

Палеонтологический журнал, 2020, № 6, стр. 112-120
Новые данные о строении стерильных и фертильных структур, спор in situ Svalbardia osmanica Petrosjan et Radczenko (археоптерисовые) из живета Кузбасса, Россия
А. Л. Юрина a, *, О. А. Орлова a, b, **, М. Г. Раскатова c, ***
a Московский государственный университет имени М.В. Ломоносова
Москва, Россия
b Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
c Воронежский государственный университет
Воронеж, Россия
* E-mail: flora.paleo@mail.ru
** E-mail: oowood@mail.ru
*** E-mail: mgraskatova@yandex.ru
Поступила в редакцию 08.10.2019
После доработки 13.12.2019
Принята к публикации 30.01.2020
Аннотация
Проведено дополнительное изучение Svalbardia osmanica Petrosjan et Radczenko из отложений изылинской свиты среднего девона Кузбасса на основе коллекции типового материала (ЦНИГР музей им. Ф.Н. Чернышева). Дан расширенный диагноз S. osmanica, включающий новые данные по морфологии и анатомии стерильных частей растения, а также его фертильных структур и спор in situ из микроспорангиев. Изученные в работе дисперсные споры из материнских пород, вмещающих растение, указывают на живетский возраст изылинской свиты.
ВВЕДЕНИЕ
Род Svalbardia, характерный компонент среднедевонских флор, был установлен О. Хёгом (Høeg, 1942) из верхней части среднедевонских или самой нижней части верхнедевонских? отложений архипелага Шпицберген. В качестве типового вида он указал S. polymorpha. Долгое время этот геологический интервал возраста типового вида отмечался во всех публикациях, связанных с родом Svalbardia (Петросян, Радченко, 1960; Лепехина и др., 1962; Carluccio et al., 1966; Stockmans, 1968; Chaloner, 1972; Matten, 1981; Meyen, 1987; Jurina, Raskatova, 2012 и др.). Х.-И. Швайтцер (Schweitzer, 1999) изучил стратиграфическое положение S. polymorpha в формации Плантерюгген (Planteryggen) Шпицбергена из местонахождений, откуда Хёг описал этот вид, и на основании данных по споровым комплексам откорректировал возраст типового вида как средне-позднеживетский.
Род Svalbardia в настоящее время отмечается, в основном, в живетских отложениях среднего девона (местонахождения на Шпицбергене, в Шотландии, Германии, Бельгии, Латвии и России), реже во франских отложениях верхнего девона (местонахождения в США и в России), а также в нерасчлененном верхнем девоне (местонахождения в Канаде и на Украине). В род включают более 10 видов: S. polymorpha Høeg, 1942; S. osmanica Petrosjan et Radczenko, 1960; S. boyi Kräusel et Weyland, 1960; S. avelinesiana Stockmans, 1968; S. scotica Chaloner, 1972; S. banksii Matten, 1981; S. fissilis (Schmalhausen) Matten, 1981; S. furcihasta (Krassilov, Raskatova et Istchenko) Jurina, 2014; S. acuta (Tschirkova-Zalesskaya) O. Orlova, Jurina et Snigirevsky, 2016; S. kuzbassica (S. Stepanov) Jurina, O. Orlova et Raskatova, 2016 (Юрина и др., 2016а); S. sphenophyllifolia (Lesquereux) Jurina et O. Orlova, comb. nov. и др. Основные признаки рода Svalbardia и его видов, дискуссия относительно его статуса, анализ публикаций по всем ранее установленным видам рода и его находкам были освещены ранее (Jurina, Raskatova, 2012, 2014).
Сравнительно хорошо изучено строение листовой пластинки представителей этого рода. Анатомическая структура осей и других частей растения ранее была не установлена. Высказывалось предположение (Carluccio et al., 1966), что петрификации Actinopodium nathorstii Høeg, 1942, найденные в одном местонахождении со Svalbardia polymorpha на Шпицбергене, являются анатомической структурой осей конечных ветвей S. polymorpha. Однако органическая связь петрификаций Actinopodium с вегетативными или фертильными осями Svalbardia до сих пор не доказана. За более чем 70-летнюю историю изучения рода Svalbardia осталось много невыясненных вопросов. Это связано, прежде всего, с не совсем удовлетворительной сохранностью ископаемого материала Svalbardia и довольно редкими его находками.
Целью настоящей работы является повторное изучение морфологического строения стерильных и фертильных структур S. osmanica, описание первых для этого вида находок инситных микроспор из микроспорангиев, установление соответствия последних таксону дисперсных миоспор, а также исследование анатомических особенностей осей, прежде не описанных. Поскольку для большинства видов рода Svalbardia споры из спорангиев неизвестны, обнаружение спор in situ еще у одного вида этого рода позволит подойти к решению проблемы родственных связей, как в пределах самого рода, так и с другими родами семейства Archaeopteridaceae.
Авторы выражают искреннюю признательность А.Р. Соколову и Н.М. Кадлец (ЦНИГР музей им. Ф.Н. Чернышева, С.-Петербург) за предоставленную возможность изучить коллекцию; Л.Г. Перегоедову (СНИИГГиМС, г. Новосибирск), Я.М. Гутаку (ИГДиГ, г. Новокузнецк), В.А. Антоновой (СибГИУ, г. Новокузнецк), С.А. Родыгину (ТГУ, г. Томск) за консультации по стратиграфии девона Кузбасса; Е.А. Жегалло и Р.А. Ракитову (ПИН РАН, г. Москва) за помощь в работе со сканирующим электронным микроскопом (СЭМ). Работа выполнена при финансовой поддержке РФФИ (проект № 19-04-00498).
МАТЕРИАЛ, МЕТОДИКА И ТЕРМИНОЛОГИЯ
Материалом для данной работы послужила типовая коллекция Svalbardia osmanica, хранящаяся в ЦНИГР музее им. Ф.Н. Чернышева (колл. № 9258), впервые описанная Н.М. Петросян и Г.П. Радченко (1960) из девонских отложений западных окраин Кузбасса. Благодаря любезности сотрудников музея, мы ознакомились с типовым материалом колл. № 9258 в самом музее, а также два экземпляра (№ 83/9258 и 97/9258), представленные отпечатками стерильных и фертильных частей растения, получили во временное пользование для детального исследования. Эти экземпляры были изучены с помощью СЭМ ТЕСКАН в режиме низкого вакуума в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН), затем некоторые фрагменты древесины фертильных и стерильных осей были смонтированы на столик для СЭМ, напылены золотом и изучены в СЭМ в режиме высокого вакуума.
Далее из спороносной ветви экз. № 97/9258 были извлечены инситные микроспоры. Споры извлекались в участках с разрушенной стенкой спорангиев по методике, описанной нами ранее (Jurina, Raskatova, 2014). В результате из четырех извлеченных спорангиев единичные тетрады и отдельные микроспоры были обнаружены только в двух. Кроме того, из окружающей ветвь породы методом современной мацерации карбонатно-терригенных пород (Раевская, Шурекова, 2011) был получен палинокомплекс, содержащий многочисленные миоспоры. Установленные инситные и дисперсные споры были изучены в СЭМ в ПИН РАН, а также в оптическом световом микроскопе LM POLAM-312 в Воронежском государственном ун-те (ВГУ).
В статье уточнена терминология для стерильных и фертильных структур некоторых археоптерисовых. До 60-х годов прошлого столетия при описании стерильных образований археоптерисовых использовались термины: вайя, перо, перышко. Исследования строения первичной сосудистой системы элементов “вайи” рода Archaeopteris (Dawson) Stur, 1875 (Ци, 2011) показали, что анатомия рахисов и перьев подобна анатомическому строению стебля (Carluccio et al., 1966). Было предложено вместо терминов “вайя, перо и перышко” употреблять термин “лист и его составляющие”. Поскольку Archaeopteris и Svalbardia представляют одну эволюционную группу, в данной работе мы следуем при описании рекомендованной Л. Карлуччо и соавторами терминологии.
Единой терминологии в описании элементов фертильной системы у исследователей не наблюдается. По-разному называются форма системы и составляющие ее элементы. Чаще всего употребляются термины: фертильные перья, части, ветви. Для общей характеристики фертильной системы археоптерисовых мы предлагаем использовать термин “стробилоподобная структура” (Юрина и др., 2016б), предложенный С.В. Мейеном (1987; Meyen, 1987). В содержание этого термина Мейен вкладывал понятие как о спороносной структуре, собранной в рыхлую группу, являющуюся переходной к плотным, хорошо оформленным стробилам. В стробилоподобной структуре нами различаются элементы: ось структуры; спорофилл и характер окончания спорофилла (простое или разветвленное); расположение спорангиев на определенной стороне спорофилла и тип расположения (одиночный или групповой). Не у всех археоптеридных спороносная структура выделяется как единая цельная. Иногда спорофиллы со спорангиями располагаются между стерильными образованиями (листьями на оси), т.е. фертильными зонами на стерильной структуре.
ВОЗРАСТ ВМЕЩАЮЩИХ ОТЛОЖЕНИЙ
Исследованное нами растение Svalbardia osmanica происходит из отложений изылинской свиты девона, широко развитой на западных окраинах Кузнецкого бассейна. В действующих по настоящее время “Постановлениях …” (1994) зафиксирован живетский возраст свиты. Остановимся на биостратиграфическом анализе макрофлоры и комплекса миоспор, позволяющем интерпретировать возраст вмещающих отложений изылинской свиты.
Петросян (Типовые разрезы…, 1992) привела комплекс флоры изылинской свиты Кузбасса в составе девяти видов: Svalbardia polymorpha Høeg, S. osmanica Radcz. et Petros., Archaeopteris sibirica Zal., A. fissilis Schmalh., A. kuzbassica S. Step., Dimeripteris gracilis Schmalh., Moresnetia zalesskyi Stockm., Condomophyton gracile Radcz. et Petros. и Platyphyllum sp. (принадлежность к родам дана по Петросян). Она сопоставляла этот комплекс флоры с гауйским комплексом растений Прибалтики, пашийским комплексом Русской платформы и яранским комплексом Тимано-Печорской области и сделала вывод о раннефранском возрасте флоры изылинской свиты. Отметим, что в настоящее время гауйские, пашийские и яранские отложения относятся к живетскому ярусу.
По нашему мнению, общий комплекс флоры изылинской свиты характеризуется большим разнообразием видов рода Svalbardia (S. polymorpha, S. osmanica, S. sphenophyllifolia, S. fissilis, S. kuzbassica), а представители рода Archaeopteris отсутствуют (Юрина и др., 2016а). Род Svalbardia является важным компонентом V комплексной родовой зоны Svalbardia, охватывающей большую часть живета (Banks, 1980). Эта зона часто используется для расчленения отложений по макрофлоре (Fairon-Demaret, 1986; Юрина, 1988; Снигиревский, 1997; Антонова, 2007; Gradstein et al., 2012; Гутак, Антонова, 2016). Флора изылинской свиты хорошо сопоставляется с комплексом флоры зоны Svalbardia, отвечающим второй половине живета.
Большое значение для определения возраста изылинской свиты имеют миоспоровые комплексы, установленные как прежними исследованиями Е.В. Чибриковой (Типовые разрезы…, 1992), так и нами. Чибрикова привела комплекс миоспор из нескольких местонахождений изылинской свиты, включающий в себя следующие 17 видов: Geminospora rugosa (Naum.) Obukh., G. micromanifesta (Naum.) Owens, G. micromanifesta (Naum.) Owens var. microspinosa Tschibr., G. notata (Naum.) Obukh., G. notata (Naum.) Obukh. var. asper Tschibr., G. nalivkinii (Naum.) Obukh., Aneurospora greggsii (McGregor) Streel, Ancyrospora incisa (Naum.) M. Rask. et Obukh., A. simplex Guennel, A. cf. fidus (Naum.) Obukh., Archaeotriletes heteromerus Nadl., Emphanizonotriletes naumovae Oshurk., Membrabaculisporis radiatus (Naum.) Arch., Stenozonotriletes conformis Naum., Ambitisporites definitus (Naum.) Oshurk., Verrucosisporites evlanensis (Naum.) Obukh., Retusotriletes communis Naum., R. raisae Tschibr. Cписок таксонов приведен по морфологической классификации М.В. Ошурковой (2003). Чибрикова не рассматривала стратиграфическую приуроченность и процентный состав видов, монографически их не описывала и не изображала, а только кратко отметила, что миоспоровый комплекс изылинской свиты по составу сходен с пашийско-тиманским комплексом Европейской части России. Важное ее замечание касалось присутствия в изылинском комплексе зонального вида Ancyrospora incisa, отмечающегося в основании этой свиты.
Нами выделен миоспоровый комплекс из отложений изылинской свиты местонахождения Осман, содержащих макрофлору. Ранее миоспоры из этого местонахождения не указывались. Выявленный комплекс дисперсных миоспор имеет много общих форм с комплексом, приводимым Чибриковой (Типовые разрезы…, 1992), но отличается бóльшим видовым разнообразием и присутствием стратиграфически важных видов, не отмеченных ранее, а именно: Geminospora micromanifesta (Naum.) Owens var. limbatus Tschibr., Reticulatisporites retiformis (Naum.) Obukh., Archaeozonotriletes variabilis Naum., Ancyrospora cf. melvillensis Owens. Отметим также встреченные нами, но не упоминаемые Чибриковой, виды: Punctatisporites pullatus (Naum.) Pashk., Calamospora simplicissima (Naum.) Oshurk., Cyclogranisporites rotundus (Naum.) Oshurk., Apiculiretusispora? hastata (Nadl.) Oshurk., Hymenozonotriletes connatus (Nadl.) Oshurk., Hystricosporites elongatus (Nadl.) Oshurk. и Contagisporites optivus (Tschibr.) Owens (табл. XIII , фиг. 3 ; см. вклейку). Таким образом, общее число видов миоспор изылинской свиты возросло до 31. В выделенном нами комплексе миоспор преобладают виды рода Geminospora, такие как G. micromanifesta (18%) (табл. XIII , фиг. 5 ), G. micromanifesta var. limbatus (6%) (табл. XIII , фиг. 7 ), G. rugosa (8%), G. notata (4%) и G. sp. (4%) (рис. 1, е, ж). Кроме того, представители этого рода часто встречаются в тетрадах (табл. XIII , фиг. 4 ). Также доминантным в комплексе отмечен вид Ancyrospora incisa (15%) (табл. XIII , фиг. 8 ), который является видом–индексом подзоны Ancyrospora incisа–Geminospora micromanifesta (IM), установленной для пашийского горизонта Восточно-Европейской платформы (Avkhimovich et al., 1993), т.е., биомаркером верхней части живетского яруса. По современным представлениям (Соболев, Евдокимова, 2008), пашийский горизонт коррелируют с верхнеживетским подъярусом. Типичными для уровня подзоны IM также являются миоспоры: Aneurospora greggsii (табл. XIII , фиг. 6 ), Ancyrospora simplex (рис. 1, з) и A. cf. fidus, присутствующие в комплексе спор изылинской свиты. Все вышеперечисленные особенности позволяют сопоставить комплекс миоспор изылинской свиты с комплексами миоспор живетского пашийского горизонта центральных районов Восточно-Европейской платформы (Avkhimovich et al., 1993), а также с живетским гауйским комплексом формации Лоде Латвии (Jurina, Raskatova, 2012). Среди отличий можно отметить присутствие в изылинской свите видов, не отмечаемых в комплексах подзоны IM, характеризующихся значительными размерами и крупными выростами: Hymenozonotriletes connatus (Nadl.) Oshurk.и Hystricosporites elongatus (Nadl.) Oshurk.
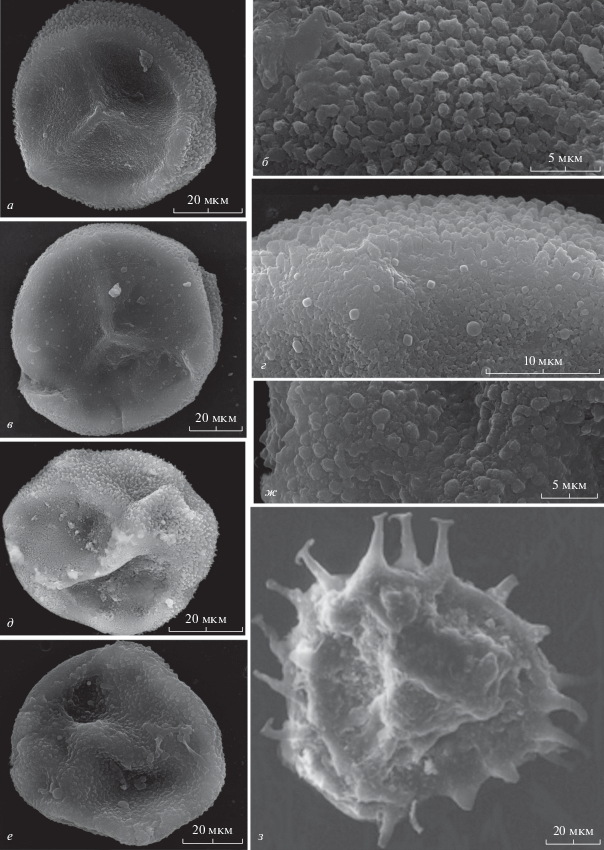
Рис. 1.
Микроспоры, извлеченные из микроспорангиев Svalbardia osmanica Petrosjan et Radczenko, emend. Jurina et al., и некоторые миоспоры из окружающей ветвь S. osmanica породы, СЭМ: а–д – Geminospora micromanifesta (Naum.) Owens, экз. № 97/9258: а, б, д – инситная микроспора, извлеченная из спорангия 1: а, д – проксимальная и экваториальная стороны; б – скульптура экваториальной поверхности; в, г – инситная микроспора, извлеченная из спорангия 2: в – проксимальная сторона; г – скульптура проксимальной и экваториальной поверхностей; е, ж – Geminospora sp., экз. № Kz-D-So-ins 2016: е – миоспора, дистальная сторона; ж – миоспора, скульптура дистальной поверхности; з – Ancyrospora simplex Guennel, экз. № Kz-D-So-dsp 2016, миоспора, проксимальная сторона; Кузнецкий бассейн, прав. берег р. Кондомы, в 0.5 км севернее ст. Осман; средний девон, живетский ярус, изылинская свита.
СПОРЫ in situ ПРЕДСТАВИТЕЛЕЙ РОДА SVALBARDIA
Изучение спор из репродуктивных структур растений является одним из важнейших направлений исследования девонских растений. Потребность в подобного рода работах связана со многими аспектами. Прежде всего, наиболее остро стоит вопрос о таксономической идентификации великого множества родов и видов дисперсных девонских спор со спорами, изученными из репродуктивных органов растений. Также исследование инситных спор является дополнительным звеном в установлении родственных связей растений. Настоящая работа представляет некоторый шаг к решению этих проблем. Среди известных видов рода Svalbardia фертильные структуры описаны у семи: S. polymorpha, S. osmanica, S. fissilis, S. furcihasta, S. scotica, S. sphenophyllifolia и S. kuzbassica (Høeg, 1942; Balme, 1995; Раскатова, 1995; Снигиревский, Снигиревская, 2008; Jurina, Raskatova, 2012, 2014; Orlova et al., 2016 и др.). У S. polymorpha, S. furcihasta, S. fissilis и S. osmanica споры извлечены из спорангиев растения, характеризующегося органической связью репродуктивной и вегетативной систем. Для этих видов установлены микроспоры, соответствующие роду Geminospora дисперсных спор, но разным его видам: G. micromanifesta (Naum.) Owens (у Svalbardia polymorpha и S. osmanica – описаны впервые в данной статье), Geminospora lemurata Balme emend. Playford (у Svalbardia furcihasta и S. fissilis) и Geminospora rugosa (Naum.) Obukh. (у Svalbardia fissilis). Мегаспоры Contagisporites optivus (Tschibr.) Owens и Biharisporites macromanifestus (Naum.) Owens выявлены в мегаспорангиях только у Svalbardia furcihasta и у S. fissilis.
Для свальбардий, у которых связь генеративных и вегетативных органов отсутствует (Медяник, 1982; Jurina, Raskatova, 2012), установлено, что микроспоры близки роду дисперсных миоспор Geminospora, мегаспоры – двум родам, Biharisporites и Contagisporites.
СИСТЕМАТИКА
КЛАСС PROGYMNOSPERMOPSIDA
ПОРЯДОК ARCHAEOPTERIDALES
Род Svalbardia Høeg, 1942
Svalbardia osmanica Petrosjan et Radczenko, emend. Jurina, O. Orlova et M. Raskatova
Табл. XIII , фиг. 1, 2 ; табл. XIV , фиг. 1–10 (см. вклейку)
Svalbardia osmanica: Петросян, Радченко, 1960, с. 43, табл. 9 , фиг. 4–7 ; Лепехина и др., 1962, с. 135, табл. XXIII , фиг. 2–8 .
Archaeopteris osmanica: Степанов, 1966, с. 111, табл. I , фиг. 1, 2 , табл. II , фиг. 1–4 , табл. III , фиг. 1, 3 , табл. IV , фиг. 1, 2, 4, 6 , табл. VI , фиг. 1, 2 ; 1975, с. 72, табл. XLV , фиг. 2–4 , табл. XLVI , фиг. 1–3 , табл. XLVII , фиг. 1–7 .
Голотип – ЦНИГР музей им. Ф. Н. Чернышева № 14/9258 (150/9258; Каталог…, 1996), отпечаток участка побега; Кузнецкий бассейн, прав. берег р. Кондомы, в 0.5 км севернее ст. Осман; средний девон, живетский ярус, изылинская свита; Петросян, Радченко, 1960, табл. 9 , фиг. 4 .
Diagnosis emended. Sterile structure consists of the longitudinally ribbed axes with the leaves helically attached to the axes. Leaves obliquus wedge-shaped, deeply dissected to eight linear, slightly curved segments, unequal in width and placed on different planes. Fertile structure is of cone-like type, longitudinally rectangular, with truncate apex. Loosely arranged sporophylls are decussate; sickle-shaped upward; terminated by simple or dichotomously divided sterile ends. Fusiform sporangia placed in clusters or singlely on the upper side of the sporophylls. Sporangia are sessile or attached by thin pedicel. Terminate axes of fertile and sterile structures are with scalariform tracheids of primary xylem. Secondary xylem is with uni-multiseriate porosity on radial walls of tracheids.
Описание (рис. 1, а−д). Сохранившиеся отпечатки представлены фрагментами стерильных и фертильных структур (табл. XIII , фиг. 1, 2 ). У стерильных структур (табл. XIII , фиг. 2 ) наблюдаются стержни как с прикрепленными к ним листовыми пластинками (табл. XIV , фиг. 2 ), так и только с их основаниями (табл. XIV , фиг. 3 ). Стержни прямые, толщиной 1−3 мм, с отчетливыми продольными ребрами до 0.5 мм ширины. Листья отходят от стержня под острым углом 10○−30○ и располагаются спирально через неравные промежутки длиной 3−12 мм. Листовая пластинка (табл. XIV , фиг. 2 ) клиновидной формы длиной 10−15 мм, состоит из дихотомирующих два, чаще четыре раза глубоко рассеченных неравных по длине восьми сегментов. Преобладают сегменты шириной 2−3 мм. Сегменты линейные, слегка изогнутые, располагающиеся в разных плоскостях, так как на отпечатках наблюдается то полная ширина сегмента, то только почти нитевидная. Основание листовой пластинки представлено узким низбегающим черешком длиной до 4−7 мм.
На осях стерильной и фертильной структуры (табл. XIV , фиг. 4–10 ) фрагментарно сохранились некоторые элементы первичной и вторичной ксилем. Первичная ксилема сложена очень узкими длинными трахеидами с лестничными окаймленными порами (табл. XIV , фиг. 4, 6, 8, 10 ). Трахеиды в сечении полигональные, диаметром 10–16 мкм, в среднем 12 мкм. Окаймления пор радиальных стенок трахеид шириной 6–8 мкм, высотой до 4 мкм; апертуры пор радиальных стенок трахеид шириной 4–5 мкм, высотой 1 мкм. Трахеиды различной, в основном, фрагментарной сохранности, представлены как внутренними слепками трахеид и камер пор, так и в виде их отпечатков наружной поверхности с очертаниями апертур и окаймлений. Клетки сердцевины не сохранились.
Вторичная ксилема (табл. XIV , фиг. 7 ) сохранилась крайне фрагментарно и представлена частично разрушенными, относительно узкими и длинными трахеидами со скошенными концами. Диаметр трахеид вторичной ксилемы варьирует в радиальном измерении от 15 до 30 мкм. Трахеиды вторичной ксилемы по форме полигональные, с одно-многорядной поровостью на радиальных стенках. Поры округлые, иногда округло-шестиугольные, диаметром 8–12 мкм. Когортоидная поровость на радиальных стенках трахеид не выявлена. Сердцевинные лучи и характер поровости полей перекреста не сохранились.
Стробилоподобная структура (табл. XIII , фиг. 1) продольно прямоугольного очертания длиной 30−45 мм и шириной 10−13 мм. Она состоит из супротивно расположенных спорофиллов. Стробилоподобная структура рыхлая. Ее ось относительно широкая, 1.5–2.0 мм шириной, продольно ребристая, подобно оси стерильной структуры. Спорофиллы отходят от оси под почти прямым углом и затем резко изгибаются вверх, приобретая почти параллельное оси стробила положение. Верхушка спорофилла дихотомически ветвится на тонкие короткие окончания, редко верхушка неразветвленная. На горизонтальной верхней части спорофилла наблюдаются веретеновидные прямостоящие с заостренной верхушкой спорангии 1.5−2.0 мм длиной, 0.4−0.5 мм шириной, расположенные группами от двух до шести спорангиев на одном спорофилле (табл. XIV , фиг. 1). Размеры спорангиев на разных спорофиллах приблизительно равны, одни спорангии сидячие, другие прикрепляются к спорофиллу тонкой ножкой.
Инситные микроспоры (рис. 1, а) радиальные, трехлучевые, каватные, округло-треугольные, 60–70 мкм в экваториальном диаметре. Щель разверзания простая с возвышающимся апексом (до 3 мкм), лучи щели прямые, длина лучей составляет полную длину радиуса центрального тела. На концах лучей наблюдается треугольное утолщение экзины (от 2 до 4 мкм). Слои экзины расслаиваются в районе экватора и дистальной полусферы. Интэкзина тонкая, образует отчетливое центральное тело округлого очертания. Экзоэкзина толстая (до 7−8 мкм) особенно на дистальной стороне, окружает центральное тело и в районе экватора формирует узкую плотную зону. На проксимальной поверхности происходит утонение экзины (до 2−4 мкм). Скульптура экзины дистальной и экваториальной частей споры представлена удлиненными бугорками, диаметр основания которых 1.0–1.2 мкм, высота – 1.0−1.8–2.5 мкм (рис. 1, б). Вершина бугорков плоская или округлая; иногда бугорки сменяются булавами с более или менее отчетливыми округлыми головками, расположенными в верхней части тонких стержней. Скульптурные элементы иногда соединены по два и образуют выросты с верхней суженной частью в виде мелких сосочков или округлых структур размером 0.3−0.5 мкм. Скульптура экзины проксимальной стороны мелкозернистая. В зоне перехода к экваториальной части наблюдается изменение характера скульптуры экзоэкзины, выражающееся в разрастании тонкого зернистого слоя в бугорки (рис. 1, д). Микроспора, извлеченная из другого спорангия того же экземпляра, при общем морфологическом подобии с вышеописанными микроспорами, несет дополнительную скульптуру экзины проксимальной поверхности, которая представлена редко расположенными круглыми бугорками размером от 0.5 до 1.2 мкм (рис. 1, в, г). Инситные мегаспоры в спорангиях не обнаружены.
Сравнение. S. osmanica отличается от других видов этого рода строением листовой пластинки, которая рассечена на более широкие (до 5 мм) сегменты. Ширина сегментов у S. banksii и S. furcihasta не более 1 мм, у S. polymorpha и S. fissilis сегменты нитевидные. Кроме того, для S. osmanica характерен длинный низбегающий черешок у листовой пластинки, а для S. polymorpha и S. banksii – короткий прямостоячий. По размерам листовой пластинки, числу сегментов в ней и неравной их длине S. osmanica имеет наибольшее сходство с S. fissilis, от которого отличается, прежде всего, формой сегментов листа: сегменты S. osmanica линейные, а у S. fissilis щетинковидные.
Инситные микроспоры S. osmanica имеют сходство с микроспорами из спорангиев S. polymorpha, но отличаются более отчетливым развитием кавы и присутствием более выраженной скульптуры экзоэкзины проксимальной стороны. Также описанные нами микроспоры имеют большое сходство с микроспорами из спорангиев S. furcihasta по типу строения спор (трилетные, каватные, зонатные). Отличие состоит в больших размерах микроспор из спорангиев S. osmanica, в больших размерах их кавы, в разной длине лучей щели и различных размерах скульптурных элементов экзоэкзины дистальной поверхности.
Замечания. Ряд морфологических признаков изученных микроспор, таких как характер утолщения экзины, ее расслоение на интэкзину и экзоэкзину и тип скульптуры экваториальной стороны и дистальной полусферы позволяют сопоставить их с дисперсными миоспорами Geminospora micromanifesta (Naum.) Owens. Подобный тип микроспор был ранее установлен у Svalbardia polymorpha.
Распространение. Средний девон (живет) Алтае-Саянской горной области: местонахождения на прав. берегу р. Кондомы и р. Курундус; верхний девон Северного Тимана: верховья р. Сула (по сборам Л.С. Коссового, 1955 из: Петросян, Радченко, 1960).
Материал. Более 20 экз. удовлетворительной сохранности, представленных отпечатками вегетативной и фертильной системы (колл. № 9258), из которых два экз. хорошей и удовлетворительной сохранности изучались детально с помощью СМ, СЭМ и с выделением инситных спор.
ЗАКЛЮЧЕНИЕ
Дополнительное комплексное изучение типового материала S. osmanica дало возможность подойти к решению вопросов, не обсуждаемых ранее, и сделать следующие выводы.
1. Терминология морфологического описания стерильных и фертильных структур растения уточнена. Для описания стерильной системы предложен термин лист и его составляющие, вместо ранее широко употребляемых терминов вайя, перо и перышко; в описании фертильной системы используется понятие стробилоподобная структура. Эти термины впервые включены в расширенный диагноз S. osmanica.
2. Впервые изучено анатомическое строение фертильных и стерильных осей S. osmanica, в результате чего показано, что трахеиды первичной ксилемы очень узкие, с лестничными утолщениями стенок; вторичная ксилема представлена узкими трахеидами с одно-многорядной поровостью радиальных стенок. К сожалению, нами не установлена когортоидная поровость, характерная для представителей археоптерисовых рода Callixylon (Archaeopteris). Скорее всего, это связано с сохранностью и крайней фрагментарностью изученного материала.
3. В спорангиях стробилоподобной структуры S. osmanica установлены микроспорангии, содержащие микроспоры, идентичные дисперсным миоспорам Geminospora micromanifesta. Взаимное расположение мега- и микроспорангиев на растении и их отличительные признаки (размеры, форма, количество спор в спорангиях) не выявлены. Эта проблема касается не только S. osmanica, но также других видов этого рода.
4. На сравнительно ограниченном материале показано, что микроспоры видов рода Svalbardia (S. polymorpha, S. furcihasta. S. osmanica и, возможно, S. fissilis) идентичны разным видам рода дисперсных спор Geminospora: G. micromanifesta, G. lemurata и G. rugosa; мегаспоры схожи с дисперсными мегаспорами Contagisporites optivus (Svalbardia furcihasta) и, возможно, с дисперсными мегаспорами Biharisporites macromanifestus (Svalbardia fissilis).
5. Сделан вывод о живетском возрасте изылинской свиты Кузбасса: данные по макрофлоре свидетельствуют о второй половине живетского века, так как комплекс дисперсных миоспор сходен с комплексом миоспор верхнего живета. Пока не найдено решение вопроса палеогеографического значения находок свальбардий.
Список литературы
Антонова В.А. Флора сергиевской свиты (поздний девон) Кузбасса // Вестн. Томск. гос. ун-та. Науки о Земле. 2007. № 304. С. 195−198.
Гутак Я.М., Антонова В.А. Корреляция средне-верхнедевонских отложений Кузнецкого прогиба (западная часть Алтае-Саянской складчатой области) // Матер. Межведомств. совещ. “Общая стратиграфическая шкала и методические проблемы разработки региональных стратиграфических шкал России”. СПб.: ВСЕГЕИ, 2016. С. 58−60.
Каталог голотипов видов фауны и флоры, хранящихся в ЦНИГР музее. Вып. 3. Палеозой, мезозой, кайнозой. СПб.: Изд-во ВСЕГЕИ, 1996. 354 с.
Лепехина В.Г., Петросян Н.М., Радченко Г.П. Важнейшие девонские растения Алтае-Саянской горной области // Матер. к фитостратиграфии девонских отложений Алтае-Саянской горной обл. Л., 1962. С. 61−188 (Тр. ВСЕГЕИ. Нов. сер. Т. 70).
Медяник С.И. Спороношение нижнефранского Archaeopteris из Южного Тимана // Палеонтол. журн. 1982. № 2. С. 121−127.
Мейен С.В. Основы палеоботаники. М.: Недра, 1987. 403 с.
Ошуркова М.В. Морфология, классификация и описания форма-родов миоспор позднего палеозоя. СПб.: Изд-во ВСЕГЕИ, 2003. 377 с.
Петросян Н.М., Радченко Г.П. Новый вид свальбардии // Hовые виды древних растений и беспозвоночных СССР. Ч. I. М.: Наука, 1960. С. 43−45.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 27 [Текст]. СПб.: ВСЕГЕИ, 1994. 67 с.
Раевская Е.Г., Шурекова О.В. Современные технологии и оборудование в обработке карбонатно-терригенных пород для палинологического анализа // Проблемы современной палинологии. Матер. XIII Росс. палинол. конф. Т. 1. Сыктывкар: ИГ Коми НЦ УрО РАН, 2011. С. 103−107.
Раскатова М.Г. Споры Tanaitis (археоптеридные) и дисперсные миоспоры зоны Contagisporites optivus – Spelaeotriletes krestovnikovii // Палинология в России. Статьи росс. палинологов к IX Междунар. палинологическому конгрессу в Хьюстоне, Техас. Ч. 1. М., 1995. С. 60−69.
Снигиревский С.М. Позднедевонская флора Северного Тимана. Автореф. дисс. … канд. геол.-мин. наук. СПб., 1997. 16 с.
Снигиревский С.М., Снигиревская Н.С. Некоторые новые данные о девонских спорах in situ и комментарии к номенклатуре дисперсных спор // Палинология: стратиграфия и геоэкология. Сб. научн. тр. XII Всеросс. палинол. конф. (29.09–4.10.2008 г., СПб.). Т. 1. СПб.: ВНИГРИ, 2008. С. 108–115.
Соболев Н.Н., Евдокимова И.О. Девонская система // Постановления Межведомств. стратигр. ком-та России. Вып. 38 / Ред. Жамойда А.И., Петров О.В. СПб.: ВСЕГЕИ, 2008. С. 52–60.
Степанов С.А. О флоре Archaeopteris с окраин Кузбасса // Тр. ТГУ. Сер. геол. 1966. Т. 184. С. 106–116.
Степанов С.А. Фитостратиграфия опорных разрезов девона окраин Кузбасса. Новосибирск: Западно-Сибирское книжн. изд-во, 1975. 152 с. (Тр. СНИИГГиМС. Вып. 211).
Типовые разрезы пограничных отложений среднего и верхнего девона, франского и фаменских ярусов окраин Кузнецкого бассейна // Матер. V выездной сессии комиссии МСК по девонской системе, Кузбасс, 16–29 июля 1991 г. / Ред. Краснов В.И., Ржонсницкая М.А., Гутак Я.М. Новосибирск: СНИИГГиМС, 1992. 136 с.
Ци В. Правильное указание автора для позднедевонского рода Archaeopteris (Progymnospermopsida) // Палеонтол. журн. 2011. № 3. С. 106–108.
Юрина А.Л. Флора среднего и позднего девона Северной Евразии. М.: Наука, 1988. 176 с. (Тр. ПИН АН СССР. Т. 227).
Юрина А.Л., Орлова О.А., Раскатова М.Г. О возрасте изылинской свиты (западная окраина Кузнецкого бассейна) по палеоботаническим данным (макрофлоре и миоспорам) // Матер. межведомственного совещ. “Общая стратиграфическая шкала и методические проблемы разработки региональных стратиграфических шкал России”. СПб.: ВСЕГЕИ, 2016а. С. 190–192.
Юрина А.Л., Орлова О.А., Снигиревский С.М. Об особенностях строения фертильных структур некоторых средне–позднедевонских археоптерисовых // Палеострат–2016. Годичн. собр. секции палеонтол. МОИП и Московского отд. Палеонтол. об-ва. М.: ПИН РАН, 2016б. С. 88.
Avkhimovitch V.I., Tchibrikova E.V., Obukhovskaya T.G. et al. Middle and Upper Devonian miospore zonation of Eastern Europe // Bull. Centre Rech. Explor.-Production Elf Aquitaine. 1993. V. 17. P. 79–147.
Balme B.E. Fossil in situ spores and pollen grains: an annotated catalogue // Rev. Palaeobot. Palynol. 1995. V. 87. P. 81–323.
Banks H.P. Floral assemblages in the Silurian-Devonian // Biostratigraphy of fossil plants. Stroudsburg: Dowden, 1980. P. 1–24.
Carluccio L.M., Hueber F.M., Banks H.P. Archaeopteris macilenta, anatomy and morphology of its frond // Amer. J. Bot. 1966. V. 53. № 7. P. 719–730.
Chaloner W.G. Devonian plants from Fair Isle, Scotland // Rev. Palaeobot. Palynol. 1972. V. 14. P. 49–61.
Fairon-Demaret M. Some uppermost Devonian megafloras: a stratigraphical review // Ann. Soc. Géol. Belg. 1986. V. 109. P. 43–48.
Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. The Geologic Time Scale. Amsterdam: Elsevier, 2012. 1176 p.
Høeg O.A. The Downtonian and Devonian flora of Spitzbergen // Norg. Svalb. Ishavs-Undersøk. 1942. V. 83. 228 p.
Jurina A., Raskatova M. New data on the Devonian plant and miospores from the Lode Formation, Latvia // Sci. Pap. Univ. Latvia. Earth and Environm. Sci. 2012. V. 783. P. 46–56.
Jurina A., Raskatova M. Svalbardia from Givetian of Central Russia (Voronezh Region): leaf morphology and spores from sporangium // Palaeobotanist. 2014. V. 63. P. 99–112.
Kräusel R., Weyland H. Die neue Pflanzen aus dem Devon // Palaeontogr. Abt. B. 1960. Bd 107. S. 65–82.
Matten L.C. Svalbardia banksii sp. nov. from the Upper Devonian (Frasnian) of New York State // Amer. J. Bot. 1981. V. 68. № 10. P. 1383–1391.
Meyen S.V. Fundamentals of Palaeobotany. L.–N. Y.: Chapman and Hall, 1987. 432 p.
Orlova O.A., Jurina A.L., Snigirevsky S.M. Late Devonian plant communities of North Russia // Rev. Palaeobot. Palynol. 2016. V. 224. P. 94–107.
Schweitzer H.-J. Die Devonfloren Spitzbergens // Palaeontogr. Abt. B. 1999. Bd 252. S. 1–122.
Stockmans F. Végétaux Mésodévoniens récoltés aux confins du Massif du Brabant (Belgique) // Mém. Inst. Roy. Sci. Natur. Belg. 1968. V. 159. P. 1–49.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал