, с. 312.
Plioperdix: Kretzoi, 1955, с. 367; Brodkorb, 1964, с. 317; Курочкин, 1985, с. 97; Bocheński, Kurochkin, 1987, с. 83; Зеленков, Курочкин, 2009, с. 80; Mourer-Сhauviré, Geraads, 2010, с. 168; Zelenkov, Panteleyev, 2015, с. 517; Зеленков, Курочкин, 2015, с. 184; Зеленков, 2016, с. 89.
Alectoris: Mlíkovský, 2002, с. 157 (part.).
Типовой вид Ammoperdix ponticus Tugarinov, 1940.
Диагноз. Мелкие фазановые птицы, морфологически очень сходные с современными Coturnix. В коракоиде краниальная часть стержня (краниальнее processus procoracoideus) не субпараллельна плоскости грудинной части, поскольку вентромедиальный край стержня отгибается вентрально; при виде с вентральной стороны медиальный край стержня коракоида (краниальнее processus procoracoideus) субпараллелен длинной оси кости и приближен к labrum glenoidale; impressio bicipitalis умеренно выдается медиально относительно медиального края processus acrocoracoideus; processus procoracoideus относительно робустный; cotyla scapularis ориентирован несколько более краниально, чем дорсально; сочленовная поверхность для соединения с акромионом лопатки, расположенная медиальнее cotyla scapularis, сужена краниокауадально. В карпометакарпусе метакарпалии удлинены: отношение длины кости к длине проксимального эпифиза (до точки бифуркации метакарпалий) составляет около 4.4. В тарсометатарсусе при виде с дистальной стороны trochlea metatarsi II сильно опущен плантарно, так что его большая часть выдается плантарнее, чем плантарная кромка trochlea metatarsi III; латеральная часть trochlea metatarsi IV также сильно выдается плантарно; при виде с дорсальной стороны дистальное окончание trochlea metatarsi IV находится на уровне середины trochlea metatarsi III.
Видовой состав. Кроме типового вида, в состав рода были включены P. hungarica (Jánossy, 1991) из позднего миоцена (MN 11-13) Венгрии и юга европейский части России, а также P. africana Mourer-Chauviré et Geraads, 2010 из раннего плейстоцена (MN 17) Марокко, однако принадлежность обоих видов к Plioperdix требует подтверждения, в связи с полученными в этой работе данными о большем разнообразии мелких фазановых позднего неогена Восточной Европы.
Сравнение. Морфологически очень близок современным Coturnix s.l. (включая Excalfactoria), но отличается рядом структурных деталей. Коракоид отличается от такового Coturnix s.l. менее выдающимся медиально impressio bicipitalis, более выраженным processus procoracoideus, а также общей робустностью. Отличий в строении дистального эпифиза плечевой кости не выявлено. Карпометакарпус имеет пропорционально более удлиненные, чем у Coturnix, метакарпалии. В строении дистального эпифиза тарсометатарсуса отличается от Coturnix тем, что trochlea metatarsi II сильнее опущен плантарно. Trochlea metatarsi III уже, чем у Coturnix. Также trochlea metatarsi IV несколько сильнее сдвинут плантарно по сравнению с Coturnix: медиальный угол плантарного края trochlea metatarsi IV находится на уровне плантарного края trochlea metatarsi III, а латеральный угол (латеральный крыловидный отросток) выступает заметно плантарнее и в целом сильнее развит, чем у Coturnix. Еще одно отличие от Coturnix заключается в том, что trochlea metatarsi IV длиннее: при виде с дорсальной стороны он доходит до середины trochlea metatarsi III, в то время как у Coturnix он укорочен и приближается к первой трети trochlea metatarsi III (это также заметно при виде с плантарной стороны).
Замечания. Приведенный Курочкиным (1985) диагноз (“описание”) здесь полностью переработан с учетом представлений о том, какие именно ископаемые остатки следует относить к Plioperdix. В диагнозе отмечены черты, позволяющие отличать Plioperdix от морфологически наиболее близкого рода Coturnix; отличия Plioperdix и Coturnix от остальных фазановых даны выше.
Plioperdix pontica (Tugarinov, 1940)
Ammoperdix ponticus: Тугаринов, 1940, с. 311, рис. 2; Brodkorb, 1964, с. 318.
Рис. 2.
Коракоиды (а–д) и плечевые кости (е–м) мелких фазановых из плиоцена и нижнего плейстоцена Северного Причерноморья: а, г–ж – Plioperdix pontica (Tugarinov, 1940): а, г – экз. NMNHU-P Av-666; местонахождение Одесские катакомбы; нижний плиоцен; д – экз. NMNHU-P Av-684; местонахождение Попово-1, Украина; верхний плиоцен; е – экз. NMNHU, № 5243; местонахождение Одесские катакомбы; нижний плиоцен; ж – экз. ПИН, № 2614/51; местонахождение Этулия, Молдова; предположительно нижний плиоцен (см. текст); б, в, з, м – Phasianidae indet. 2: б, в – экз. NMNHU-P Av-709; местонахождение Котловина; предположительно, нижний плейстоцен (см. текст); з – экз. ПИН, № 2614/51; местонахождение Этулия; предположительно, нижний плиоцен; л – Phasianidae indet. (?Plioperdix pontica), экз. ПИН, № 2614/59, проксимальный фрагмент правой плечевой кости; местонахождение Этулия, Молдова; предположительно, нижний плиоцен (см. текст); м – экз. NMNHU-P Av-697; местонахождение Котловина; предположительно, нижний плейстоцен; и, к – Coturnix coturnix, современный. Обозначения: cp – выставленный дорсально угол crista deltopectoralis; cs1 – краниально ориентированный cotyla scapularis; cs2 – дорсально ориентированный cotyla scapularis; faa1 – редуцированная сочленовная поверхность для соединения с акромионом лопатки; faa2 – обширная сочленовная поверхность для соединения с акромионом лопатки; fac – зауженная вентрально facies articularis clavicularis; ftd – желоб, представляющий собой продолжение fossa tricipitalis dorsalis; ied – вырезка в epicondylus dorsalis; pf1 – не выдвинутый дистально processus flexorius; pf2 – выдвинутый дистально processus flexorius. а, б – вид с дорсальной стороны; в–д – вид с медиальной стороны; е–и – вид с краниальной стороны; к–м – вид с каудальной стороны. Длина масштабной линейки – 1 см.
Pliogallus coturnoides: Тугаринов, 1940, с. 312, рис. 3; Воинственский, 1967, с. 18 (part.).
Рис. 3.
Карпометакарпусы мелких фазановых из плиоцена и нижнего плейстоцена Северного Причерноморья с вентральной стороны: а–в, ж – Plioperdix pontica (Tugarinov, 1940): а, б – экз. NMNHU-P Av-340, фотография сохранившейся части (а) и прорисовка (б) по изображению, приведенному в первоописании (Тугаринов, 1940); местонахождение Одесские катакомбы; верхи нижнего плиоцена; в – экз. б/н в колл. Ин-та систематики и эволюции животных (Краков); местонахождение Рембелицы Крулевски-2, Польша; низы верхнего плиоцена; ж – экз. NMNHU-P Av-671, дистальный фрагмент (отражен); местонахождение Одесские катакомбы; верхи нижнего плиоцена; г – “Francolinus” minor Jánossy, 1974, голотип, Ин-т систематики и эволюции животных (Краков), № AF 61; местонахождение Рембелицы Крулевски-1, Польша; верхний плиоцен; д, е – Phasianidae indet. 2: д – экз. ПИН, № 2614/50; местонахождение Этулия-3, Молдова; нижний плейстоцен; е – экз. NMNHU-P Av-716; местонахождение Котловина, Украина; предположительно нижний плейстоцен; з – Coturnix coturnix, современный. Обозначения: dcf – ямка в дистальной части metacapalia minor; pcf – ямка в проксимальной части metacapalia minor; pds – дистальный симфиз с каудальным краем, субпараллельным длинной оси кости; tc1 – trochlea carpalis без выраженной вырезки и без сдвинутой дистально каудальной части; tc2 – trochlea carpalis с вырезкой, каудальная часть сдвинута дистально.
Plioperdix coturnoides: Brodkorb, 1964, с. 317; Bocheński, 1997, с. 310.
Plioperdix ponticus (part.): Курочкин, 1985, с. 100, табл. XV , фиг. 1–12 ; Bocheński, 1997, с. 310; Зеленков, Курочкин, 2009, с. 83, табл. IX , фиг. 7–16 .
Alectoris donnezani (part.): Mlíkovský, 2002, с. 159.
Plioperdix pontica (part.): Зеленков, Курочкин, 2015, с. 185, табл. XIX , фиг. 9–22 .
Голотип – NMNHU-P 6489, правый тарсометатарсус без наиболее проксимальной части; Украина, Одесская обл., местонахождение Одесские катакомбы; плиоцен (молдавский фаунистический комплекс; верхи биозоны MN 15).
Описание (рис. 1, а, б, з, к; рис. 2, а, г–ж, л; рис. 3, а–в, ж). Cм. раздел “Морфологическая идентификация Plioperdix pontica”.
Сравнение. Отличается от P. africana меньшими размерами, мелким processus intermetacarpalis, не загнутым проксимально processus extensorius и некрупным processus pisiformis карпометакарпуса; от P. hungarica – удлиненным карпометакарпусом и сильно прижатым к длинной оси тарсометатарсуса trochlea metatarsi IV.
Замечания. Проведенная в настоящей работе ревизия материалов из местонахождений Украины позволила исключить из P. pontica кости, относящиеся к другим таксонам. В то же время, к P. pontica мы также относим фрагмент плечевой кости из местонахождения Обуховка в Ростовской обл. (экз. NMNHU-P Av-683), характеризующийся слабо выдающимся дистально processus flexorius. Дистальные фрагменты тибиотарсусов из местонахождения Котловина (экз. NMNHU-P Av-704-706) предварительно отнесены к P. pontica, поскольку характеризуются неглубокой incisura intercondylaris (как у Coturnix), однако аналогичных костей из Одесских катакомб не известно, поэтому морфология данного элемента скелета у Plioperdix остается неопределенной. Локтевые кости у фазановых слабо диагностичны, по этой причине отнесение локтевых костей из Одесских катакомб к P. pontica нужно расценивать как предварительное. Фрагментарные лопатки из Одесских катакомб также отнесены к этому виду предварительно, поскольку у них не сохранился акромион, имеющий диагностичную форму у современных Coturnix. Проксимальная фаланга большого пальца крыла также предварительно отнесена к P. pontica.
Распространение. Верхи нижнего плиоцена–верхний плиоцен (верхняя часть зоны MN 15 и зона MN 16) Северного Причерноморья и Южной Польши. Достоверных данных о существовании P. pontica в нижнем плейстоцене (MN 17) нет.
Материал. Местонахождение Одесские катакомбы (верхняя часть MN 15): экз. NMNHU-P, Av-344, 666, 667, краниальные фрагменты правых коракоидов; экз. NMNHU-P, Av-676, 677, фрагменты лопаток; экз. NMNHU-P 5242, 5243, Av‑669, дистальные фрагменты плечевых костей; экз. NMNHU-P Av-342, 343, 670, О-2043, фрагментарные локтевые кости; экз. NMNHU-P, Av‑345, неполная лучевая кость; экз. NMNHU-P Av-340, 341, проксимальные фрагменты левых карпометакарпусов (ранее имели номера ИЗ НАНУ 6479, 6471, соответственно); экз. NMNHU-P Av-671, 672, дистальные фрагменты левых карпометакарпусов; экз. NMNHU-P Av-674, проксимальный фрагмент правого тарсометатарсуса. Местонахождение Котловина (Украина, Одесская обл.; нижний плиоцен или, возможно, нижний плейстоцен; MN 15–17; Тесаков, 2004): экз. NMNHU-P Av-698, 713, дистальные фрагменты правых плечевых костей; экз. NMNHU-P Av-699, 700, дистальные фрагменты левых плечевых костей; экз. NMNHU-P Av-707, дистальный фрагмент тарсометатарсуса. Местонахождение Рембелицы Крулевски 2 (Польша; верхний плиоцен, MN 16a): экз. б/н в колл. Ин-та систематики и эволюции животных (Краков), правый карпометакарпус без metacarpalia minor. Местонахождение Соколовский карьер (Украина, Кривой Рог; вероятно, плиоцен – судя по указанию Воинственского): экз. NMNHU-P Av-88, правая плечевая кость без проксимального эпифиза. Местонахождение Обуховка (Россия, Ростовская обл.; плиоцен; MN 15 или нижняя часть MN 16b; Nesin, Nadachowski, 2001; Tesakov et al., 2007): экз. NMNHU-P Av-683, дистальный фрагмент плечевой кости. Местонахождение Попово-1 (Украина; Запорожская обл., Васильевский р-н; нижний плейстоцен, верхи MN 16b; Рековец, Пашков, 2009): экз. NMNHU-P Av-684, краниальный фрагмент левого коракоида.
Замечания. Местонахождения Котловина и Обуховка – многослойные памятники с широким определением возраста в пределах верхнего русциния–виллафранка. Точное происхождение остатков P. pontica из этих местонахождений, к сожалению, остается неизвестным (это справедливо и по отношению к многослойному молдавскому местонахождению Этулия). Местонахождение Попово-1 первоначально было сопоставлено в верхами зоны MN 16 (Рековец, Пашков, 2009), но позднее обозначалось как MN 16/17 (Rzebik-Kowalska, Rekovets, 2016).
* * *
Многие материалы из плиоцен–плейстоценовых местонахождений Украины не могут быть отнесены к P. pontica, и относятся нами к нескольким другим видам и родам мелких фазановых.
?Eurobambusicola turolicus Zelenkov, 2016. Левый тарсометатарсус с поврежденным дистальным эпифизом (экз. NMNHU-P Av-486; рис. 1, д) из местонахождения Кучурган (Одесская обл.; нижний плиоцен, MN 14) сходен с современными Bambusicola общими удлиненными пропорциями, слабой расширенностью эпифизов относительно стержня, а также слабо выраженными fossae parahypotarsales. Эти же признаки прослеживаются и у позднемиоценового таксона Eurobambusicola turolicus из позднего миоцена местонахождения Полгарди (MN 13) Венгрии (за исключением не сохранившихся fossae). Близость возраста и сходство с современными Bambusicola позволяют предполагать, что экз. NMNHU-P Av-486 также принадлежал E. turolicus. Судя по размерам, к этому же виду относится и очень плохо сохранившийся правый коракоид (экз. NMNHU-P Av-485) из того же местонахождения.
Phasianidae indet. 1. Плечевая кость и тарсометатарсусы из местонахождения Войничево (экз. ПИН, №№ 2614/145, 146) сходны с P. hungarica, известным в данном регионе из местонахождения Морская-2 в Ростовской обл. (Zelenkov, Panteleyev, 2015), однако принадлежность мелких фазановых из Морской к этому виду, а также вообще отнесение вида hungarica к роду Plioperdix требуют подтверждения, поскольку, как показывает настоящая ревизия, предшествующие определения были основаны на некорректном представлении о морфологии P. pontica.
Phasianidae indet. 2. Большинство костных остатков из местонахождения Котловина также не могут быть отнесены к Plioperdix. Коракоиды (экз. NMNHU-P Av-690, 691, 692, 695, 709, 711) отличаются от таковых Plioperdix строением cotyla scapularis и сочленовной поверхности для акромиона лопатки (см. выше). Экз. NMNHU-P Av‑693, 694, 696 имеют очень плохую сохранность, в связи с чем информативное сравнение этих материалов невозможно.
Плечевая кость из Котловины (экз. NMNHU-P Av-697) не имеет заметно выступающего дорсально crista deltopectoralis и характерного для Coturnix четко очерченной дорсальной fossa tricipitalis. Карпометакарпусы из Котловины (экз. NMNHU-P Av-686, 716; ПИН, № 2614/137) характеризуются наличием вырезки в вентральном крае trochlea carpalis и проксимальной ямки в основании metacarpalia minor, а также изогнутостью metacarpalia major при виде с краниальной стороны. У других экземпляров (экз. NMNHU-P Av-717 и 718) вырезки как таковой не заметно из-за сохранности, но они отличаются от Plioperdix ориентацией вентрального края пястного блока (под углом около 130°, в отличие от 150°–160° у P. pontica). Проксимальный тарсометатарсус из местонахождения Котловина (экз. NMNHU-P Av-708) не может быть отнесен к P. pontica, поскольку характеризуется не выдающимся дорсально cotyla lateralis (см. выше). Кроме того, в отличие от P. pontica, стержень кости заметно суживается по направлению к дистальной части кости – как у Palaeocryptonyx edwardsi.
Серия костей из многослойного местонахождения Крыжановка (верхи MN 16b – низы MQ1; Тесаков, 2004), по-видимому, также не может относиться к P. pontica. Наиболее диагностичен из представленных на местонахождении костей коракоид (экз. NMNHU-P Av-654), который по форме facies articularis clavicularis и области cotyla scapularis идентичен коракоидам из Котловины (см. выше) и поэтому, несомненно, представляет этот же таксон.
Дистальный фрагмент правого тарсометатарсуса из Одесских катакомб, упомянутый Воинственским (1967) без номера (ныне обозначенный как NMNHU-P Av-675), не может относиться к Plioperdix pontica, так как его trochlea metatarsi IV слабо сдвинут плантарно (медиоплантарный угол trochlea metatarsi IV не выдается плантарнее trochlea metatarsi III, дорсомедиальный угол trochlea metatarsi IV приближен к дорсальному краю trochlea metatarsi III) и широко отставлен латерально (широкая incisura intertrochlearis lateralis). Кроме того, trochlea metatarsi III у этого экземпляра дорсовентрально низкий и асимметричными полублоками. По этим признакам экз. NMNHU-P Av-675 сходен с тарсометатарсусом Palaeocryptonyx edwardsi, от которого он отличается только заметно меньшими размерами. Таким образом, экз. NMNHU-P Av-675 можно было бы отнести к одновозрастному (раннеплиоценовому) P. donnezani, который сходен с P. pontica по размерам, но для P. donnezani тарсометатарсус пока не известен.
ОБСУЖДЕНИЕ
Предыдущие авторы (Курочкин, 1985; Bocheński, Kurochkin, 1987) отмечали сходство Plioperdix pontica с современными Coturnix, но все же признавали отсутствие между этими формами близкого родства, обращая внимание на ряд значительных морфологических отличий. Проведенная нами ревизия украинских материалов подтвердила морфологическую близость P. pontica и Coturnix; выявленные ранее отличия в основном были установлены по костным остаткам, принадлежащим другим таксонам фазановых. В то же время, некоторые отличительные признаки (напр., необычно удлиненная кисть) подтверждают родовую самостоятельность Plioperdix. В связи с уточнением морфологической характеристики P. pontica требуется подтверждение отнесения к Plioperdix других описанных в составе этого рода видов.
По нашим данным, P. pontica надежно установлен только из местонахождений верхов русциния – нижнего виллафранка (верхи MN 15–MN 16) Северного Причерноморья и Польши. В более древних местонахождениях русциния юга Украины (кучурганский фаунистических комплекс; MN 14) присутствует близкий по размерам перепел, сходный с “Plioperdix” hungarica, описанным из верхнего миоцена (MN 13) Венгрии. В составе кучурганского комплекса также присутствует ископаемая бамбуковая куропатка Eurobambusicola turolicus, также первоначально описанная из Венгрии. Материалы из местонахождения Крыжановка и значительная часть материалов из местонахождения Котловина представлены еще одной формой сходного размера (Phasianidae indet. 2); для установления ее точного систематического положения необходима ревизия материалов по другим виллафранкским мелким фазановым, описанным из Восточной Европы – в частности, Francolinus minor и Chauvireria balcanica (Jánossy, 1974; Boev, 1997).
Возраст мелких фазановых из Котловины требует обсуждения. Установлено, что в ранних сборах на местонахождении Котловина произошло перемешивание материала из разных возрастных горизонтов – от позднего русциния до среднего виллафранка (Тесаков, 2004). Примечательно, что в данном местонахождении определяются остатки как Plioperdix, так и других неопределенных фазановых (см. выше). По нашему мнению, кости Phasianidae indet. 2 из Котловины, относящиеся к ранним сборам, могут происходить из средневиллафранкских слоев (MN 17) – это могло бы объяснить доминирование этого таксона в данном местонахождении, в отличие от фауны Одесских катакомб (MN 15), где присутствует почти исключительно P. pontica. Данное предположение подтверждается сходством Phasianidae indet. 2 из Котловины с мелкими фазановыми из Крыжановки – местонахождения, также относимого преимущественно к MN 17 (Тесаков, 2004). Кроме того, мелкие фазановые Phasianidae indet. 2 из Котловины морфологически сходны с таковыми из еще одного многослойного и разновозрастного местонахождения Этулия (Молдова). Материалы по мелким фазановым из Этулии, описанные Курочкиным (1985) как P. pontica, были переданы в ПИН Н.А. Константиновой (Курочкин, 1985), которая изучала разрез, в настоящее время называемый Этулия-3 и также соотносимый с MN 17 (Тесаков, 2004). Таким образом, мелкая фазановая птица, описанная здесь как Phasianidae indet. 2, может быть характерна только для среднего виллафранка (MN 17) региона или, возможно, самых верхов MN 16. Мелкие фазановые из Котловины, отнесенные к P. pontica, в действительности могут происходить из более древних русцинийских слоев этого местонахождения. Отсутствие несомненных свидетельств сосуществования двух форм мелких фазановых (P. pontica и Phasianidae gen. indet. 2) в русцинии или виллафранке Северного Причерноморья подтверждает высказанное ранее предположение о возможности биохронологических корреляций континентальных фаун Восточной Европы по мелким курообразным птицам (Зеленков, 2015).
Список литературы
Воинственский М.А. Ископаемая орнитофауна Украины // Природная обстановка и фауны прошлого. Вып. 3 / Ред. Пидопличко И.Г. Киев: Наук. думка, 1967. С. 3–76.
Зеленков Н.В. Птицы и биохронология неогена Евразии // Современные проблемы палеонтологии. Матер. LXI сесс. Палеонтол. об-ва при РАН. СПб., 2015. С. 150–152.
Зеленков Н.В. Ревизия неворобьиных птиц Полгарди (Венгрия, верхний миоцен). 2. Galliformes // Палеонтол. журн. 2016. № 6. С. 79–91.
Зеленков Н.В., Курочкин Е.Н. Неогеновые фазановые (Aves: Phasianidae) Центральной Азии. 2. Роды Perdix, Ploperdix и Bantamyx // Палеонтол. журн. 2009. № 3. С. 79–86.
Зеленков Н.В., Курочкин Е.Н. Класс Aves // Ископаемые рептилии и птицы. Часть 3 / Ред. Курочкин Е.Н., Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2015. С. 86–290.
Курочкин Е.Н. Птицы Центральной Азии в плиоцене. М.: Наука, 1985. 119 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 26).
Рековец Л.И., Пашков А.В. Новые местонахождения микротериофауны позднего неогена Украины // Викопна фауна і флора України: палеоекологічний та стратиграфічний аспекти / Ред. Гожик П.Ф. Київ: Інст. Геол. наук НАН України, 2009. P. 354–360.
Серебровский П.В. Птицы из плиоценовых отложений Одессы // Докл. АН СССР. 1941. Т. 33. С. 476–479.
Тесаков А.С. Биостратиграфия среднего плиоцена-эоплейстоцена Восточной Европы (по мелким млекопитающим). М.: Наука, 2004. 247 с.
Тугаринов А.И. Новые находки плиоценовой орнитофауны Одессы // Докл. АН СССР. 1940. Т. 26. С. 311–313.
Bocheński Z. List of European fossil bird species // Acta Zool. Cracov. 1997. V. 40. № 2. P. 293–333.
Bocheński Z., Bocheński Zb.,Tomek T. A History of Polish Birds. Kraków: Inst. Syst. Evol. Anim. Pol. Acad. Sci., 2012. 226 p.
Bocheński Z., Kurochkin E.N. New data on Pliocene phasianids (Aves: Phasianidae) of Moldavia and S. Ukraine // Acta Zool. Cracov. 1987. V. 30. P. 81–96.
Boev Z. Chauvireria balcanica gen. n., sp. n. (Phasianidae — Galliformes) from the Middle Villafranchian of Western Bulgaria // Geol. Balcan. 1997. V. 27 № 3–4. P. 69–78.
Brodkorb P. Catalogue of fossil birds: Part 2 (Anseriformes through Galliformes) // Bull. Florida St. Mus. Biol. Sci. 1964. V. 8. P. 195–335.
Dickinson E.C., Remsen J.V., Jr. (Eds). The Howard and Moore Complete Checklist of the Birds of the World, V. 1: Non-Passerines. Eastbourne, U.K.: Aves Press, 2013. 461 p.
Göhlich U.B., Mourer-Chauviré C. Revision of the phasianids (Aves: Galliformes) from the Lower Miocene of Saint-Gerand-Le-Puy (Allier, France) // Palaeontology. 2005. V. 48. P. 1331–1350.
Jánossy D. Upper Pliocene and Lower Pleistocene bird remains from Poland // Acta Zool. Cracov. 1974. V. 19. P. 531–566.
Jánossy D. Late Miocene birds remains from Polgárdi (W‑Hungary) // Aquila. 1991. V. 98. P. 13–55.
Kretzoi M. Pliogallus Gaillard, 1939 es Pliogallus Tugarinov, 1940 // Aquila. 1955. V. 59/62. P. 367.
Mlíkovský J. Early Pleistocene birds of Stránská skála: 1. Musil’s talus cone // Anthropos. 1995. V. 26. P. 111–126.
Mlíkovský J. Cenozoic birds of the world. Pt 1: Europe. Praha: Ninox Press, 2002. 406 p.
Mourer-Chauviré C. [Review of] Cenozoic birds of the World, part 1: Europe. J. Mlíkovský. 2002 // Auk. 2004. V. 121. P. 623–627.
Mourer-Chauviré C., Geraads D. The upper Pliocene avifauna of Ahl al Oughlam, Morocco. Systematics and Biogeography // Rec. Austral. Mus. 2010. V. 62. P. 157–184.
Nesin V.A., Nadachowski A. Late Miocene and Pliocene small mammal faunas (Insectivora, Lagomorpha, Rodentia) of Southeastern Europe // Acta Zool. Cracov. 2001. V. 44. P. 107–135.
Rzebik-Kowalska B., Rekovets L.I. New data on Eulipotyphla (Insectivora, Mammalia) from the Late Miocene to the Middle Pleistocene of Ukraine // Palaeontol. Electron. 2016. Vol. 19.11.19A: 11-31.
Tesakov A.S., Vangengeim E.A., Pevzner M. Arvicolid zonation of continental Pliocene deposits of East Europe // Cour. Forsch. Senckenb. 2007. Bd 259. S. 227–236.
Zelenkov N.V., Panteleyev A.V. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia) // Paläontol. Z. 2015. V. 89. P. 515–527.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал