

Палеонтологический журнал, 2020, № 6, стр. 83-95
Редкие и эндемичные элементы в триасовых тетраподных сообществах Общего Сырта (Восточная Европа)
И. В. Новиков a, b, *, А. Г. Сенников a, b, **, А. В. Иванов c, ***
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Казанский федеральный университет
Казань, Россия
c Саратовский государственный технический университет им. Ю.А. Гагарина
Саратов, Россия
* E-mail: inovik@paleo.ru
** E-mail: sennikov@paleo.ru
*** E-mail: yashkovia@mail.ru
Поступила в редакцию 24.12.2019
После доработки 21.01.2020
Принята к публикации 21.01.2020
Аннотация
Известная для территории Общего Сырта история смены раннетриасовых тетраподных сообществ включает восемь последовательных фаунистических комплексов, что по дробности выделяемых эволюционных этапов не имеет аналогов в мире. Доминирующими элементами в этих тетраподных сообществах являются темноспондильные амфибии, среди которых наиболее широкое распространение имеют три группы капитозавроморф – капитозавриды, бентозухиды и трематозавриды. Гораздо реже встречаются остатки рептилий. В статье приводятся краткие сведения о находках редких и эндемичных групп тетрапод, известных из нижнего триаса Общего Сырта. К ним относятся квонтасиды, лонхоринхиды, капитозавриды-селенокарины, ритидостеиды, плагиозавры, некоторые формы бентозухид и трематозаврид, реликтовые антракозавры-хрониозухии – из амфибий, а также эозавроптеригии, териодонты и отдельные роды проколофонов, пролацертилий и текодонтов – среди рептилий. Подчеркивается своеобразие систематического состава раннетриасовых тетраподных сообществ Общего Сырта и Южного Приуралья по отношению к таковому одновозрастных фаун наземных позвоночных более северных регионов Восточной Европы, и правомерность выделения этих двух районов в отдельный регион, характеризующийся особенностями зоогеографических связей.
Триасовые отложения возвышенности Общий Сырт представлены исключительно континентальными фациями, отнесены к нижнему отделу и хорошо охарактеризованы остатками наземных позвоночных, которые играют ведущую роль при расчленении и корреляции. Выявленная для этой территории история смены раннетриасовых тетраподных сообществ включает восемь последовательных фаунистических комплексов, что по дробности выделяемых эволюционных этапов не имеет аналогов в мире. Наиболее крупные этапы – фауны – обозначаются по доминирующим родам темноспондильных амфибий как (в восходящем порядке): Tupilakosaurus (нижний инд, вохминский горизонт), Selenocara–Syrtosuchus (верхний инд, заплавненский горизонт), Benthosuchus (нижний оленек, рыбинский горизонт), Wetlugasaurus (нижний оленек, слудкинский и устьмыльский горизонты) и Parotosuchus (верхний оленек, федоровский и гамский горизонты), в составе последних трех из которых выделяются по две группировки (Новиков, 2018).
Доминирующими элементами в этих тетраподных сообществах, как уже указывалось выше, являются темноспондильные амфибии, среди которых наиболее широкое распространение имеют три группы капитозавроморф – капитозавриды, бентозухиды и трематозавриды. Гораздо реже встречаются остатки рептилий. Планомерные комплексные экспедиционные исследования последних двадцати лет, проводимые авторами на триасовых отложениях Общего Сырта, а также переизучение всего имеющегося с этой территории материала по триасовым тетраподам позволили получить новые данные о присутствии здесь редких и эндемичных групп. Среди амфибий это квонтасиды, лонхоринхиды, ритидостеиды, плагиозавры, капитозавриды-селенокарины и некоторые формы бентозухид и трематозаврид – из темноспондилов, а также реликтовые антракозавры-хрониозухии. Рептилийный материал в основном принадлежит проколофонам, текодонтам и пролацертилиям, среди которых также отмечены эндемичные и редкие формы. Вместе с ними были встречены единичные остатки териодонтов и такой экзотической для раннего триаса Восточной Европы группы, как эозавроптеригии. Ниже приводится краткая характеристика находок всех этих редких и эндемичных групп, а также обсуждается их роль в процессе восстановления тетраподного сообщества после позднепермского вымирания.
ТЕМНОСПОНДИЛЬНЫЕ АМФИБИИ
Квонтасиды (Qantasidae). Это монотипичное семейство достоверно известно только из семи местонахождений11 (Заплавное-Сосновое IV, Безымянный, Каменный Дол, Краснояр, Савельевка, Новая Таволжанка, Съезжая), приуроченных к заплавненскому, рыбинскому и слудкинскому горизонтам (Новиков, 2012а, 2018). Его типовой род (Qantas) по особенностям своей морфологии является выражением “архаического многообразия”, характерного для начального этапа дифференциации трематозавроидов. Мозаичность организации Qantas заключается в специфическом сочетании бентозухидных и трематозавридных черт (отмеченном из-за характера сохранности материала, главным образом, в строении нижней челюсти), к которым добавляется и ряд архаичных особенностей, необычных для триасовых Temnospondyli (присутствие шагрени и мелких зубов на всех короноидах и округлая форма оснований челюстных зубов) (Новиков, 2012а, 2018). Это своеобразие квонтасид выделяет их среди других трематозавроидов и затрудняет выяснение их родственных связей. Вероятно, предки этой группы довольно рано отделились от основного ствола трематозавроидов и развивались параллельно с другими группами надсемейства, показывая иной порядок преобразований в сторону трематозавридной организации. За пределами рассматриваемой территории к квонтасидам может относиться известный только по переднему концу морды монотипичный Tirraturhinus Nield, Damiani et Warren из низов триасового разреза Восточной Австралии (Nield et al., 2006).
Лонхоринхиды (Lonchorhynchidae) – наиболее аберрантная ветвь трематозавроидов, отличающаяся, в первую очередь, сильно удлиненным ростром и округлыми в сечении основаниями зубов. Эта космополитная группа уже при своем появлении в самом начале триаса показывает значительное систематическое разнообразие. В нижнем триасе за пределами Восточной Европы они известны из грисбахских отложений Индии, Восточной Австралии и Южной Африки, динера Пакистана, Восточной Гренландии и Австралии, а также из оленекских отложений Мадагаскара, Шпицбергена, Западной Австралии, Аризоны (США) и Южной Африки (см. Новиков, 2013, 2018). В среднем триасе фрагментарные находки лонхоринхид известны из анизия Индии, Канады и Алжира (Welles, 1993).
На территории Восточной Европы остатки лонхоринхид крайне редки и известны преимущественно (за исключением единственной находки в федоровском горизонте Московской синеклизы) из позднеиндских (заплавненский горизонт; местонахождения Заплавное-Березовое, Заплавное-Сосновое II, Никольское, Тупиковка) и раннеоленекских (рыбинский горизонт; местонахождение Мечеть I) отложений Общего Сырта (Новиков, 2013, 2018). Фрагментарный характер этих находок значительно затрудняет их определение ниже уровня семейства. Исключение составляет лишь материал из верхней части разреза местонахождения Никольское, относящейся к заплавненскому горизонту (см. Новиков, 2018). Изучение этого материала позволило М.А. Шишкину (2002) выделить новое подсемейство (первоначально – семейство) Cosgriffiinae с типовым родом (Cosgriffius Welles), описанным из верхов нижнего триаса Аризоны (Welles, 1993). При этом на основании сходства косгриффиид с палеозойскими архегозавроидами по архаичному типу сошников и наличию ложковидного расширения челюстного симфиза было высказано предположение о наличии между этими группами темноспондилов непосредственной родственной связи. Позднее, в результате новых сборов, отсюда же был описан фрагмент черепа лонхоринхида (экз. ПИН, № 4454/8), предположительно отнесенный к гренландскому роду Stoschiosaurus Säve-Söderbergh по строению заглазничной области (Новиков, 2018).
Бентозухиды (Benthosuchidae). Среди бентозухид несомненными эндемиками Общего Сырта являются сыртозухины и архаичный вид типового рода – Benthosuchus gusevae Novikov. Остатки представителей монотипичного подсемейства Syrtosuchinae широко распространены на территории Общего Сырта и известны более чем из 20 местонахождений, приуроченных преимущественно22 к заплавненскому горизонту (Новиков, 2016, 2018). Для сыртозухин, как представителей наиболее архаичных трематозавроидов, характерна слабая выраженность типично трематозавроидных признаков (напр., парности переднего небного отверстия, смещения супраорбитального канала на lacrimale). С другой стороны, типовой род этого подсемейства (Syrtosuchus Novikov: рис. 1, а, б) по целому ряду признаков (мелкоячеистый характер покровного орнамента, строение и положение crista obliqua, слабое развитие системы боковой линии и др.) показывает значительное сходство с архаичными капитозавридами–селенокаринами, так же широко распространенными в одновозрастных отложениях Общего Сырта (см. ниже). Такая близость структурного плана может указывать на происхождение Syrtosuchus от селенокарин или наличие у них непосредственного общего предка.
Рис. 1.
Остатки раннетриасовых амфибий из нижнего триаса Оренбургской обл.: а, б – Syrtosuchus sp., экз. ПИН, № 4629/10, неполный череп: a – сверху, б – снизу; Бузулукский р-н, меcтонахождение Суриково II; заплавненский горизонт; в – Melanopelta sp., экз. ПИН, № 5497/27, тело туловищного позвонка спереди; Илекский р-н, меcтонахождение Дьяконов II; федоровский горизонт. Обозначения: cch – канал хорды, fpa – foramen palatinum anterior, Ps – parasphenoideum, Pt – pterygoideum. Длина масштабной линейки 10 мм.

Benthosuchus gusevae является ведущим элементом одноименной группировки фауны Benthosuchus, известной только на рассматриваемой территории (местонахождения Алексеевка I, Алексеевка II, Безымянный, Глинный II, Каменный II, Каменный Дол, Каменный овраг, Красная Яруга II, Репный). Несмотря на то, что эта форма является уже представителем типичных бентозухид – бентозухин, она сохраняет ряд наиболее полно выраженных капитозавроидных черт (параболические очертания черепа, расширенные и уплощенные задние отростки сошников, расположение межхоанального зубного ряда в виде широкого клина, укороченный симфизный отдел нижней челюсти), подобно селенокаринам и другим ранним капитозавридам – ветлугазавринам, причем среди селенокарин этот вид наиболее близок с типовым родом (Новиков, 2012б, 2018). С другой стороны, B. gusevae также обнаруживает значительное сходство с появившимися несколько раньше сыртозухинами по таким признакам (помимо типично бентозухидных), как слабое развитие системы боковой линии, мелкоячеистый характер покровного орнамента, расположение foramen chordae tympani близко к уровню наружного супраангулярного отверстия, постоянное присутствие зубов на передних короноидах, а также расширенные и уплощенные задние отростки сошников. Однако непосредственная филогенетическая преемственность Syrtosuchus и Benthosuchus представляется маловероятной, и обе ветви бентозухид – сыртозухины и бентозухины – могут иметь происхождение от общего капитозавроидного предка, морфотип которого близок к таковому Selenocara (Новиков, 2018).
Трематозавриды (Trematosauridae). Это несомненно монофилетичное и лавразийское по происхождению семейство представлено в триасе Общего Сырта тремя подсемействами (Thoosuchinae Getmanov, Platysteginae Schoch et Milner и Trematosaurinae Watson), известными и из других регионов Восточной Европы. При этом тоозухин Prothoosuchus Getmanov и платистегин Trematotegmen Getmanov достоверно известны только на рассматриваемой территории. Остатки наиболее архаичного и раннего среди всех трематозаврид рода Prothoosuchus довольно редки, и известны из шести местонахождений (Корольки, Федоровка, Девятичный II, Каменный Яр – Сухушка, Новая Таволжанка, Корнеевка II), приуроченных к широкому стратиграфическому интервалу (от заплавненского до устьмыльского горизонтов). Специфическим для этого рода является присутствие ряда педоморфных черт (маленькие размеры; относительно крупные орбиты, расположенные на уровне середины длины черепа; приближенность пинеального отверстия к уровню задних краев орбит; наличие эктоптеригоидных клыков; вхождение наружных крыловидных костей в края межптеригоидных ям и др.) в сочетании со взрослыми (окостеневавшие плевроцентры), и иллюстрирует влияние педоморфоза на формирование трематозаврового плана организации, впервые отмеченного М.А. Шишкиным и Т. Сулей (Shishkin, Sulej, 2009).
Монотипичный род Trematotegmen, впервые описанный из нижнего триаса Общего Сырта (Гетманов, 1982) и известный здесь33 только из трех слудкинских по возрасту местонахождений (Кострикин Дол, Нижнеозерное II, Прокаевка), является среди платистегин наиболее продвинутым в сторону трематозавровой организации. Это выражается в более высоком затылке, строении exoccipitale (редукция нижнеушного отростка и его субпродольная ориентация), слабом развитии crista falciformis, врастании видиева канала в тело парасфеноида, и в особенностях системы боковой линии (полное развитие, очень широкие и глубокие желобки, отсутствие flexura lacrimalis). Вместе с тем этот род обнаруживает ряд признаков (прежде всего, сильная укороченность костей постпариетального ряда), отличающих его как от остальных примитивных трематозаврид (тоозухин, платистегин Angusaurus Getmanov и Platystega Wiman), так и от собственно трематозаврин, и указывающих на его отклонение от магистрального пути становления трематозаврового морфотипа (Новиков, 2018).
Селенокарины (Selenocarinae) включают в себя позднеиндские роды Selenocara Bjerring и Samarabatrachus Novikov, характеризующиеся наиболее примитивной морфологией среди капитозаврид, широко представленные в заплавненском горизонте Общего Сырта (обе формы) и известные по единичным находкам в верхнеиндском подъярусе Восточной Гренландии (Selenocara) (Новиков, 2016). Ряд структурных особенностей этого подсемейства (общих или мозаично выступающих у того или иного из двух его родов) сближает его с древнейшими капитозавроморфами–ринезухидами, а также с другими палеозойскими темноспондилами. Так, оба рода селенокарин характеризуются рядом типично ринезухидных черт: слабое развитие преорбитального отдела черепной крыши, положение заднего края supratemporale каудальнее уровня вершины ушных вырезок, расположение вершин межптеригоидных окон у заднего края хоан, сильное развитие шагрени на небной поверхности, относительно длинная тимпанальная часть затылочного фланга squamosum, резкая выраженность crista tympanica, сильное окостенение basioccipitale. Следует также отметить присутствие у селенокарин такой архаичной черты, как наличие пре- и посторбитальных продольных валиков и бугров на дорсальной поверхности крыши черепа.
Кроме того, каждый из родов селенокарин дополнительно показывает и другие примитивные черты, необычные для триасовых капитозавроидов, в большинстве также разделяемые с ринезухидами и другими палеозойскими темноспондилами. Для рода Selenocara этими чертами, дополнительно подчеркивающими мозаичный характер его морфотипа, являются: дорсолингвальная ориентировка поверхности костей короноидной серии, слабое развитие ретроартикулярного отростка нижней челюсти (рис. 2), участие basioccipitale в строении затылочного мыщелка, а также присутствие шагрени на небных костях и эктоптеригоидах (помимо парасфеноида и небных ветвей птеригоидов). Еще одним примитивным признаком, известным у Selenocara, является прямолинейное очертание вентрального контура нижней челюсти позади ангулярного изгиба (рис. 2, а, б).
Рис. 2.
Selenocara rossica Novikov, экз. ПИН, № 2426/12, левая ветвь нижней челюсти: а – с лабиальной стороны, б – с лингвальной стороны, в – сверху; Самарская обл., Борский р-н, меcтонахождение Заплавное-Сосновое II; нижний триас, заплавненский горизонт. Обозначения: An – angulare, Ar – articulare, Cor – coronoideum, D – dentale, fcht – foramen chordae tympani, fmp – foramen meckelianum posterior, fsa – foramen supraangularis, Icor – intercoronoideum, pra – processus retroarticularis, Psp – postspleniale, San – supraangulare, sdpr – sulcus depressorius, Sp – spleniale. Длина масштабной линейки 10 мм.
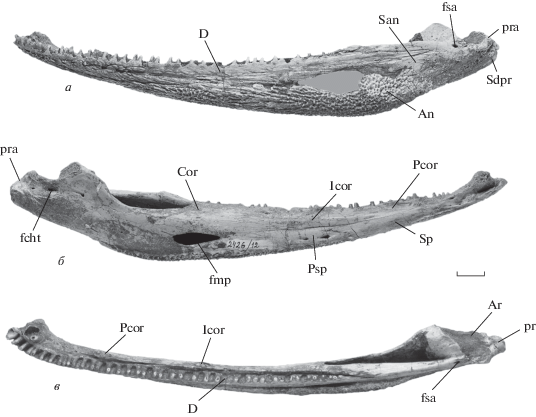
К дополнительным плезиоморфным признакам ринезухидного уровня, наблюдаемым среди селенокарин у рода Samarabatrachus, относятся коленообразная форма слезного изгиба (flexura lacrimalis) инфраорбитального сенсорного канала (известна также у лидеккеринид) и наличие срединного разделенного промежутка между задними отделами сошников (fodina vomeralis), характерного также для большинства триасовых капитозавроидов, кроме ветлугазаврин.
С другой стороны, селенокарины резко противопоставляются наиболее ранним гондванским представителям семейства (род Rewanobatrachus Schoch et Milner) по строению crista obliqua восходящей пластины птеригоида (низко расположенный, полого ориентированный в затылочном плане, с округлым сечением верхнего края – у селенокарин44 и высокий, заостренный и круто направленный – у Rewanobatrachus), подтверждая мнение ряда авторов (Очев, 1966; Maryańska, Shishkin, 1996) о наличии двух независимых линий в развитии капитозаврид – лавразийской и гондванской.
Ритидостеиды (Rhytidosteidae). Гондванские по происхождению, ритидостеиды представлены на территории Восточной Европы (Общий Сырт, Южное Приуралье и Прикаспий) только родом Rhytidosteus (Сенников, Новиков, 2012), описанным первоначально по единственному образцу из Южной Африки (R. capensis: Owen, 1884). В триасе Общего Сырта их присутствие в виде фрагментарных остатков отмечено только в двух местонахождениях яренского (позднеоленекского) возраста – Дьяконов I (R. uralensis Shishkin) и Дьяконов II (Rhytidosteus sp.) (Новиков, 2018). Широкое распространение остатков ритидостеид в отложениях прибрежно-морского генезиса может указывать на распространение Rhytidosteus из гондванских областей (Южная Африка) на территорию Восточной Европы через прибрежные биотопы вдоль северного побережья Тетиса (Шишкин, 2010; Новиков, 2018).
Плагиозавриды (Plagiosauridae). Остатки плагиозавров, выделяющихся среди других групп темноспондилов, помимо особенностей морфологии черепа и плечевого пояса, также уникальным строением тел позвонков, в раннетриасовых тетраподных сообществах Восточной Европы редки, и известны, в основном, из яренских (позднеоленекских) отложений Московской и Мезенской синеклиз. В нижнем триасе более южных регионов до недавнего времени было известно лишь по одной их находке в Южном Приуралье и в бассейне р. Дон, причем все они с некоторой долей условности были отнесены к плагиостернину Melanopelta (Шишкин, 1967; Новиков, 2018). В 2017 г. была сделана первая находка остатков плагиозавров и в нижнем триасе Общего Сырта (местонахождение Дьяконов II). Она представляет собой слабо амфицельное и дорсовентрально уплощенное тело позвонка с хордальным каналом (экз. ПИН, № 5497/27: рис. 1, в). По своему строению этот образец обнаруживает значительно сходство с телом позвонка (экз. ПИН, № 2394/3), описанным Шишкиным (1967) из Южного Приуралья (местонахождения Кзыл-Сай II). Учитывая вероятные гондванские корни, ограниченность ареала распространения средне- и позднетриасовых форм (Европа, Шпицберген, Гренландия, Таиланд), а также приуроченность многих местонахождений этого возраста к отложениям эпиконтинентальных морских бассейнов, вполне возможно расселение плагиозавров из Гондваны, как и в случае с ритидостеидами, через прибрежно-морские биотопы (Shishkin, 2007; Новиков, 2018).
ХРОНИОЗУХИИ
Реликтовые антракозавры–хрониозухии в триасе Восточной Европы известны по редким находкам, в основном дермальных щитков (остеодерм) и позвонков, представителей семейства Bystrowianidae (Novikov, Shishkin, 2000; Шишкин и др., 2014; Новиков, 2018). Их присутствие задокументировано почти во всех (кроме слудкинского) выделяемых здесь региональных стратиграфических подразделениях, охватывающих интервал от нижнеиндского подъяруса нижнего триаса (вохминский горизонт) до ладинского яруса среднего триаса (букобайский горизонт). За пределами Восточной Европы известен лишь один триасовый быстровианид (монотипичный род Bystrowiella Witzmann, Schoch et Maisch), описанный из нижнего кейпера Южной Германии (Witzmann et al., 2008).
В триасе Общего Сырта единичные находки быстровианид происходят из двух стратиграфических уровней – вохминского (местонахождения Алексеевка III, Перевозинка, Переволоцкое) и рыбинского (местонахождения Мечеть I, Мечеть II, Каменный Дол и Красная Яруга II) горизонтов. Все известные отсюда вохминские образцы условно (из-за отсутствия наиболее диагностичной части скелета – остеодерм) отнесены к роду Axitectum (A. vjushkovi Shishkin et Novikov и Axitectum sp.). Этот род, характеризующийся узкими, продольно вытянутыми щитками, впервые был описан из Московской синеклизы, где он встречен в шести местонахождениях, приуроченных к вохминскому (A. vjushkovi) и федоровскому (A. georgi Novikov et Shishkin) горизонтам (Novikov, Shishkin, 2000; Новиков, 2018). Материал по быстровианидам из рыбинских по возрасту местонахождений Общего Сырта представлен как щитками (местонахождения Мечеть I и Мечеть II), так и отдельными позвонками (местонахождения Каменный Дол и Красная Яруга II), при этом родовая принадлежность (Dromotectum) установлена только для остеодерм (Dromotectum sp. и D. spinosum Novikov et Shishkin)55. Последние имеют сильный сводообразный изгиб в поперечном сечении и незначительно расширены в плане. За пределами Общего Сырта представители рода Dromotectum (D. abditum Shishkin, Novikov et Fortuny) известны только из гамского горизонта бассейна р. Дон66 (Шишкин и др., 2014).
ПРОКОЛОФОНЫ
Проколофоны составляют обычный компонент триасовых тетраподных сообществ в северных регионах Восточной Европы (Московская, Мезенская и Печорская синеклизы, Северное Приуралье). Однако на территории Общего Сырта их находки единичны. Это в равной степени относится к обоим встреченным на территории Восточной Европы семействам – Spondylolestidae и Procolophonidae. Представители первого из этих семейств, характеризующегося недифференцированной зубной системой с простыми коническими зубами, достоверно известны по единичным находкам из трех вохминских местонахождений – Никольское (Contritosaurus sp.), Переволоцкое (Contritosaurus sp.) и Яблоновый Враг (Phaanthosaurus sp.), причем образцы из последнего местонахождения (см. Блом, 1968) в настоящее время утеряны. Предположительно спондилолестиды встречены и в одновозрастном местонахождении Перевозинка, однако происходящий отсюда материал (посткраниальные остатки) не диагностичен. Более продвинутые проколофоны из семейства Procolophonidae (с дифференцированной на квазирезцы и двухвершинковые квазикоренные зубной системой) представлены двумя родами – широко распространенным в Восточной Европе как географически, так и стратиграфически (рыбинский – федоровский горизонты) родом Tichvinskia и эндемичным родом Samaria. Остатки Tichvinskia определены из трех местонахождений – Мечеть II, Мойка II (оба приурочены к рыбинскому горизонту) и Дьяконов II (федоровский горизонт). Из первого местонахождения происходят находки Т. cf. jugensis Vjuschkov et Tchudinov (экз. ПИН, № 2424/2, фрагмент черепа), а также фрагмент сочлененного посткраниального скелета неопределимого проколофона (экз. ПИН, № 2424/41). Остатки Tichvinskia sp. из местонахождения Мойка II представлены фрагментами верхне- и нижнечелюстной костей и частью сочлененного посткраниального скелета (экз. ПИН, № 3890/5–7), вероятно, принадлежавшими одной особи (Новиков и др., 2017), а из местонахождения Дьяконов II – фрагментом нижней челюсти (экз. ПИН, № 5497/21).
Монотипичный род Samaria, впервые описанный из низов рыбинского горизонта77 Общего Сырта (группировка Benthosuchus gusevae) является древнейшим представителем проколофонид в Восточной Европе. Несмотря на это, он уже показывает хорошо дифференцированную зубную систему и специфическую форму квазикоренных зубов (сильно расширены поперечно, с максимальной шириной на уровне нижней трети высоты). Помимо типового местонахождения, типовой вид рода S. concinna (Ivachnenko) известен (экз. ПИН, № 3892/2, нижнечелюстная кость) из верхней части рыбинского горизонта (группировка Benthosuchus sushkini; местонахождение Липовка).
ЭОЗАВРОПТЕРИГИИ
Эта крайне редкая для триаса Восточной Европы группа до недавнего времени была известна по находкам из двух местонахождений (Сенников, 2001) – раннетриасового (позднеоленекского) Донская Лука (циматозаврид Tanaisosaurus kalandadzei Sennikov) и среднетриасового (ладинского) Большая Сыня II (пистозаврид Pistosaurus sp.).
Остатки эозавроптеригий недавно установлены нами из двух раннеоленекских (рыбинский горизонт) местонахождений Общего Сырта. Так, среди материалов, собранных В.П. Твердохлебовым (Саратовский государственный ун-т) из местонахождения Нижнеозерное, был обнаружен коракоид ближе не определимой эозавроптеригии (экз. ПИН, № 4734/5; рис. 3, б), а из местонахождения Алексеевка II определена квадратная кость (экз. ПИН, № 4513/55; рис. 3, а), сходная с таковой Tanaisosaurus kalandadzei (Сенников, Новиков, 2014). Эти новые материалы, вероятно, представляют собой самые древние находки эозавроптеригий на территории Восточной Европы и в мире. Учитывая континентальное (аллювиальное) происхождение указанных местонахождений, можно предположить, что наиболее ранние эозавроптеригии обитали не только в прибрежных биотопах на морском мелководье, но могли заходить и в континентальные водоемы. Так как конечности эозавроптеригий были преобразованы в ласты [и это исключало возможность передвижения по суше (Сенников, 2001)], то они, вероятно, заселяли достаточно крупные, не пересыхающие водоемы, существовавшие, очевидно, в раннетриасовую эпоху на территории Общего Сырта. Последняя в это время представляла собой обширную сильно обводненную предгорную равнину, где имел место значительный сток с молодых Уральских гор (Мазарович, 1936; Блом, 1972).
Рис. 3.
Остатки раннетриасовых рептилий: а, б – Eosauropterygia fam. indet.: а – экз. ПИН, № 4513/55, левая квадратная кость спереди; Самарская обл., Борский р-н, местонахождение Алексеевка II; рыбинский горизонт; б – экз. ПИН, № 4734/5, левый коракоид сверху; Оренбургская обл., Илекский р-н, местонахождение Нижнеозерное; рыбинский горизонт; в – Vritramimosaurus dzerzhinskii Sennikov, голотип ПИН, № 951/72, шейный позвонок, вид слева; Оренбургская обл., Илекский р-н, местонахождение Рассыпная; гамский горизонт; г – Chasmatosuchus rossicus F. von Huene, лектотип ПИН, № 2252/381, два сочлененных переднетуловищных позвонка, вид справа; Вологодская обл., Никольский р-н, местонахождение Вахнево; рыбинский горизонт; д – Chasmatosuchus sp., экз. ПИН, № 3362/29, заднешейный–переднетуловищный позвонок, вид справа; Самарская обл., Тоцкий р-н, местонахождение Безымянный; рыбинский горизонт; е – Tsylmosuchus jakovlevi Sennikov, 1990, голотип ПИН, № 4332/1, шейный позвонок, вид справа; Республика Коми, Усть-Цилемский р-н, местонахождение Черепанка-III; устьмыльский горизонт; ж, з – Tsylmosuchus samariensis Sennikov, неполные шейные позвонки, вид справа: ж – голотип ПИН, № 2424/6, з – экз. ПИН, № 2424/7; Оренбургская обл., Бузулукский р-н, местонахождение Мечеть II; рыбинский горизонт.

ПРОТОРОЗАВРЫ
Остатки проторозавров довольно многочисленны на Общем Сырте в стратиграфическом интервале от заплавненского до устьмыльского горизонта и представлены посткраниальными остатками (позвонки, кости конечностей) пролацертида Microcnemus – широко распространенной формы в одновозрастных отложениях других регионов Восточно-Европейской платформы и Южного Приуралья. При этом остатки Microcnemus из общесыртовского разреза существенно не отличаются от типовой серии Microcnemus efremovi F. von Huene (von Huene, 1940), поэтому можно предположить, что здесь представлен именно этот вид.
Находки более молодых триасовых проторозавров на Общем Сырте известны лишь из местонахождения Рассыпное (гамский горизонт, верхний оленек), откуда описан эндемичный пролацертид Vritramimosaurus dzerzhinskii Sennikov (Сенников, 2005; рис. 3, в). Эта форма является самым крупным проторозавром из триаса Восточной Европы, крупнее одновозрастного танистрофеида Augustaburiania vatagini Sennikov из местонахождения Донская Лука и даже среднетриасового (ладинского) пролацертида Malutinisuchus gratus Otschev из Северного и Южного Приуралья. Присутствие в одновозрастных (гамских), но разных по происхождению отложениях двух форм проторозавров (пролацертида Vritramimosaurus dzerzhinskii – в континентальных и танистрофеида Augustaburiania vatagini – в прибрежно-морских) хорошо согласуется с различием их образа жизни – наземного для первой формы и полуводного – для второй.
ТЕКОДОНТЫ
Остатки текодонтов обнаружены на территории Общего Сырта во множестве местонахождений. Они относятся к протерозухидам (Proterosuchidae), ранним рауизухидам (Rauisuchidae) и гондванским по происхождению эритрозухидам (Erythrosuchidae). Первая из этих групп представлена родом Chasmatosuchus F. von Huene, известным по изолированным позвонкам и костям конечностей из нескольких местонахождений заплавненского–устьмыльского возраста. Более точное (до вида) определение остатков известных отсюда представителей этого рода до сих пор не представляется возможным. В то же время, они обнаруживают ряд особенностей, отличающих их от таковых из более северных регионов (Московская, Мезенская и Печорская синеклизы; рис. 3, г). В первую очередь необходимо отметить в среднем более крупные размеры позвонков и костей конечностей общесыртовских хасматозухов (напр., позвонка из местонахождения Безымянный: экз. ПИН, № 3362/29; рис. 3, д). Остатки Chasmatosuchus из разных регионов Восточной Европы обнаруживают и некоторые различия в морфологии подвздошных костей. Для образцов из мечетинской группы местонахождений Общего Сырта (экз. ПИН, №№ 2424/44, 70 и др.) эти кости имеют более продвинутые черты строения, несколько сближающие их с таковыми эритрозухид, – более глубокая вертлужная впадина, несколько более протяженный и сильнее выступающий супраацетабулярный гребень, более обособленная дорсальная пластина подвздошной кости, отделенная от тела менее широкой шейкой, более развитый передний отросток дорсальной пластины, более выпуклый ее верхний край.
В противоположность этому, подвздошные кости представителей Chasmatosuchus из Мезенской синеклизы (напр., из местонахождения Родома I: экз. ПИН, № 3716/2) имеют более примитивный протерозухидный облик. Несмотря на эти различия, отсутствие подвздошных костей в типовой серии единственного известного на сегодняшний день вида Chasmatosuchus88 – C. rossicus – не позволяет отнести именно к этому виду или северный, или южный морфотип, как и выделить новый вид. Возможно, в будущем, с получением более массовой, разнообразной и представительной выборки остатков хазматозухов из различных регионов Восточной Европы, появится возможность разрешить вопрос о наличии в составе рода Chasmatosuchus одного (C. rossicus) или нескольких видов. К протерозухиям, вероятно, относится и мелкий архозавроморф Exilisuchus tubercularis Otschev, известный из местонахождения Каменный Яр (рыбинский горизонт) по неполной правой подвздошной кости (экз. ПИН, № 4197/61: Сенников, 2008). Эта форма, впервые описанная по единственному образцу (неполной левой подвздошной кости своеобразного строения) из нижнего триаса Южного Приуралья и отнесенная к протерозухиям условно (Очев, 1979; Сенников, 1995, 2008), является крайне редким элементом раннетриасовых тетраподных сообществ Общего Сырта и Южного Приуралья.
Рауизухиды представлены двумя родами – Tsylmosuchus (T. samariensis Sennikov из рыбинского горизонта и T. jakovlevi Sennikov из устьмыльского горизонта: Сенников, 1990, 1995, 2008) и Jaikosuchus [J. magnus (Otschev) из гамского горизонта: Сенников, 1995]. Типовой вид первого из этих родов (T. jakovlevi), широко распространенный в слудкинских и устьмыльских отложениях северных регионов Восточной Европы (Московская, Мезенская и Печорская синеклизы), на территории Общего Сырта известен только из устьмыльского горизонта (местонахождения Логачевка и Мансурово). Шейный позвонок из первого местонахождения (экз. ПИН, № 4196/2) отличается от аналогичных позвонков этого вида из северных регионов более сильно выступающим килем в виде одинарного вентрального гребня.
Tsylmosuchus samariensis является эндемиком Общего Сырта и известен из ряда местонахождений [Мечеть II (голотип ПИН, № 2424/6; экз. ПИН, № 2424/7; рис. 3, ж, з), Каменный Дол, Каменный Яр, Безымянный и др.]. Этот вид, помимо других черт строения (см. Сенников, 1990), отличается от Tsylmosuchus sp. из одновозрастного местонахождения Вахнево и федоровского местонахождения Окуневское (Московская синеклиза), T. jakovlevi из слудкинского и усть-мыльского горизонтов Московской, Мезенской и Печорской синеклиз (рис. 3, е) и от T. donensis Sennikov из гамского горизонта Донской Луки более крупными размерами (Сенников, 1990, 1995, 2008). Здесь один из авторов (АГС) усматривает аналогию в различии размерных классов представителей как Tsylmosuchus, так и Chasmatosuchus, известных из южных (Общий Сырт и Южное Приуралье) и северных (Московская, Мезенская, Печорская синеклизы) регионов Восточной Европы.
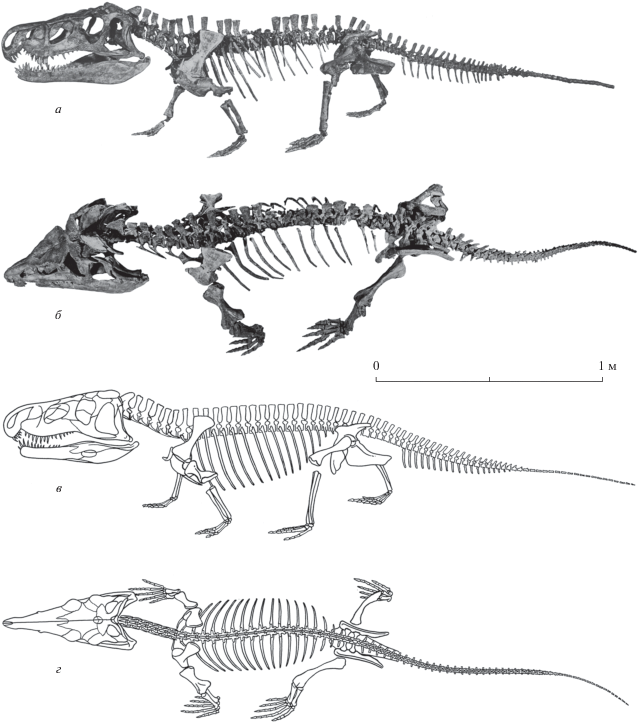
Рауизухид Jaikosuchus magnus (Otschev) и ранний эритрозухид Garjainia triplicostata (F. von Huene) известны только из местонахождения Рассыпное гамского возраста, причем последняя форма представлена здесь цельно скелетным материалом (рис. 4; Huene, 1960; Очев, 1979; Сенников, 1995, 2008). Помимо Общего Сырта, остатки этих двух родов встречены в местонахождениях федоровского и гамского возраста только в Южном Приуралье. При этом Garjainia triplicostata обнаруживает значительное сходство с G. prima Otschev, описанным из федоровского горизонта последнего региона.
Рис. 4.
Garjainia triplicostata (F. von Huene): а, б – фотография сборного, частично реконструированного скелета (по лектотипу ПИН, № 951/ 59 и экз. ПИН, №№ 951/1–40, 54–60): а – вид слева, б – вид сверху; в, г – рисунок скелета: в – вид слева, г – вид сверху; Оренбургская обл., Илекский р-н, местонахождение Рассыпное; нижний триас, гамский горизонт.
ТЕРИОДОНТЫ
Териодонты являются наиболее редкими элементами раннетриасовых тетраподных сообществ Восточной Европы. В нижнем триасе Общего Сырта они известны только из двух местонахождений, приуроченных к позднеоленекским отложениям. В первом из них (местонахождение Рассыпная) был найден череп тероцефала, описанный как Silphedosuchus orenburgensis Tatarinov (Татаринов, 1977; Ivakhnenko, 2011). Вторая находка этой группы была сделана авторами в 2011 г. на общесыртовском местонахождении Дьяконов II (верхи федоровского и низы гамского горизонтов). Она представляет собой фрагмент зубной кости (без зубов) мелкого териодонта (экз. ПИН, № 5497/26), ближе не определимого.
* * *
Новые данные по раннетриасовым тетраподным сообществам Общего Сырта, с одной стороны, дополнительно подчеркивают их своеобразие и отличия от одновозрастных фаун более северных районов Восточной Европы, с другой, – близость к тетраподным сообществам Южного Приуралья, а по отдельным элементам показывают сходство с триасовыми фаунами более удаленных регионов (Южная Африка, Восточная Гренландия, Австралия). Эти обстоятельства позволяют рассматривать Общий Сырт и Южное Приуралье в качестве особого (южноприуральского) биогеографического региона, характеризующегося особенностями зоогеографических связей (Шишкин и др., 2006; Новиков, 2018). На протяжении почти всего раннего триаса этот регион выделялся сохранением фаунистических связей с гондванскими областями. Так, в позднеиндском тетраподном сообществе Общего Сырта (фауна Selenocara–Syrtosuchus) отмечено присутствие ряда форм (древнейшие капитозавриды–ceленокарины Selenocara и Samarabatrachus), имеющих южногондванские родственные связи. Кроме того, именно к этому времени относится первое появление трематозавроида Qantas, обнаруживающего наибольшее сходство с плохо известным австралийским Tirraturhinus (Новиков, 2012а, 2018). Время существования фауны Parotosuchus характеризуется новой инвазией на территорию Восточной Европы тетраподных групп, имеющих гондванское происхождение (среди амфибий это ритидостеиды и, вероятно, плагиозавриды, а среди рептилий – эритрозухиды), причем присутствие ритидостеид и эритрозухид зафиксировано только в пределах южноприуральского биогеографического региона, где эти группы представлены общими с Южной Африкой родами – Rhytidosteus и Garjainia, соответственно.
Результаты изучения раннетриасовых тетраподных фаун Общего Сырта также позволили решить ряд вопросов по становлению, родственным связям и путям диверсификации основных групп амфибий на территории Восточной Европы. Проведенный недавно анализ всех имеющихся данных позволил сделать вывод о том, что территория Общего Сырта может рассматриваться в качестве центра происхождения трех близко родственных доминантных групп раннетриасовых темноспондилов, представленных семействами Capitosauridae99, Benthosuchidae и Trematosauridae (Новиков, 2018). Основой для такого заключения, помимо сведений о наиболее древних находках представителей каждой из этих групп (Selenocara и Samarabatrachus – среди капитозаврид, Syrtosuchus – среди бентозухид и Prothoosuchus – среди трематозаврид), учитывался и тот факт, что именно Общий Сырт является областью максимальной структурной диверсификации (“архаического многообразия”: Шишкин, 2011) каждого из этих семейств.
В заключение следует отметить, что для территории Общего Сырта наибольшее таксономическое разнообразие триасовых тетрапод, так же как и наибольшее число их местонахождений и остатков, приходится на начало раннеоленекского времени – времени существования фауны Benthosuchus. Местонахождения с фауной Tupilakosaurus немногочисленны, что может быть связано с ограниченным распространением здесь отложений раннеиндского возраста. В то же время, широко распространенные местонахождения фауны Wetlugasaurus (слудкинский горизонт) гораздо беднее рыбинских. Вероятно, это связано с наиболее благоприятными условиями для существования тетраподной биоты на территории Общего Сырта именно в рыбинское время, которое знаменуется максимумом раннеоленекской бореальной трансгрессии во всем северном полушарии (Lozovsky, 1989). Такие условия, предположительно, и были обусловлены гумидизацией климата, на что может указывать большая сероцветность рыбинских отложений по сравнению с выше- и нижележащими.
Работа выполнена при финансовой поддержке РФФИ (проекты №№ 17-04-00410 и 17-54-10013), Министерства образования и науки России (инициативный проект № 5.5177.2017/8.9.), за счет субсидии, выделенной Казанскому федеральному университету для выполнения государственного задания № 671-2020-0049 в сфере научной деятельности, а также в рамках государственной программы повышения конкурентоспособности Казанского (Приволжского) федерального университета среди ведущих мировых научно-образовательных центров и программы фундаментальных научных исследований Президиума РАН № 22 “Эволюция органического мира и планетарных процессов”. ИВН и АГС выражают благодарность ОАО “Оренбургэнерго” за всестороннюю поддержку проведения экспедиционных исследований, а также любителям-палеонтологам В.В. Константинову, А.В. Груздеву и членам Самарского палеонтологического общества за переданные материалы по тетраподам.
Список литературы
Блом Г.И. Каталог местонахождений фаунистических остатков в нижнетриасовых отложениях Среднего Поволжья и Прикамья. Казань: Изд-во Казан. ун-та, 1968. 376 с.
Блом Г.И. Фации и палеогеография Московской синеклизы и Волжско-Камской антеклизы в раннетриасовую эпоху. Казань: Изд-во Казан. ун-та, 1972. 368 с.
Гетманов С.Н. Лабиринтодонт из нижнего триаса Общего Сырта // Палеонтол. журн. 1982. № 2. С. 103–108.
Мазарович А.Н. Геологическое строение Заволжья между Куйбышевым и Оренбургом // Бюлл.МОИП, отд. геол. 1936. Т. 14. Вып. 6. С. 487–549.
Новиков И.В. Новые данные по проколофонинам СССР // Палеонтол. журн. 1991. № 3. С. 73–85.
Новиков И.В. Новые данные по трематозавроидным лабиринтодонтам Восточной Европы. 3. Qantas samarensis gen. et sp. nov. // Палеонтол. журн. 2012а. № 2. С. 68–77.
Новиков И.В. Новые данные по трематозавроидным лабиринтодонтам Восточной Европы. 4. Род Benthosuchus Efremov, 1937 // Палеонтол. журн. 2012б. № 4. С. 68–79.
Новиков И.В. Лонхоринхиды (Amphibia, Temnospondyli) в нижнем триасе Восточно-Европейской платформы // Проблемы геологии Европейской России: сб. науч. трудов Всерос. науч. конференции, посвящ. 130‑летию со дня рождения проф. Бориса Александровича Можаровского. Саратов: СГТУ, 2013. С. 148–155.
Новиков И.В. Новые темноспондильные амфибии из базального триаса Общего Сырта (Восточная Европа) // Палеонтол. журн. 2016. № 3. С. 88–100.
Новиков И.В. Раннетриасовые амфибии Восточной Европы: эволюция доминантных групп и особенности смены сообществ. М.: РАН, 2018. 358 с.
Новиков И.В., Константинов В.В., Сенников А.Г. и др. Новые уникальные находки раннетриасовых тетрапод на Общем Сырте // Проблемы палеоэкологии и исторической геоэкологии: Сб. трудов Всерос. науч. конференции, посвящ. памяти проф. Виталия Георгиевича Очева / Ред. Иванов А.В., Новиков И.В., Яшков И.А. М.–Саратов: ПИН РАН – СГТУ – ООО “Кузница рекламы”, 2017. С. 70–74.
Очев В.Г. Систематика и филогения капитозавроидных лабиринтодонтов. Саратов: Изд-во Сарат. ун-та, 1966. 184 с.
Очев В.Г. Новые раннетриасовые архозавры с востока Европейской части СССР // Палеонтол. журн. 1979. № 1. С. 104–109.
Сенников А.Г. Новые данные по рауизухидам Восточной Европы // Палеонтол. журн. 1990. № 3. С. 3–16.
Сенников А.Г. Ранние текодонты Восточной Европы. М.: Наука, 1995. 140 с. (Тр. Палеонтол. ин-та РАН. Т. 263).
Сенников А.Г. Открытие примитивной завроптеригии из нижнего триаса Донской Луки и распространение триасовых морских рептилий в России // Палеонтол. журн. 2001. № 3. С. 76–85.
Сенников А.Г. Новая специализированная пролацертилия (Reptilia: Archosauromorpha) из нижнего триаса Оренбургской области // Палеонтол. журн. 2005. № 2. С. 88–97.
Сенников А.Г. Подкласс Archosauromorpha // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 1 / Ред. Ивахненко М.Ф., Курочкин Е.Н. М.: ГЕОС, 2008. С. 266–318.
Сенников А.Г., Новиков И.В. О находках Rhytidosteidae (Amphibia, Temnospondyli) в нижнем триасе Восточной Европы и возможных пищевых адаптациях представителей этого семейства // Палеострат-2012. Годичн. собр. (науч.конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва (Москва, 30 января–1 февраля 2012 г.): тез. докл. / Ред. Алексеев А.С. М.: ПИН РАН, 2012. С. 60–61.
Татаринов Л.П. Новый териодонт из нижнего триаса Оренбургской области // Палеонтол. журн. 1977. № 4. С. 86–90.
Шишкин М.А. Плагиозавры в триасе СССР // Палеонтол. журн. 1967. № 1. С. 92–99.
Шишкин М.А. О реликтах палеозойских архегозавроидов (Amphibia, Temnospondyli) в триасе Еврамерики // Палеонтология и стратиграфия перми и триаса Северной Евразии. IV Всерос. конф., г. Москва, 4–5 апреля 2002 г.: тез. докл. М.: ПИН РАН, 2002. С. 99.
Шишкин М.А. Начальная диверсификация амфибий после терминального пермского кризиса // Палеонтология и стратиграфия перми и триаса Северной Евразии: Матер. V Междунар. конф., посвящ. 150-летию со дня рождения В. П. Амалицкого (1860–1917) (Москва, 22–23 ноября 2010 г.). М.: ПИН РАН, 2010. С. 119–122.
Шишкин М.А. Амфибии-тупилакозавриды (Temnospondyli) в пионерных тетраподных сообществах раннего триаса: происхождение и вопрос об очаге экспансии// Позвоночные палеозоя и мезозоя Евразии: эволюция, смена сообществ, тафономия и палеобиогеография: Матер. конф., посвящ. 80-летию со дня рождения Виталия Георгиевича Очева (1931–2004) (6 декабря 2011 г., Москва). М.: ПИН РАН, 2011. С. 53–57.
Шишкин М.А., Новиков И.В., Фортуни Ж. Новые быстровианидные хрониозухии (Amphibia, Anthracosauromorpha) из триаса России и особенности диверсификации Bystrowianidae // Палеонтол. журн. 2014. № 5. С. 60–71.
Шишкин М.А., Сенников А.Г., Новиков И.В., Ильина Н.В. Дифференциация тетраподных сообществ и некоторые особенности биотических событий в раннем триасе Восточной Европы // Палеонтол. журн. 2006. № 1. С. 3–12.
Huene F. von. Eine Reptilfauna aus der ältesten Trias Nordrusslands // N. Jb. Miner. etc. Abt. B. 1940. Beil.-Bd 84. S. 1–23.
Huene F. von. Ein grosser Pseudosuchier aus der Orenburger Trias // Palaeontogr. Abt. A. 1960. Bd 114. Lf. 1–4. S. 105–111.
Ivakhnenko M.F. Permian and Triassic therocephals (Eutherapsida) of Eastern Europe // Paleontol. J. 2011. V. 45. № 9. P. 981–1144.
Liu J., Xu L., Jia S-H. et al. The Jijuan tetrapod fauna of the Upper Permian of China-2. Stratigraphy, taxonomical review, and correlation // Vertebr. PalAsiat. 2014. V. 52. № 3. P. 328–339.
Lozovsky V.R. Some peculiarities of development of East European and North American platforms during Early Triassic // 28 Intern. Geol. Congr. Abstracts. 1989. V. 2. P. 329–330.
Maryańska T., Shishkin M.A. New Cyclotosaurid (Amphibia: Temnospondyli) from the Middle Triassic of Poland and some problems of interrelationships of capitosauroids // Pr. Muzeum Ziem. 1996. № 43. P. 53–82.
Nield C.M., Damiani R., Warren A. A short-snouted trematosauroid (Tetrapoda, Temnospondyli) from the Early Triassic of Australia: the oldest known trematosaurine // Alcheringa. 2006. № 30. P. 263–271.
Novikov I.V., Shishkin M.A. Triassic Chroniosuchia (Amphibia, Anthracosauromorpha) and evolution of dermal thoracic scutes in the Bystrowianidae // Paleontol. J. 2000. V. 34. Suppl. 2. P. 165–178.
Owen R. On a labyrinthodont amphibian (Rhytidosteus capensis) from the Trias of the Orange Free State, Cape of Good Hope // Quart. J. Geol. Soc. London. 1884. V. 40. P. 333–339.
Shishkin M.A. Patterns of recovery of amphibian diversity in the Triassic // Bull. New Mexico Mus. Natur. Hist. 2007. № 41. P. 369–370.
Shishkin M.A., Rubidge B.S. A relict rhinesuchid (Amphibia: Temnospondyli) from the Lower Triassic of South Africa // Palaeontology. 2000. V. 43. № 4. P. 653–670.
Shishkin M.A., Sulej T. The Early Triassic temnospondyls of the Czatkowice 1 tetrapod assemblage // Palaeontol. Pol. 2009. V. 65. P. 31–77.
Welles S.P. A review of the lonchorhynchine trematosaurs (Labyrinthodontia), and a description of a new genus and species from the Lower Moenkopi Formation of Arizona // PaleoBios. 1993. V. 14. № 3. P. 1–24.
Witzmann F., Schoch R.R., Maisch M.W. A relic basal tetrapod from the Middle Triassic of Germany // Naturwiss. 2008. № 95. S. 67–72.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал