

Палеонтологический журнал, 2020, № 6, стр. 26-36
Динамика развития ринхонеллид (Brachiopoda) в девоне и раннем карбоне Закавказья
А. В. Пахневич *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: alvpb@mail.ru
Поступила в редакцию 21.11.2019
После доработки 23.12.2019
Принята к публикации 26.12.2019
Аннотация
Охарактеризованы комплексы видов девонских и раннекарбоновых брахиопод из отряда Rhynchonellida Закавказья (Нахичеванская АР Азербайджана и Армения). Выявлены смены доминирующих надсемейств и экологических типов ринхонеллид. В девоне ринхонеллиды многочисленны. В раннем и среднем девоне преобладают представители надсемейства Uncinuloidea. Их реликтовые формы доживают до начала фамена. В раннем и среднем девоне доминируют свободнолежащие и якорные каверновые ринхонеллиды. Последний экологический подтип выделен впервые. Смена преобладающих таксономических и экологических групп ринхонеллид происходит на границе франского и фаменского веков. Но Rhynchonellida остаются многочисленными компонентами брахиоподовой фауны. На границе фаменского и турнейского веков происходит уменьшение разнообразия этих брахиопод, но не выявлено смены доминирующих во фране и фамене надсемейства Rhynchotrematoidea и экологического типа якорных ринхонеллид. Роль брахиопод отряда Rhynchonellida в брахиоподовой фауне раннего карбона незначительная. Причина падения разнообразия в пределах некоторых зон, вероятно, связана с тафономическими условиями.
ВВЕДЕНИЕ
В Нахичеванской автономной республике Азербайджана и ближайших районах Армении располагаются важные с точки зрения фаунистики и стратиграфии разрезы нижнего девона–нижнего карбона. Один из таких разрезов находится на склоне горы Геран-Каласы (Гречишникова, Левицкий, 2011). Он является одним из наиболее полных разрезов интервала верхний девон–нижний карбон, сложенных непрерывными морскими отложениям. Разрезы Закавказья содержат богатую разнообразную фауну, среди которой в девонских отложениях встречаются: фораминиферы, строматопораты, хететиды, гелиолитиды, табуляты, ругозы, брюхоногие, двустворчатые и головоногие моллюски, в том числе, наутилоидеи, тентакулиты, трилобиты, остракоды, мшанки, конодонты, криноидеи, рыбы, а также водоросли. В каменноугольных отложениях – фораминиферы, табуляты, остракоды, криноидеи, конодонты, а среди растений – водоросли. И в тех, и в других отложениях одной из наиболее многочисленных групп являются брахиоподы. Они доминировали в морях, как девона, так и карбона, но преобладали при этом различные отряды. Ринхонеллиды известны в отложениях почти всех зон (приняты по: Grechishnikova, 2018). Первое описание комплексов видов ринхонеллид опубликовала Г.Т. Мириева (2010). Однако присутствие некоторых видов ринхонеллид спорно, а отдельные комплексы неполные. Их подробное описание дано нами (Pakhnevich, 2018).
Целью настоящей работы является выявление особенностей динамики фауны брахиопод отряда Rhynchonellida, доминирующих таксономических и экологических групп ринхонеллид в девоне и раннем карбоне Закавказья, в том числе, в периоды массовых вымираний.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Зональные комплексы ринхонеллид девона и карбона
Самые древние девонские отложения Закавказья относятся к эмскому ярусу нижнего девона (табл. 1). Они выделены в зону Arduspirifer extensis, в которой ископаемая фауна немногочисленна. Ринхонеллиды здесь не встречены. Возможно, это связано с привносом большого количества терригенного материала, о чем можно судить по отложениям в верхах зоны. Или же слой частично размыт и подвергся выветриванию, признаки этого были найдены.
Таблица 1.
Зональная схема нижнего девона–нижнего карбона Закавказья
| Система | Отдел | Ярус | Брахиоподовые зоны |
|---|---|---|---|
| Каменноугольная | Нижний | Визейский | Moderatoproductus moderatus |
| –* | |||
| Турнейский | Spirifer baiani–Marginatia burlingtonensis | ||
| Rhipidomella michelini | |||
| Parallelora praeulbanensis–Rhytiophora curtirostris | |||
| Девонская | Верхний | Фаменский | Sphenospira julii–Spinocarinifera nigra |
| Paurogastroderhynchus nalivkini | |||
| Cyrtospirifer pamiricus–Enchondrospirifer ghorensis | |||
| Dmitria seminoi | |||
| Cyrtospirifer asiaticus–Mesoplica meisteri | |||
| Франский | Uchtospirifer subarchiaci–Cyphoterorhynchus arpaensis | ||
| Adolfia zickzack | |||
| Средний | Живетский | Indospirifer pseudowilliamsi | |
| Stringocephalus burtini | |||
| Эйфельский | Mucrospirifer diluvianoides–Radiomena irregularis | ||
| Alatiformia araxica–Dagnachonetes caucasius | |||
| Arduspirifer intermedius | |||
| Zdimir pseudobaschkiricus–Megastrophia uralensis | |||
| Нижний | Эмский | ||
| Arduspirifer extensus |
Первые ринхонеллиды в девоне Закавказья известны из отложений зоны Zdimir pseudobaschkiricus–Megastrophia uralensis. Внутри зоны Zdimir pseudobaschkiricus–Megastrophia uralensis проходит граница нижнего и среднего девона, эмса и эйфеля. Здесь ринхонеллиды малочисленны, присутствуют всего шесть видов: Septalaria subtetragona (Schnur, 1851) (рис. 1, а–д), Schnurella transversa (Reed, 1908), Glosshypothyridina procuboides (Kayser, 1871), Beckmannia pentagona (Kayser, 1871), Isopoma brachyptycta (Schnur, 1853), Kransia sp. (рис. 3, а–д). Зональный вид Uncinulus keltibericus Schumann, 1965, указанный в стратиграфической схеме М.А. Ржонсницкой и А.Б. Мамедовым (Rzhonsnitskaya, Mamedov, 2000), не обнаружен. Наиболее важным для этих отложений видом является S. subtetragona. Ринхонеллиды принадлежат к надсемействам: Uncinuloidea, Pugnacoidea и Camarotoechioidea. Доминируют представители унцинулоидей. Среди ринхонеллид встречаются брахиоподы из разных экологических типов, выделенных Е.А. Ивановой (1949). Преобладают ринхонеллиды свободнолежащего экологического типа, у которых во взрослом состоянии ножка атрофировалась, и тогда они переходили к свободному лежанию на дне, используя для поднятия над субстратом синус или седло, или высокий передний край. К этой группе относятся брахиоподы S. transversa и G. procuboides. S. subtetragona и I. brachyptycta принадлежали экологической группе якорных брахиопод, то есть прикреплялись ножкой. Вероятно, такой же образ жизни вели и мелкораковинные Uncinuloidea – B. pentagona (рис. 4, е–к) и Kransia sp., но они представляли особый экологический подтип якорных брахиопод. Эти брахиоподы были адаптированы к поселению в кавернах субстрата, возможно среди кораллов. Это достигалось за счет уменьшения в размерах. Такая адаптация известна и для современных брахиопод, и для ископаемых (Pakhnevich, 2009), например, у современных родов: Argyrotheca Dall, 1900, Megathyris Orbigny, 1847 и т.д. Все перечисленные девонские виды продолжают обитать в данном бассейне и в более позднее время, за исключением специфичных для зоны S. subtetragona и Kransia sp. Последний вид имеет необычную для рода Kransia Westbroek, 1968 форму раковины, но по внутреннему строению полностью соответствует ему (рис. 3, а–д). Он интересен тем, что является наиболее ранним представителем рода. Большинство видов рода Kransia известно из более поздних отложений среднего девона.
Рис. 1.
Характерные девонские виды ринхонеллид Закавказья: а–д – Septalaria subtetragona (Schnur, 1851), раковина, экз. ПИН, № 3744/850; трехмерная модель (×1.5): а – вид с брюшной створки, б – вид сбоку, в – вид со спинной створки, г – вид со стороны замочного края, д – вид со стороны переднего края; р-н горы Дагна, обн. 1710; верхний эмс – нижний эйфель, зона Zdimir pseudobaschkiricus – Megastrophia uralensis; е–к – Oligoptycherhynchus daleidensis (Roemer, 1844), раковина, экз. ПИН, № 3744/823 (×2): е – вид с брюшной створки, ж – вид со спинной створки, з – вид со стороны переднего края, и – вид со стороны замочного края, к – вид сбоку; левый берег р. Арпы у с. Данзик (Нахичеванская АР), обн. 19, сл. 29; средний эйфель, зона Alatiformia araxica – Dagnachonetes caucasius; л–п – Paropamisorhynchus kotalensis (Brice, 1970), раковина, экз. ПИН, № 3744/828 (×2): л – вид с брюшной створки, м – вид со спинной створки, н – вид сбоку, о – вид со стороны замочного края, п – вид со стороны переднего края; р-н горы Кабахдаг (Нахичеванская АР), обн. 1044а; верхний фамен, зона Paurogastroderhynchus nalivkini; р–ф – Sartenaerus letiensis (Gosselet, 1887), раковина, экз. ПИН, № 3744/826 (×2): р – вид с брюшной створки, с – вид со спинной створки, т – вид сбоку, у – вид со стороны переднего края, ф – вид со стороны замочного края; правый берег р. Арпы, напротив с. Данзик (Нахичеванская АР), обн. 36, сл. 3; верхний фамен, зона Cyrtospirifer pamiricus–Enchondrospirifer ghorensis.

В зоне Arduspirifer intermedius не встречено ни одного вида ринхонеллид. Пробелы в распространении ринхонеллид, скорее всего, имеют тафономическую причину, но не являются периодами резкого обеднения брахиоподовой фауны. С другой стороны, и прочая фауна зоны немногочисленна. И это может быть признаком снижения биоразнообразия за счет формирования неблагоприятных условий обитания, например, увеличение терригенного сноса с суши.
Для зоны Alatiformia araxica – Dagnachonetes caucasius характерны 7 видов: Oligoptycherhynchus daleidensis (Roemer, 1844), Uncinulus subwilsoni (Orbigny, 1850), Beckmannia angularis (Phillips, 1841), B. pentagona, Solidipontirostrum sp., Nymphorhynchia sp., Kransia parallelepipeda (Bronn, 1837). По количеству видов преобладающим остается надсемейство Uncinuloidea, а Pugnacoidea уходят на третий план, освобождая место, появившимся в данном бассейне представителям надсемейства Rhynchotrematoidea. Из последнего надсемейства наиболее многочисленными являются брахиоподы вида O. daleidensis (рис. 1, е–к). Он может быть использован в качестве зонального. Среди представителей семейства Uncinuloidea становятся все более многочисленными мелкораковинные формы родов Kransia и Beckmannia Mohanti, 1972. Возможно, это связано с распространением рифовых экосистем. К крупным якорным формам относятся O. daleidensis и Nymphorhynchia sp. Вероятнее всего, Solidipontirostrum sp., также как U. subwilsoni, принадлежал к свободнолежащим ринхонеллидам. Возможно, на ювенильной стадии развития Solidipontirostrum sp. прикреплялся к субстрату ножкой, но с увеличением раковины ножка уже не могла выполнять функцию прикрепления, атрофировалась, и брахиопода свободно лежала на дне, возможно, частично погружаясь в донный осадок. Несмотря на обилие слоев с терригенными породами, И.А. Гречишниковой (Gre-chishnikova, 2018) отмечается высокое биоразнообразие фауны.
Зона среднего девона Mucrospirifer diluvianoides–Radiomena irregularis соответствует верхам эйфеля. Для нее характерно самое высокое родовое и видовое разнообразие ринхонеллид в среднем девоне. Это 15 видов: Kransia parallelepipeda, K. praecedens (Kulkov, 1960), K. subcordiformis (Schnur, 1853), Primipilaria primipilaris (Buch, 1834), Beckmannia pentagona, B. minor (Schnur, 1853), Pseudocamarophoria undulataeformis Biernat, 1966, Pugnax praevius Schmidt, 1941, Isopoma brachyptycta, Camerophorina pachyderma (Quenstedt, 1871), Schnurella transversa, Glosshypothyridina procuboides, Uncinulus korovini Khalfin, 1937, Corvinopugnax sp., Mirantesia sp. Здесь вновь преобладают ринхонеллиды надсемейства Uncinuloidea, также есть представители надсемейств Camarotoechioidea, Pugnacoidea. Субдоминантами становятся представители надсемейства Camarotoechioidea. Ни одна из этих групп не остается устойчиво на уровне субдоминантной на протяжении среднего девона Закавказья. Наиболее многочисленными в этой зоне являются виды K. parallelepipeda и B. pentagona. Представители родов Kransia, Primipilaria Struve, 1992, Beckmannia преобладают в фауне ринхонеллид. Они формируют экологическую группу якорных каверновых брахиопод. Это хорошо сочетается с развитием в это время на территории Закавказья биогермов (Grechishnikova, 2018). К экологическому типу свободнолежащих брахиопод относятся: P. praevius, S. transversa, G. procuboides, U. korovini, Corvinopugnax sp., Mirantesia sp. И лишь редко встречающиеся P. undulataeformis, I. brachyptycta и C. pachyderma принадлежали к основному якорному типу.
Отложения зоны Stringocephalus burtini нижнего живета бедны ринхонеллидами. Это виды унцинулоидей Kransia parallelepipeda и Glosshypothyridina procuboides. Первый относится к якорному каверновому экологическому подтипу, второй – к свободнолежащему. Причины падения разнообразия непонятны, потому что отложения этой зоны наполнены разнообразными ископаемыми остатками беспозвоночных животных, вплоть до образования органогенных известняков.
В позднем живете, зона Indospirifer pseudowilliamsi, разнообразие ринхонеллид увеличивается, несмотря на возрастание терригенного материала в отложениях. Так же, как и в предыдущих слоях, преобладают представители надсемейства Uncinuloidea. Наряду с ними встречаются ринхонеллиды из надсемейств Pugnacoidea и, вновь появившиеся, Rhynchotrematoidea. Для зоны верхнего живета Indospirifer pseudowilliamsi характерны виды: Uncinulus korovini, Kransia parallelepipeda, K. subcordiformis, Beckmannia minor, Isopoma brachyptycta, Schnurella transversa, Pugnax praevius, Glosshypothyridina procuboides, Ripidiorhynchus sp. В этих отложениях появляется первый представитель рода Ripidiorhynchus Sartenaer, 1966, который чаще встречается в верхнем девоне и является элементом фауны франа (рис. 3, е–к). Наиболее часто встречающиеся роды, найденные как в верхах эмса, так и в верхах живета, – унцинулоидеи Beckmannia и Kransia. Вид B. pentagona прослеживается с перерывом от верхов эмса до верхов живета, а вид K. parallelepipeda – на протяжении всего эйфеля и живета. Значительных смен видов и родов ринхонеллид на границе эйфеля и живета не наблюдается. Опять же, преобладают две экологические группы – якорные каверновые брахиоподы (K. parallelepipeda, K. subcordiformis, B. minor) и свободнолежащие (U. korovini, S. transversa, P. praevius, G. procuboides). Единичны представители основного якорного типа I. brachyptycta и Ripidiorhynchus sp.
В начале верхнего девона меняются доминирующие группы ринхонеллид. На смену Uncinuloidea в качестве доминантов появляются ринхонеллиды из семейства Trigonirhynchiidae (Rhynchotrematoidea). В отложениях первой половины франского яруса видовой состав ринхонеллид обеднен. Для зоны Adolphia zickzack характерны виды: Ripidiorhynchus gnishikensis (Abramian, 1959), Cyphoterorhynchus arpaensis (Abramian, 1957), Paropamisorhynchus kotalensis (Brice, 1970), Porthmorhynchus ferquensis (Gosselet, 1887), Pugnax acuminatus (Sowerby, 1822), Coeloterorhynchus sp., Zezinia multicostata Pakhnevich, 2018 (рис. 3, л–о), Septalariidae gen. et sp. indet. Интересны два последних вида, которые являются продолжением ранне-среднедевонской фауны ринхонеллид, исчезающей в живете (подсем. Betterbergiinae) и фране. Интересно, что по таким группам фауны, как ругозы, гелиолитиды, конодонты также прослеживается наличие среднедевонских элементов (Grechishnikova, 2018). Возможно, данную акваторию следует рассматривать как рефугиум живетской фауны, но не стоит относить ее к живету, поскольку доминирующая фауна ринхонеллид имеет верхнедевонский облик. В ней преобладают представители надсемейства Rhynchotrematoidea, также присутствуют роды надсемейств Pugnacoidea и единичные представители надсемейств Uncinuloidea и Camarotoechioidea. В конце первой половины франа виды надсемейства Uncinuloidea полностью исчезают. Меняются и экологические доминирующие группы. Полностью исчезает экологический тип прикрепленных каверновых брахиопод. Из свободнолежащих представлены единичные P. acuminatus, Z. multicostata, Septalariidae gen. et sp. indet., Coeloterorhynchus sp. Остальные ринхонеллиды (R. gnishikensis, C. arpaensis, P. kotalensis, P. ferquensis), доминирующие в фауне, принадлежали к основному якорному типу.
Зона Cyrtospirifer subarchiaci–Cyphoterorhynchus arpaensis верхнего девона, франского яруса характеризуется видами: Ripidiorhynchus gnishikensis, Cyphoterorhynchus arpaensis, Paropamisorhynchus kotalensis, Porthmorhynchus ferquensis. Наиболее многочисленным становится C. arpaensis (рис. 2, а–д). Он является зональным видом. Полностью исчезают остатки живетской фауны. Представлены только виды семейства Trigonirhynchiidae (Rhynchotrematoidea). Во время, соответствующее формированию данной зоны, происходит падение биоразнообразия, как на уровне семейств, так и родов, и видов. Все брахиоподы принадлежали к основному якорному экологическому типу.
Рис. 2.
Зональные виды ринхонеллид Закавказья: а–д – Cyphoterorhynchus arpaensis (Abramian, 1957), раковина, экз. ПИН, № 3744/832 (×2): а – вид с брюшной створки, б – вид со спинной створки, в – вид сбоку, г – вид со стороны переднего края, д – вид со стороны замочного края; р-н с. Данзик (Нахичеванская АР), обн. 101, сл. 14; верхний фран, зона Cyrtospirifer subarchiaci – Cyphoterorhynchus arpaensis; е–к –Paurogastroderhynchus nalivkini (Abramian, 1957), раковина, экз. ПИН, № 3744/800 (×1.5): е – вид с брюшной створки, ж – вид со спинной створки, з – вид сбоку, и – вид со стороны замочного края, к – вид со стороны переднего края; р-н с. Данзик, обн. 15, сл. 22; верхний фамен, зона Paurogastroderhynchus nalivkini.

Рис. 3.
Необычные девонские представители отряда Rhynchonellida: а–д – Kransia sp., раковина, экз. ПИН, № 3744/844, трехмерная модель (×3): а – вид с брюшной створки, б – вид со спинной створки, в – вид сбоку, г – вид со стороны переднего края, д – вид со стороны замочного края; юж. склон горы Велидаг (Нахичеванская АР), обн. 48, сл. 2; верхний эмс – нижний эйфель, зона Zdimir pseudobaschkiricus – Megastrophia uralensis; е–к – Ripidiorhynchus sp., раковина, экз. ПИН, № 3744/936 (×1): е – вид с брюшной створки, ж – вид со спинной створки, з – вид сбоку, и – вид со стороны переднего края, к – вид со стороны замочного края; район обогатительной фабрики пос. Гюмушлуг (Нахичеванская АР), обн. 1155; верхний живет, зона Indospirifer pseudowilliamsi; л–о – Zezinia multicostata Pakhnevich, 2018, раковина, экз. ПИН, № 3744/846 (×1.5): л – вид с брюшной створки, м – вид со стороны переднего края, н – вид со стороны замочного края, о – вид сбоку; прав. берег р. Джаанам-Дереси, в 1.5 км восточнее горы Тежгар (Нахичеванская АР), обн. 601; нижний фран, зона Adolfia zickzack.
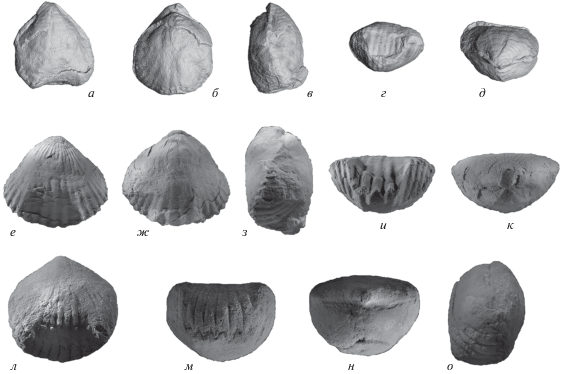
Первая фаменская зона Cyrtospirifer asiaticus–Mesoplica meisteri отличается увеличением разнообразия ринхонеллид. В ее отложениях встречены виды: Paropamisorhynchus kotalensis, Stenaulacorhynchus sp., Sharovaella mirabilis Pakhnevich, 2012, Sartenaerus letiensis (Gosselet, 1887), S. charakensis (Brice, 1967), Greira transcaucasica O. Erlanger, 1993, Sinotectirostrum zobeida (Nalivkin, 1937), Gesoriacorostrum cf. boloniensis (Orbigny, 1850) и Ptychomaletoechia sp. Все виды принадлежат надсемейству Rhynchotrematoidea, за исключением Stenaulacorhynchus sp., который относится к надсемейству Camarotoechioidea. Впервые в этих слоях встречаются пористые ринхонеллиды, представленные сразу двумя родами и видами. Они принадлежат к семейству Trigonirhynchiidae. Появление ринхонеллид с пористой стенкой раковины является признаком сформировавшихся новых условий обитания в начале фамена, поскольку упомянутый признак характеризует появление новой физиологической особенности, связанной с формированием эндопор.
Зона Dmitria seminoi фамена плохо охарактеризована комплексом ринхонеллид. Отмечены виды Sartenaerus letiensis, Sharovaella mirabilis, Araratella dichotomians (Abramian, 1954). Преобладают представители семейства Trigonirhynchiidae. Большая часть ринхонеллид относятся к пористым. Впервые пористость появляется сразу в разных семействах, помимо тригониринхид она отмечена у A. dichotomians из семейства Septalariidae (Camarotoechioidea). Все ринхонеллиды относятся к якорному экологическому типу.
Следующая зона, относящаяся к низам верхнего фамена, Cyrtospirifer pamiricus–Enchondrospirifer ghorensis, характеризуется видами: Sartenaerus charakensis, S. letiensis, Paropamisorhynchus kotalensis, Sharovaella mirabilis, Zaigunrostrum nakhichevanensis Pakhnevich, 2018, Araratella dichotomians, Navalicria sp., Stenaulacorhynchus sp., Platyterorhynchinae gen et sp. indet. Это наиболее богатая видами зона фамена. Доминирующая группа ринхонеллид снова не меняется, но присутствуют еще и представители надсемейства Camarotoechioidea. Здесь они наиболее многочисленны из всех зон верхнего девона. Вид S. letiensis является наиболее часто встречающимся в отложениях этой, предыдущий и последующей зон (рис. 1, р–ф). Все виды принадлежат к экологической группе якорных брахиопод, за исключением Navalicria sp. Если в нижнем и среднем девоне, а также фране Закавказья свободнолежащие ринхонеллиды относились к надсемействам Uncinuloidea и Pugnacoidea, то в фамене к этому экологическому типу принадлежали единичные виды надсемейств Camarotoechioidea (в отложениях этой зоны – Navalicria sp.), Rhynchotrematoidea и Pugnacoidea.
В отложениях зоны Paurogastroderhynchus nalivkini верхнего фамена обнаружены ринхонеллиды: Paurogastroderhynchus nalivkini (Abramian, 1957), Araratella dichotomians, Sartenaerus charakensis, S. letiensis, Sharovaella mirabilis, Zaigunrostrum nakhichevanensis, Gesoriacorostrum cf. boloniensis, Paropamisorhynchus kotalensis, Hypseloterorhynchus sp., Sinotectirostrum sp. Вид P. nalivkini является зональным (рис. 2, е–к). Вид P. kotalensis – единственный перешедший из франа в фамен (рис. 1, л–п). Он встречается вплоть до конца зоны Paurogastroderhynchus nalivkini, но в отложениях более поздних зон не обнаружен. Помимо ринхотрематоидей, в фауне присутствует по одному представителю семейств Pugnacidae (Pugnacoidea) и Septalariidae (Camarotoechioidea). Впервые свободнолежащие ринхонеллиды появляются в семействе Trigonirhynchiidae (Rhynchotrematoidea), это вид P. nalivkini. Вновь появляются свободнолежащие брахиоподы из надсемейства Pugnacoidea – Hypseloterorhynchus sp. Остальные виды принадлежат к якорному экологическому типу.
Зона Sphenospira julii–Spinocarinifera nigra верхнего фамена характеризуется видами: Araratella dichotomians, Tchanakhtchirostrum araraticum (Abramian, 1957), Sharovaella mirabilis, Zaigunrostrum nakhichevanensis, Ptychomaletoechia panderi (Semenov et Möller, 1864), Sinotectirostrum delicatacostata (Abramian, 1957), Leptocaryorhynchus sp. В самом конце фамена происходит уменьшение биоразнообразия ринхонеллид. Из предыдущих зон в упомянутую переходят только пористые ринхонеллиды. Здесь появляется еще одна форма с пористой раковиной, T. araraticum, также принадлежащая ринхотрематоидеям. Наиболее многочисленными также являются пористые ринхонеллиды видов A. dichotomians, T. araraticum, S. mirabilis. Доминирующем остается надсемейство Rhynchotrematoidea. Встречаются и представители надсемейства Camarotoechioidea. Все ринхонеллиды относятся к якорному экологическому типу. Вполне возможно, что мелкие представители, P. panderi (рис. 4, р–ф) и Leptocaryorhynchus sp. (рис. 4, м–п) могут относиться к якорному каверновому экологическому подтипу.
Рис. 4.
Некоторые представители якорного кавернового экологического подтипа ринхонеллид: а–д – Kransia parallelepipeda (Bronn, 1837), раковина, экз. ПИН, № 3744/825 (×2): а – вид с брюшной створки, б – вид со спинной створки, в – вид со стороны переднего края, г – вид со стороны замочного края, д – вид сбоку; вост. склон горы Касан-Гулу-Бах (Нахичеванская АР), обн. 63, сл. 6; живет, зона Stringocephalus burtini; е–к – Beckmannia pentagona (Kayser, 1871), раковина, экз. ПИН, № 3744/827 (×2): е – вид с брюшной створки, ж – вид со спинной створки, з – вид со стороны переднего края, и – вид со стороны замочного края, к – вид сбоку; лев. берег р. Джаанам-Дереси, в 1.5 км. западнее горы Казма (Нахичеванская АР), обн. 57, сл. 9; верхний эйфель, зона Mucrospirifer diluvianoides – Radiomena irregularis; л–п – Leptocaryorhynchus sp., раковина, экз. ПИН, № 3744/849, трехмерная модель (×2): л – вид с брюшной створки, м – вид со спинной створки, н – вид сбоку, о – вид со стороны переднего края, п – вид со стороны замочного края; восточнее горы Геран-Каласы (Нахичеванская АР), обн. 10, сл. 3; верхний фамен, зона Sphenospira julii – Spinocarinifera nigra; р–ф – Ptychomaletoechia panderi (Sem. et Möl., 1864), раковина, экз. ПИН, № 3744/841 (×2): р – вид с брюшной створки, с – вид со спинной створки, т – вид сбоку, у – вид со стороны замочного края, ф – вид со стороны переднего края; бассейн р. Джаанам-Дереси (Нахичеванская АР), обн. 1412 б; верхний фамен, зона Sphenospira julii – Spinocarinifera nigra; х–э – Hemiplethorhynchus sp. 1, раковина, экз. ПИН, № 3744/851, трехмерная модель (×2): х – вид с брюшной створки, ч – вид со спинной створки, ш – вид сбоку, щ – вид со стороны переднего края, э – вид со стороны замочного края; гора Геран-Каласы (Нахичеванская АР), обн. 10, сл. 6; нижний карбон, нижний турне, зона Unispirifer praeulbanensis – Rhytiophora curtirostris.

Разнообразие ринхонеллид в нижнем карбоне резко падает. Из фамена переходят только два вида Ptychomaletoechia panderi и Tchanakhtchirostrum araraticum. Кроме T. araraticum, полностью исчезают пористые ринхонеллиды. Первая зона нижнего карбона Unispirifer praeulbanensis–Rhytiophora curtirostris характеризуется видами надсемейства Rhynchotrematoidea: T. araraticum, P. panderi и Hemiplethorhynchus sp. 1 (рис. 4, р–ф); все относятся к якорному экологическому типу, возможно, два последних вида – к каверновому подтипу.
Два вида известны из отложений зоны Spirifer baiani–Marginatia burlingtonensis (верхний турне): Hemiplethorhynchus sp. 2 и Wellerellinae gen. et sp. indet. Последний вид является представителем нового для Закавказья надсемейства ринхонеллид Wellerelloidea. Оба вида относятся к якорному экологическому типу.
Новый экологический подтип брахиопод
Некоторые современные брахиоподы встречаются на коралловых рифах и образуют там большие скопления. Так около Бермудских о-вов на рифах обитает теребратулида Argyrotheca bermudana Dall, 1911. Она образует скопления от 85 до 3595 экз/м2 (Logan, 1975). Все представители рода Argyrotheca – микроморфные брахиоподы. У северного побережья Ямайки другой вид, Argyrotheca sp., является одним из доминантов в рифовом сообществе, образуя скопления до 100 экз/м2 (Jackson et al., 1971). Эти брахиоподы, имея мелкие размеры раковины, прекрасно себя чувствуют в кавернах коралловых построек. В кавернах подводных пещер живут и другие современные микроморфные брахиоподы: Tethyrhynchia mediterranea Logan, 1994 (отряд Rhynchonellida, остальные виды относятся к отряду Terebratulida), Megathyris detruncata (Gmelin, 1790), Argyrotheca cuneata (Risso, 1826), A. cistellula (Searles-Wood, 1841), Joania cordata (Risso, 1826) (Logan, Zibrowius, 1994; Simon, Willems, 1999; Ruggiero, 2000; Saito et al., 2000). Хорошо себя чувствует на коралловых постройках Platidia anomioides (Scacchi et Philippi, 1844). Этот вид найден на банке Брукер (Средиземное море) на глубинах 97 и 160 м (Маккавеева, 1963). Но не только каверны в коралловых колониях пригодны для обитания P. anomioides. Вид найден в кавернах толеитового базальта на глубине 730–970 м в проливе Брансфилд близ Западного Антарктического полуострова на подводном вулкане Орка (Зезина, Пахневич, 2000). Однако не всегда появление брахиопод на коралловых постройках приводит к измельчанию раковины как адаптации для обитания в кавернозном субстрате. Крайний случай измельчания брахиопод в процессе адаптации к обитанию в мелких элементах субстрата – современная теребратулида Gwynia capsula (Jeffreys, 1859), которая принадлежит к интерстициальной мейофауне и может обитать между песчинок субстрата (Swedmark, 1964).
В морях Северной Атлантики в зарослях мадрепорового коралла Lophelia prolifera (Pallas, 1766) характерным видом является брахиопода Macandrevia cranium (Müller, 1776) (Филатова, 1938). Несмотря на обитание на колониях кораллов, ни замедления роста, ни укорочения онтогенеза, что привело бы к измельчанию раковины, не происходит. То есть перечисленные мелкораковинные брахиоподы специально адаптировались к обитанию в кавернозном субстрате. Поэтому следует выделить отдельный новый экологический подтип якорного типа брахиопод – якорные каверновые. Он не соответствует экологическим группам, выделенным Е.А. Ивановой, поскольку принципы их выделения отличаются.
К подобным брахиоподам отнесен и Microsphaeridiorhynchus sp. из силура [лудловский ярус, формация Хамра (Hamra Formation), местонахождение Банквят 1 (Bankvat 1)] Швеции (о-в Готланд). Эти микроморфные брахиоподы (максимальная длина раковины 4.5–6.5 мм) встречались в местонахождении наряду с крупнораковинными брахиоподами. Вид доминировал в сообществе. Здесь же были найдены кораллы: ругозы и табуляты (Pakhnevich, 2009). Вероятно, брахиоподы Microsphaeridiorhynchus sp. селились на постройках кораллов и относились к якорному каверновому подтипу.
Вероятно, к таким же якорным каверновым брахиоподам принадлежали и упомянутые здесь Kransia sp., K. parallelepipeda, K. praecedens, K. subcordiformis, P. primipilaris, B. pentagona, B. minor, B. angularis, P. panderi, Leptocaryorhynchus sp. и Hemiplethorhynchus sp. 1 и 2. Вместе с ними встречаются строматопораты, табуляты, ругозы, мшанки, водоросли, которые являются рифостроителями (Большакова и др., 1997), и на их постройках, и в кавернах могли жить эти брахиоподы.
Основные закономерности смены таксономических и экологических групп ринхонеллид
Таким образом, изучая ринхонеллид с верхнего эмса до верхнего турне, удалось установить, что доминантами верхов нижнего и среднего девона, как по количеству родов и видов, так и экземпляров, были брахиоподы надсемейства Uncinuloidea, в основном семейства Hebetoechiidae. В сопутствующей фауне присутствовали представители еще трех надсемейств ринхонеллид. На границе эйфеля и живета не происходит какой-либо значимой смены фаун. Среди ринхонеллид верхов нижнего и среднего девона преобладают свободнолежащие животные – это представители надсемейств Uncinuloidea и Pugnacoidea. Для этого времени характерен и выделенный экологический тип – якорные каверновые брахиоподы.
На границе живета и франа происходит полная перестройка сообществ ринхонеллид. Во фране сохраняются некоторые единичные ранне- и среднедевонские представители Uncinuloidea. Среднедевонские элементы фауны отмечены и в других группах франских беспозвоночных. Вероятнее всего, франские моря Закавказья служили рефугиумом для последних ранне- и среднедевонских представителей фауны, в том числе, и для ринхонеллид. Несмотря на присутствие реликтов, экологический облик брахиопод меняется – начинают преобладать якорные брахиоподы. Последние потомки среднедевонской фауны исчезают после первой половины франа. Доминантами становятся представители надсемейства Rhynchotrematoidea, семейства Trigonirhynchiidae. Из сопутствующих ринхонеллид почти исчезают роды надсемейства Pugnacoidea. Наиболее многочисленными во франских отложениях являются представители вида C. arpaensis. В фаменских отложениях, за исключением самых верхов, преобладают ринхонеллиды рода Sartenaerus Özdikmen, 2008. Разнообразие видов падает на границе франа и фамена (почти в 2 раза), за счет вымирания большинства франских родов семейства Trigonirhynchiidae (Rhynchotrematoidea). Среди новых родов тригониринхиид появляются брахиоподы с пористой раковиной. Это событие характеризует данную границу и приурочено к изменениям условий обитания в рассматриваемом бассейне. Скорее всего, оно связано с недостатком кислорода в воде (Пахневич, 2015). Возможно, поры выполняли функцию увеличения газообмена. Параллельно пористость возникает и у представителей надсемейства Camarotoechioidea. Но пористые камаротоэхиодеи имеют более широкое географическое распространение, чем тригониринхииды. Пористые ринхонеллиды становятся наиболее многочисленными на границе девона и карбона. В карбон переходит единственный вид T. araraticum. Из непористых девонских брахиопод до карбона доживает P. panderi. Этот вид, а также Leptocaryorhynchus sp. и карбоновые Hemiplethorhynchus sp., возможно, снова занимают экологическую нишу брахиопод, обитающих в кавернах субстрата. Помимо этого экологического подтипа, в фамене вновь появляются свободнолежащие брахиоподы, но они немногочисленны и принадлежат к надсемействам Pugnacoidea и Rhynchotrematoidea. Однако доминируют во фране, фамене и турне брахиоподы якорного экологического типа. В начале нижнего карбона отмечается падение биоразнообразия ринхонеллид. Они становятся редки и не играют значительной роли в брахиоподовой фауне раннего карбона. За весь промежуток рассмотренной геологической истории Закавказья доминантами являются представители только двух надсемейств, Uncinuloidea и Rhynchotrematoidea, которые сменяют друг друга. Ринхонеллиды надсемейств Pugnacoidea и Camarotoechioidea также присутствуют, но никогда не переходят в ранг доминирующих таксонов.
Причин уменьшения разнообразия ринхонеллид в отложениях зон Arduspirifer extensis, Arduspirifer intermedius, Stringocephalus burtini и Dmitria seminoi может быть несколько. Одна из возможных причин – увеличение поступления терригенного материала с суши. Но, судя по литологическим характеристикам зон, где ринхонеллиды были многочисленны, значительный терригенный материал поступал в море и при их большом разнообразии. Возможно, это связано с тафономическими условиями формирования отложений, при которых раковины брахиопод разрушались, что не отражает подлинное разнообразие.
Список литературы
Большакова Л.Н., Геккер М.Р., Горюнова Р.В. и др. Рифогенные постройки в палеозое России. М.: Наука, 1997. 157 с.
Гречишникова И.А., Левицкий Е.С. Опорный разрез фамена и нижнего карбона Геран-Каласи (Нахичеванская АР, Азербайджан) // Стратигр. Геол. корреляция. 2011. Т. 19. № 1. С. 24–46.
Зезина О.Н., Пахневич А.В. О нахождении современных замковых брахиопод в районе подводной вулканической активности Антарктического полуострова // Докл. Акад. наук. 2000. Т. 370. № 3. С. 397–398.
Иванова Е.А. Условия существования, образ жизни и историческое развитие некоторых брахиопод среднего и верхнего карбона Подмосковной котловины. М.: Изд-во АН СССР, 1949. 152 с. (Тр. ПИН АН СССР. Т. 21).
Маккавеева Е.Б. Население некоторых банок Эгейского моря // Тр. Севастопольской биол. станции. 1963. Т. 16. С. 211–214.
Мириева Г.Т. Зональное расчленение отложений верхнего девона Южного Закавказья (Нахчыванской АР) по ринхонеллидам // Эволюция органического мира и биотические кризисы. Матер. LVI сессии Палеонтол. об-ва при РАН. СПб., 2010. С. 74–76.
Пахневич А.В. О появлении пористости раковины у позднедевонских брахиопод отряда Rhynchonellida // Матер. конф. “Морфогенез в индивидуальном и историческом развитии: устойчивость и вариабельность” (21–23 апреля 2015 г., Москва). М.: ПИН РАН, 2015. С. 50–51.
Филатова З.А. Количественный учет донной фауны юго-западной части Баренцева моря // Тр. ПИНРО. 1938. Вып. 2. С. 3–58.
Grechishnikova I.A. Biostratigraphy of the Devonian and Carboniferous of Transcaucasia (Brachiopod Zonal Scale) // Paleontol. J. 2018. V. 52. № 8. P. 832–862.
Jackson J.B.C., Goreau T.F., Hartman W.D. Recent brachiopod—coralline sponge communites and their paleoecological significance // Science. 1971. V. 173. P. 623–625.
Logan A. Ecological observations on the recent articulate brachiopod Argyrotheca bermudana Dall, from the Bermuda platform // Bull. Biol. Sci. 1975. V. 25. P. 186–204.
Logan A., Zibrowius H. A new genus and species of rhynchonellids (Brachiopoda, recent) from submarine caves in the Mediterranean Sea // Publ. Staz. Zool. Napoli: Mar. Ecol. 1994. V. 15. № 1. P. 77–88.
Pakhnevich A.V. Reasons of micromorphism in modern and fossil brachiopods // Paleontol. J. 2009. V. 43. № 11. P. 1458–1468.
Pakhnevich A.V. Order Rhynchonellida Kuhn, 1949 // Paleontol. J. 2018. V. 52. № 8. P. 892–956.
Ruggiero E.T. Brachiopods of the Isca submarine cave: observations during ten years // 4th Intern. Brachiopod Congr., London, Great Britain, Vol.: The Millennium Brachiopod Congress. Abstracts. L.: Natur. Hist. Mus., 2000. P. 74.
Rzhonsnitskaya M.A., Mamedov A.B. Devonian stage boundaries in the southern Transcaucasus // Cour. Forsch.-Inst. Senckenb. 2000. V. 225. P. 329–333.
Saito M., Motchorova-Dekova N., Endo K. Recent brachiopod fauna from the submarine caves of Okinawa, Japan // 4th Intern. Brachiopod Congr., London, Great Britain, Vol.: The Millennium Brachiopod Congress. Abstracts. L.: Natur. Hist. Mus., 2000. P. 77.
Simon E., Willems G. Gwynia capsula (Jeffreys, 1859) and other recent brachiopods from submarine caves in Croatia // Bull. Inst. Roy. Sci. Natur. Belg. Biol. 1999. V. 69. P. 15–21.
Swedmark B. The interstitial fauna of marine sand // Biol. Rev. 1964. V. 39. P. 1–42.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал