

Палеонтологический журнал, 2021, № 2, стр. 107-118
Pseudoschizaea sp. из ранней юры Италии: строение и сравнение
М. В. Теклева a, *, С. В. Полевова b, О. А. Гаврилова c, Г. Роги d, М. Нери e
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Московский государственный университет имени М.В. Ломоносова
Москва, Россия
c Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
d Институт наук о Земле
Падуя, Италия
e Музей естественной истории Виньолы
Виньола, Италия
* E-mail: tekleva@mail.ru
Поступила в редакцию 27.12.2019
После доработки 31.03.2020
Принята к публикации 22.05.2020
Аннотация
Исследована дисперсная палиноморфа Pseudoschizaea sp. неизвестного родства из юрских отложений Италии с помощью светового, конфокального, флуоресцентного, сканирующего и трансмиссионного электронных микроскопов. Проведено сравнение со спорами мхов и грибов, цистами водорослей и яйцами червей. Сходный характер скульптуры поверхности оболочки спор и цист наблюдается у некоторых водорослей и грибов, сходный размерный ранг также обнаружен в этих группах. Поперечная исчерченность однослойной оболочки, наблюдаемая у Pseudoschizaea, пока не обнаружена у исследованных с помощью трансмиссионного электронного микроскопа спор грибов и цист водорослей, хотя в отдельных слоях оболочек некоторых представителей водорослей наблюдаются каналы. Обсуждается возможное родство и экология продуцентов данной палиноморфы.
ВВЕДЕНИЕ
Сфероидальные и эллипсоидальные уплощенные палиноморфы с характерным концентрическим ребристым рисунком поверхности и размером около 35–40 мкм представлены в отложениях разной природы, разных географических областей и разного возраста (начиная с палеозоя практически до настоящего времени) и объединяются под родовым названием Pseudoschizaea Thiergart et Frantz ex R. Potonie emend. или под группой родов Pseudoschizaea, Circulisporites de Jersey, Concentricystes Rossignol и Chomotriletes Naumova. Несмотря на это, данные палиноморфы до настоящего времени были в основном изучены только на световом уровне (СМ), реже с помощью сканирующего электронного микроскопа (СЭМ), и предположения об их родственных связях довольно умозрительны. Это значительно обедняет интерпретацию отложений, где встречаются эти остатки (учитывая их широкое распространение), и понимание экологии и эволюции организмов, которые могли продуцировать подобный тип палиноморф. Возможные интерпретации данной палиноморфы разными авторами включают споры высших растений, цисты водорослей, споры грибов, яйца ряда групп беспозвоночных (см., напр., обзор в: Scott, 1992). Для аккуратной интерпретации этих палиноморф необходимо их детальное изучение в световых и электронных микроскопах с получением максимального объема информации, что явилось целью настоящей работы.
Исследование поддержано грантом РФФИ № 17-04-01094 для М.Т. и С.П., исследования в Центре коллективного пользования научным оборудованием “Клеточные и молекулярные технологии изучения растений и грибов” БИН РАН проводились в рамках госзадания № AAA-A18-118031690084-9. Авторы благодарны В.Ф. Тарасевич (БИН РАН) и Н.Е. Завьяловой (ПИН РАН) за ценные замечания, М. Брейгиной и Н. Максимову (каф. физиологии растений МГУ) за помощь в работе на флуоресцентном микроскопе.
МАТЕРИАЛ И МЕТОДЫ
Изученный образец происходит из отложений свиты Ротцо (Rotzo Formation) недалеко от с. Беллори, пров. Верона, северо-восток Италии (рис. 1). Эта свита относится к группе Калкари Гриджи (Calcari Grigi), из нее описаны ископаемые остатки моллюсков, фораминифер, растений из янтаря и отпечатки следов динозавров (Wesley, 1956, 1958; Bosellini et al., 1971; Clari, 1975; Avanzini et al., 2006; Neri et al., 2017). Возраст двух слоев с углистой глиной, содержащих Pseudoschizaea, соответствует плинсбаху (см. подробнее в: Neri et al., 2017, 2018; рис. 2). Состав и процентное содержание палинологического комплекса приведены в работе М. Нери с соавт. (Neri et al., 2017). Для палиноморфологического анализа образцы были подготовлены по стандартной методике, отмацерированы и обработаны с помощью HCl, HF и HNO3. Полученный после промывки и просеивания осадок был заключен в глицерин для дальнейших исследований. Палинологический анализ данного образца выявил содержание гладких и орнаментированных спор, принадлежащих папоротникообразным, и пыльцевых зерен Circumpolles.
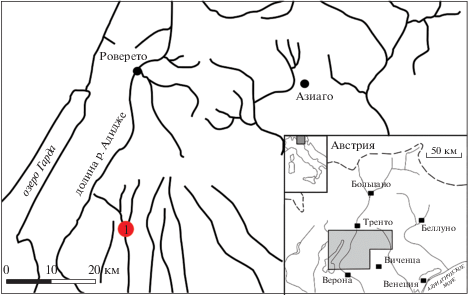
Рис. 2.
Схема изученной части разреза: a – схема разреза; б – часть разреза, содержащая слои с янтарем. Обозначения: 1) глина, богатая органическим веществом и углем, 2) известняк массивный, от серого до жeлтого цвета, 3) тонкозернистый известняк, 4) мергелевый известняк. C1–C4 – отобранные образцы, C1 – изученный образец.

Образец просматривался под световым микроскопом Olympus CX41, обнаруженные палиноморфы Pseudoschizaea переносились поодиночке на новое предметное стекло в каплю глицерина. Дальнейшая обработка для СМ, СЭМ и трансмиссионного электронного микроскопа (ТЭМ) проведена согласно методике, описанной в монографии Н.Е. Завьяловой с соавт. (Zavialova et al., 2018). Столик для СЭМ с перенесенными на него пыльцевыми зернами напылялся золотом и изучался с помощью СЭМ TESCAN VEGA-II XMU (ускоряющее напряжение 30 кВ). Ультратонкие срезы были сделаны с помощью ультратома Leica UC6. Часть срезов были дополнительно контрастированы цитратом свинца и уранил ацетатом и изучены с помощью ТЭМ Jeol 100 B и Jeol 1011 (ускоряющее напряжение 80 кВ). Другая часть срезов была изучена без дополнительного контрастирования на тех же микроскопах. Пыльцевые зерна были также изучены с помощью конфокального (КЛСМ) микроскопа LSM 780 [методика описана в работе О.А. Гавриловой с соавт. (Gavrilova et al., 2018)]. Всего было исследовано в СМ, СЭМ и ТЭМ 15 палиноморф.
Споры и цисты водорослей из юрских отложений России и девонских отложений Польши были использованы в сравнительных целях при изучении автофлуоресценции клеточных стенок с помощью флуоресцентного микроскопа (ФМ). Для изучения автофлуоресценции использовали исследовательский микроскоп Axioplan 2 imaging MOT, цифровую камеру AxioCam HRc и пакет программ AxioVision 4.7 (Zeiss, Германия). Флуоресценцию возбуждали ртутной лампой. Для анализа флуоресценции использовали разные блоки фильтров. Для автофлуоресценции возбуждение – в диапазоне 540–552 нм, регистрация – при длинах волн > 590 нм, применяемых при регистрации автофлуоресценции спорополлениновых оболочек современных и ископаемых пыльцы и спор (Matveyeva et al., 2012). Измерения исследованных палиноморф сделаны в СМ, СЭМ и ТЭМ. В статье использована терминология М. Хессе с соавт. (Hesse et al., 2009). В работе применялось оборудование Центра коллективного пользования ПИН РАН (СМ и СЭМ), биологического фак-та МГУ им. М.В. Ломоносова (ТЭМ, ФМ) и Ботанического ин-та им. В.Л. Комарова (КЛСМ) в Центре коллективного пользования научным оборудованием “Клеточные и молекулярные технологии изучения растений и грибов” БИН РАН.
Изученный образец №С1, хранится в Ин-те наук о Земле, Падуя, Италия.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
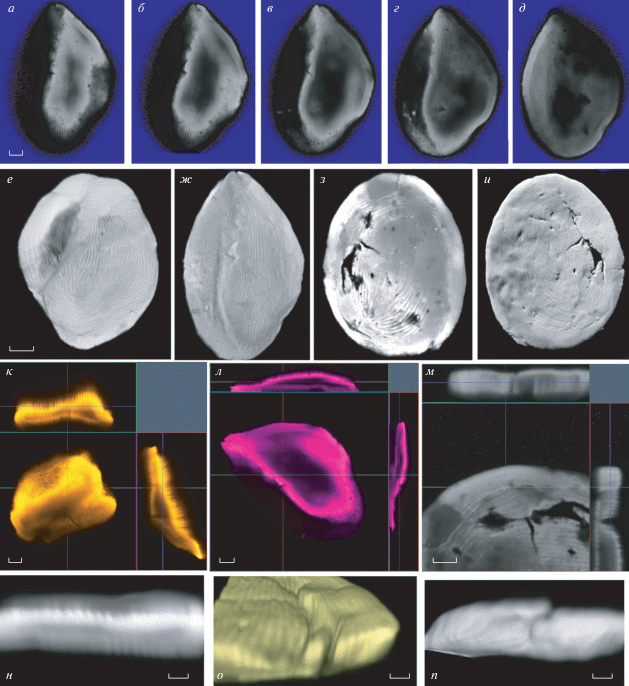
СМ. Палиноморфа эллипсоидальная (рис. 3, а, б, г) или сфероидальная (рис. 3, в, д, е), 37.3 (30.7–44.8) × 47.0 (42.3–50.8) мкм, уплощенная. Поверхность представлена характерным концентрическим ребристым рисунком, “завернутым” вокруг располагающегося в центральной области эллиптического (редко округлого) участка. На поверхности встречаются редкие, иногда довольно крупные, нерегулярно расположенные перфорации (рис. 4, а–е). Никаких следов апертуры или сходной структуры не выявлено. Оболочка относительно толстая, однослойная.
Рис. 3.
Pseudoschizaea sp., обр. № С1, СМ: a, б, г – эллипсоидальная форма; в, д, е – сфероидальная форма. Длина масштабной линейки 10 мкм.

Рис. 4.
Pseudoschizaea sp., обр. № С1, СЭМ: а–е – общий вид палиноморфы; ж–и – участки поверхности при большем увеличении, видны ребра и перфорации. Длина масштабной линейки: а–е – 10 мкм; ж–и – 5 мкм.

СЭМ. Размеры палиноморфы в СЭМ: 35.9 (30.4–41.4) × 47.2 (42.8–52.2) мкм. Скульптура поверхности ребристая, с редкими, иногда довольно крупными, нерегулярно расположенными перфорациями (рис. 4, а–е), ширина ребер варьирует от 0.14 до 0.21 мкм (рис. 4, ж–и).
ТЭМ. Оболочка неравномерной толщины по периметру (рис. 5, а–г), в среднем 1.35 (0.2–4.1) мкм. Ультраструктура оболочки состоит из одного гомогенного слоя неравномерной толщины с “зубцами” как с наружной, так и с внутренней сторон. Большая толщина обычно наблюдается в латеральных областях палиноморфы (рис. 5, д–ж). Наружные “зубцы” соответствуют ребристому рисунку поверхности, внутренние “зубцы” являются продолжением (противоположным концом) наружных “зубцов”. Местами также наблюдается исчерченность сквозь толщу оболочки (рис. 5, д–ж). Ширина “зубцов” 0.3 (0.2–0.4) мкм; в основном, они различимы в местах сгиба в латеральных областях, остальные участки оболочки выглядят гомогенными. Ковнутри от оболочки местами наблюдаются обрывки бесструктурного слоя – возможно, это несохранившийся внутренний слой оболочки, либо артефакт (рис. 5, д, ж).
Рис. 5.
Pseudoschizaea sp., обр. № С1, ТЭМ: а–г – срезы через палиноморфу, общий вид; д–ж – участки оболочки, видна поперечная исчерченность и плохо сохранившийся внутренний слой (стрелка). Длина масштабной линейки: а, д, е – 1.25 мкм; б, в – 5 мкм; г – 2.5 мкм; ж – 0.5 мкм.

КЛСМ. Палиноморфы эллипсоидальные или сфероидальные, уплощенные, поверхность ребристая, ребра тонкие. В СМ и СЭМ у некоторых палиноморф в оболочке просматривались округлые микроотверстия (в числе одного–двух на палиноморфу); возникло предположение, что эти отверстия являются своего рода апертурами. Однако при КЛСМ исследовании серийных оптических срезов (рис. 6, а–д) было выявлено, что это трещины, идущие в разных направлениях и видимые на сгибе фоссилизированной оболочки (рис. 6, о, п). Трещины часто обнаруживаются в оболочке палиноморф (рис. 6, з, и, м). Палиноморфы сплющены неравномерно, внутри часто образуется полость (рис. 6, к, л). Реконструированные из срезов изображения палиноморф демонстрируют ребристую поверхность; ребра тонкие, длинные, в основном однонаправленные, довольно плотно расположенные. КЛСМ выявляет все признаки поверхности (рис. 6, е, ж), доступные для исследования с помощью СЭМ. КЛСМ привлекали также для помощи в решении еще одного вопроса, а именно – реконструкции внутренней поверхности оболочки (рис. 6, з). На внутренней поверхности реконструированной палиноморфы иногда заметны “зубцы” (рис. 6, з), также иногда обнаруживается “поперечная полосатость” (рис. 6, м–о) оболочки, что может свидетельствовать о том, что оболочка сформирована из ряда узких, близко прилегающих друг к другу “полосок”. Однако, в других случаях оболочка гомогенная, вероятно, в тех случаях, когда срез проходит вдоль “полосок”.
Рис. 6.
Pseudoschizaea sp., обр. № С1, КЛСМ: а–д – серия оптических срезов через одну палиноморфу; е, ж, и – общий вид реконструированных палиноморф; з – внутренняя поверхность палиноморфы “и”; к–м – “ortho"-режим (проекция трехмерных объектов на двумерную поверхность): срезы через три палиноморфы е, ж, и соответственно, показывающие оптический срез и две его проекции; н – виртуальный срез через реконструированную палиноморфу перпендикулярно длинной оси морфы и ребрам на ее поверхности, о, п – реконструированные части палиноморф, показывающие трещины оболочки. Длина масштабной линейки: а–д, к–м – 5 мкм; е–и –10 мкм; н–п – 2 мкм.
ФМ. При исследовании автофлюоресценции Pseudoschizaea светилась слабо при всех светофильтрах (рис. 7, а, г). Сходную интенсивность ответа демонстрировали исследованные для сравнения ископаемые споры и пыльцевые зерна (рис. 7, б, д), а также водоросли [Endoscrinium (Klement) T.F. Vozzhennikova, Fromea Cookson et Eisenack, Pareodinia Deflandre, Tubotuberella Vozzhennikova] (рис. 7, в, е). Заметно отличались по интенсивности автофлуоресценции на разных светофильтрах палиноморфы и фрагменты кутикулы. Кутикулы ярко светились, особенно на длинах волн больше 590 нм в обоих исследованных образцах (рис. 7, д, е).
Рис. 7.
а–в – СМ, г–е – ФМ: а – Pseudoschizaea sp., обр. № С1; б – дисперсные споры и остатки кутикулы из девонских отложений Польши; в – остатки кутикулы и водоросли (Tubotuberella и Fromea) из юрских отложений России; г – Pseudoschizaea sp., обр. № С1; д – дисперсные споры и остатки кутикулы из девонских отложений Польши; е – остатки кутикулы и водоросли (Tubotuberella и Fromea) из юрских отложений России. Длина масштабной линейки: а, б, г, д – 30 мкм; в, е –100 мкм.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ ИССЛЕДОВАНИЯ
Отмеченное ранее и отраженное в названии Pseudoschizaea сходство со спорами схизейных папоротников не отражает их родственные отношения. Сходство это выражается в характере поверхности палиноморф, однако, у Pseudoschizaea отсутствует щель разверзания (и вообще какая-либо апертура) и значительно отличается ультраструктура оболочки. У схизейных никогда не наблюдается исчерченности и внутренних “зубцов”, характерных для Pseudoschizaea (Tryon, Lugardon, 1991; Skog, 1993; Parkinson, 1994, 1995; Ramos Giacosa et al., 2015; Ramos Giacosa, Barakat, 2018). То же можно сказать и про большинство остальных споровых и всех семенных растений: комплекс признаков строения спородермы (отсутствие апертуры, выраженной морфологически или ультраструктурно; характерная исчерченность оболочки; ребристость, выраженная как в наружной, так и во внутренней части оболочки) однозначно отвергает принадлежность данного материала к каким-либо остаткам споровых или семенных растений. Поверхность Pseudoschizaea мы, вслед за многими исследователями (напр., Christopher, 1976; Scott, 1992), описываем как ребристую; часть других авторов (напр., Milanesi et al., 2006) иногда называют ее струйчатой. Ребристая (plicate) поверхность часто захватывает несколько слоев эктэкзины или даже все слои эктэкзины, в то время как струйчатая (striate) захватывает лишь покров или его часть. В нашем случае из этих двух терминов “ребристая поверхность” больше подходит для описания Pseudoschizaea. Однако, учитывая внутренние “зубцы” оболочки Pseudoschizaea, очевидно, что здесь нужен другой, не палинологический термин – например, ленточная, шнуровидная поверхность. В нашем случае “зубцы” проявляются как на наружной, так и на внутренней поверхностях оболочки Pseudoschizaea. Можно предположить, что развитие подобного типа оболочки с перпендикулярными поверхности слоями должно происходить несколько иначе, чем у споровых и семенных растений, у которых слои оболочки формируются параллельно поверхности. Однако на данный момент это невозможно доказать, и необходимы дополнительные данные для интерпретации такого типа оболочки.
Наиболее перспективные кандидаты для сравнения, которые рассматривались и предыдущими исследователями Pseudoschizaea – споры мхов, цисты водорослей, споры грибов и яйца беспозвоночных.
Сравнение изученной палиноморфы с мохообразными. Мохообразные (Bryophytes) включают в себя три основные группы: антоцеротовые, печеночники и настоящие мхи. Антоцеротовые – небольшая группа растений, продуцирующих споры средних размеров (10–80 мкм), в основном, одиночные [тетрады известны для единственного представителя рода Leiosporoceros Hässel, L. dussii (Stephani) Hässel de Menéndez], трилетные (монолетные известны для L. dussii). Скульптура может быть шипиковатая, папиллярная, шероховатая, бугорчатая, сетчатая, редко стриатная [у Hattorioceros striatisporus (J. Haseg.) J. Haseg.] или гладкая (у L. dussii). Экзоспорий обычно толстый, хотя есть представители с довольно тонким экзоспорием (0.4–1.0 мкм), гранулярный. Из представителей данной группы с Pseudoschizaea сравним только Hattorioceros striatisporus (J. Haseg.) J. Haseg. по признакам скульптуры оболочки, но споры данного вида значительно (почти в 3 раза) мельче, и ультраструктура их оболочки пока не изучена (Zhang et al., 2011).
Печеночники – очень разнообразная группа, продуцирует споры одиночные или в тетрадах, крупные, иногда мелкие (от 17 до 130 мкм), но всегда в коробочке имеются и споры, и элатеры (кроме сферокарповых). Тетрадный рубец обычно хорошо виден. Скульптура может быть очень разная, шиповатая и гранулярная, выросты часто очень причудливые, могут соединяться гребнями или формировать сеть. Спородерма толстая (если измерять с шипами), состоит из многих слоев, причем с самого начала формируется как ламеллятная, и эта ламеллятность всегда сохраняется в зрелых оболочках. Споры всех представителей этой группы резко отличаются от изученной палиноморфы Pseudoschizaea.
Настоящие мхи – самая обширная группа среди мохообразных. Споры мхов разнообразные, обычно одиночные, иногда в тетрадах, размеры от очень мелких до очень крупных (6–310 мкм), иногда с тетрадным рубцом. Поверхность спор представлена различными типами: от гладкой или гранулярной до бугорчатой и шипиковатой, или даже сетчато-столбиковой. Ультраструктура оболочек спор может быть сложной, многослойной, с участием ламеллятных, гомогенных и гранулярных слоев. Наиболее сложной ультраструктурой оболочек спор характеризуются базальные группы мхов (сфагновые, андреевые). Обычно спородерма довольно тонкая, представлена гранулами наружного периспория и гомогенным экзоспорием. Часто выражено утолщение эндоспориума на месте скрытой лезуры, которая не видна ни в СМ, ни в СЭМ. У перины часто встречается волнистая внутренняя поверхность, но перина всегда рыхлая, более или менее гранулярная, обычно не составляет сплошного слоя, выглядит как наваленные гранулы. Перина хорошо развита только у настоящих мхов. Больше всего внутренняя поверхность на срезах у Pseudoschizaea сходна с тем, что наблюдается у Andreaea Hedw. (Brown et al., 2015). Подобно Pseudoschizaea, у мхов иногда наблюдается поперечная исчерченность [напр., Oedipodium griffithianum (Dicks.) Schwägr.; Polevova, 2015] в наружном экзоспориуме, выглядящая как зазоры от слившихся гранул. Внутренний экзоспориум более толстый и гомогенный. В отличие от оболочек спор мхов, у Pseudoschizaea весь слой пронизан узкими полостями, которые производят впечатление слипшихся пластин, формирующих рельеф наружной и внутренней поверхностей.
Сравнение изученной палиноморфы с водорослями. Сведений об ультраструктуре спор грибов и цист водорослей значительно меньше. У водорослей рассматриваются в основном цисты и споры (имеющие разные названия у разных групп), гирогониты (ископаемая известковая оболочка харовых водорослей), редко клеточные стенки (в случае некоторых представителей зеленых водорослей). Большинство исследователей относят Pseudoschizaea к водорослям, вслед за М. Россиньол (Rossignol, 1962) и Ф. Тьегар и У. Франц (Thiergart, Frantz, 1962). Главным образом, рассматриваются Zygnemataceae (напр., Zippe, 1998; Scafati et al., 2009 и др.), в частности, Debarya Wittrock. Однако характер скульптуры и форма зигоспор Debarya резко отличаются от таковых у Pseudoschizaea, a внутреннее строение оболочки пока изучено не было (Van Geel, Van der Hammen, 1978; Head, 1992; Kołackzek et al., 2012). Также сходные по отдельным признакам общей морфологии клеточной оболочки и цист виды встречаются у представителей Chrysophyceae и Chlorellaceae. Ребристая поверхность спор встречается также среди представителей Oedogoniaceae и Characeae (Van Geel, Van der Hammen, 1978; Zippe, 1998; Tiss et al., 2019). Размеры цист и толщина оболочки также сравнимы с таковыми у Pseudoschizaea. Однако характер ребристости совершенно иной, чем у изученной палиноморфы, ультраструктура оболочки у изученных представителей также значительно отличается (Van Geel, Van der Hammen, 1978; Zippe, 1998; Domozych et al., 2010; Tiss et al., 2019). Скульптура поверхности цист, наиболее сходная с таковой у Pseudoschizaea, наблюдается у эвгленовых (напр., Hindak et al., 2000; Leander, Farmer, 2000). Размеры цист и толщина оболочки эвгленовых также сравнимы с таковыми у Pseudoschizaea. У эвгленовых, подобно Pseudoschizaea, наблюдаются перфорации на поверхности оболочки (pellicle pores, напр., Leander, Farmer, 2000), однако, у изученного вида эти перфорации не располагаются так же равномерно, как у эвгленовых, и их меньше. Ископаемая палиноморфа Moyeria Thusu со сходной скульптурой поверхности также сближается с эвгленовыми (Gray, Boucot, 1989). П. Строзе с соавт. (Strother et al., 2019) изучили ультраструктуру оболочки двух образцов Moyeria: Moyeria sp. и M. uticana Thusu, из верхнего силура Шотландии, и подтвердили отнесение подобных палиноморф к эвгленовым. Ультраструктура оболочки Moyeria, проиллюстрированная ими, однослойная, как и у Pseudoschizaea, однако, в ней отсутствует характерная поперечная исчерченность и “зубцы”, присущие изученной нами палиноморфе. Однако, учитывая разнообразие эвгленовых, показанное на современном материале, не исключается возможность того, что Pseudoschizaea могла также принадлежать к данной группе. Поперечная исчерченность оболочки, характерная для Pseudoschizaea, обнаружена у некоторых слоев оболочки ископаемого таксона Tasmanites Newton (Arouri et al., 2000; Vigran et al., 2008; Тельнова, 2012), предположительно принадлежавшего водорослям, однако, морфология Tasmanites разительно отличается от таковой Pseudoschizaea.
Сравнение изученной палиноморфы со спорами грибов. Споры грибов в подавляющем большинстве характеризуются значительно меньшими размерами, однако, у многих представителей обнаружена весьма сходная скульптура оболочки, особенно у аскоспор видов Neurospora Shear et B.O. Dodge (Ascomycota) (напр., Byrne, 1975). Также ребристая поверхность отмечена еще для ряда таксонов, напр., Ascobolus Pers., Caryospora Leger (Ascomycota) [см. небольшой обзор по спорам грибов с ребристой поверхностью y Дж. Грэй и А. Боукот (Gray, Boucot, 1989) и обзор ископаемых находок Ascomycota y T. Тэйлорa с соавт. (Taylor et al., 2015)]. Единственные приближающиеся по размерам к Pseudoschizaea споры грибов – урединиоспоры – характеризуются шипиковатой скульптурой (напр., Baka et al., 2004). Ультраструктура оболочки простая, обычно гомогенная, в общем, можно сказать, сходная с таковой у Pseudoschizaea у ряда представителей, однако нигде не наблюдается подобной поперечной исчерченности и внутренней волнистой поверхности.
Сравнение изученной палиноморфы с яйцами беспозвоночных. Среди беспозвоночных наиболее перспективными для сравнения с исследованной палиноморфой являются яйца представителей Collembola, Nematoda, Trematoda и Rotifera. Ультраструктура оболочки яиц известных представителей делает сравнение с Rotifera и Collembola мало результативным, ввиду существенного различия их строения; размеры яиц представителей этих групп также часто крупнее, и их скульптура демонстрирует иное строение (напр., Gilbert, Wurdack, 1978; Wurdack et al., 1978). Ультраструктура яиц у ряда таксонов червей (особенно, Trematoda), хотя в основном, гомогенная, но сходной толщины, и размеры яиц также сопоставимы с размерами Pseudoschizaea (напр., Bird, McClure, 1976; Swiderski et al., 2010, 2014; Conn et al., 2018). Плохо сохранившиеся остатки какого-то слоя ковнутри от оболочки Pseudoschizaea также сходны с гранулярным слоем наружной оболочки у представителей червей. Однако поверхность, наблюдаемая обычно у покоящихся яиц червей, в отличие от Pseudoschizaea, гладкая.
* * *
Автофлуоресценция также не смогла помочь в уточнения интерпретации Pseudoschizaea. Все палиноморфы одинаково слабо светятся, в отличие от ископаемых кутикул, что не противоречит сопоставлениям как со спорами высших растений, так и с водорослями. Возможно ожидать, что ископаемые оболочки яиц беспозвоночных будут отличаться от палиноморф не меньше кутикул.
Таким образом, выявлены сходные отдельные признаки скульптуры нашей палиноморфы у одного представителя мхов и целого ряда грибов; сходные черты общей морфологии с цистами ряда водорослей. Среди водорослей и мхов есть сходные черты ультраструктуры оболочки цист и спор. На настоящий момент наименее противоречиво выглядит гипотеза принадлежности Pseudoschizaea к водорослям, однако, основанная не на сравнении с ранее предложенными Zygnemataceae, а с эвгленовыми.
Список литературы
Tельнова O.П. Морфология и ультраструктура девонских празинофитов (Chlorophyta) // Палеонтол. журн. 2012. № 5. P. 99–105.
Arouri K.R., Greenwood P.F., Walter M.R. Biological affinities of Neoproterozoic acritarchs from Australia: microscopic and chemical characterization // Organic Geochem. 2000. V. 31. P. 75–89.
Avanzini M., Pibelli D., Mietto P. et al. Lower Jurassic (Hettangian–Sinemurian) dinosaur track megasites, southern Alps, Northern Italy // The Triassic – Jurassic Terrestrial Transition / Eds. Harris J.D., Lucas S.G., Spielmann J.A. Albuquerque: New Mexico Museum of Natur. Hist. and Sci. Bull., 2006. P. 207–216.
Baka Z.A.M., Alwadie H.M., Mostafa Y.S. Comparative morphology of uredinia and urediniospores of six Puccinia species parasitic on Poaceae in Saudi Arabia // Arch. Phytopathol. Plant Protect. 2004. V. 37. № 2. P. 89–101.
Bird A.F., McClure M.A. The tylenchid (Nematoda) egg shell: structure, composition and permeability // Parasitology. 1976. V. 72. P. 19–28.
Bosellini A., Broglio Loriga C. I ‘‘Calcari Grigi’’ di Rotzo (Giurassico Inferiore, Altopiano di Asiago) e loro inquadramento nella paleogeografia e nella evoluzione tettonico-sedimentaria delle Prealpi venete // Ann. Univ. Ferrara (Sez. Sci. Geol. e Paleontol.). 1971. V. 5. P. 1–61.
Brown R.C., Lemmon B.E., Shimamura M. et al. Spores of relictual bryophytes: Diverse adaptations to life on land // Rev. Palaeobot. Palynol. 2015. V. 216. P. 1–17.
Byrne B.J. Freeze-etch and thin section studies of Neurospora crassa ascospores // Protoplasma. 1975. V. 84. P. 357–377.
Christopher R.A. Morphology and taxonomic status of Pseudoschizaea Thiegart and Frantz ex R. Potonié emend. // Micropaleontology. 1976. V. 22. № 2. P. 143–150.
Clari P. Caratteristiche sedimentologiche e paleontologiche di alcune sezioni dei Calcari Grigi del Veneto // Mem. Ist. Geol. e Miner. Univ. Padova. 1975. V. 31. P. 1–63.
Conn D.B., Swiderski Z., Miquel J. Ultrastructure of digenean trematode eggs (Platyhelminthes: Neoophora): a review emphasizing new comparative data on four European Microphalloidea // Acta Parasitol. 2018. V. 63. № 1. P. 1–14.
Domozych D.S., Sørensen I., Pettolino F.A. et al. The cell wall polymers of the charophycean green alga Chara coralline: immunobinging and biochemical screening // Int. J. Plant Sci. 2010. V. 171. № 4. P. 345–361.
Gavrilova O., Zavialova N., Tekleva M., Karasev E. Potential of CLSM in studying some modern and fossil palynological objects // J. Microsc. 2018. V. 269. № 3. P. 291–309.
Gilbert J.J., Wurdack E.S. Species-specific morphology of resting eggs in the Rotifer Asplanchna // Trans. Amer. Microscop. Soc. 1978. V. 97. № 3. P. 330–339.
Gray J., Boucot A.J. Is Moyeria a euglenoid? // Lethaia. 1989. V. 22. P. 447–456.
Head M.J. Zygospores of the Sygnemataceae (Division Chlorophyta) and other freshwater algal spores from the Uppermost Pliocene St. Erth Beds of Cornwall, Southwestern England // Micropaleontology. 1992. V. 38. № 3. P. 237–260.
Hesse M., Halbritter H., Weber M. et al. Pollen terminology—an illustrated handbook. N. Y.: Springer, 2009. 266 p.
Hindak A., Wolowdki K., Hindakova A. Cysts and their formation in some neustonic Euglena species // Ann. Limnol. 2000. V. 36. № 2. P. 83–93.
Kołackzek P., Karpińska-Kołackzek M., Worobiec E., Heise W. Debarya glyptosperma (De Bary) Wittrock 1872 (Zygnemataceae, Chlorophyta) as a possible airborne alga—a contribution to its palaeoecological interpretation // Acta Palaeobot. 2012. V. 52. № 1. P. 139–146.
Leander B.S., Farmer M.A. Comparative morphology of the euglenid pellicle. I. Patterns of strips and pores // J. Eukaryot. Microbiol. 2000. V. 47. № 5. P. 469–479.
Matveyeva N.P., Polevova S.V., Smirnova A.V., Yermakov I.P. Sporopollenin accumulation in Nicotiana tabacum L. microspore wall during its development // Cell Tiss. Biol. 2012. V. 6. P. 293–301.
Milanesi C., Vignani G., Ciampolini F. et al. Ultrastructure and DNA sequence analysis of single Concentricystis cells from Alta Val Tiberina Holocene sediment // J. Archaeol. Sci. 2006. V. 33. P. 1081–1087.
Neri M., Kustatscher E., Roghi G. Megaspores from the Lower Jurassic (Pliensbachian) Rotzo Formation (Monti Lessini, northern Italy) and their palaeoenvironmental implications // Palaeobiodiv. Palaeoenv. 2018. V. 98. P. 97–110.
Neri M., Roghi G., Ragazzi E., Papazzoni C.A. Lower Jurassic amber from the Rotzo Formation (Monti Lessini, northern Italy) // Geobios. 2017. V. 50. P. 49–63.
Parkinson B.M. Morphological and ultrastructural variations in Schizaea pectinate (Schizaeaceae: Pteridophyta) // Bothalia. 1994. V. 24. № 2. P. 203–210.
Parkinson B.M. Spore wall development in Schizaea pectinate (Schizaeaceae: Pteridophyta) // Grana. 1995. V. 34. P. 217–228.
Polevova S.V. Sporoderm ultrastructure of Oedipodium griffithianum (Oedipodiopsida, Bryophyta) // Arctoa. 2015. V. 23. P. 389–400.
Ramos Giacosa J.P., Barakat M.C. Spore morphology and wall ultrastructure of Actinostachys pennula (Sw.) Hook. and A. subtrijuga (Mart.) C. Presl. (Schizaeaceae) // Palynology. 2018. V. 42. P. 483–491.
Ramos Giacosa J.P., Morbelli M.A., Giudice G.E. Morphology and ultrastructure of Schizaea fistulosa (Schizaeaceae) spores from Chile // Bol. Soc. Argent. Bot. 2015. V. 50. № 1. P. 17–22.
Rossignol M. Analyse pollenique de sediments marins Quaternaires an Israel. 2. Sediments Pleistocènes // Pollen et Spores. 1962. V. 4. P. 121–148.
Scafati L., Melendi D.L., Volkheimer W. A Danian subtropical lacustrine palynobiota from South America (Bororo Formation, San Jorge Basin, Patagonia–Argentina) // Geol. Acta. 2009. V. 7. № 1–2. P. 35–61.
Scott L. Environmental implications and origin of microscopic Pseudoschizaea Thiergart and Frantz ex R. Potonie emend. in sediments // J. Biogeogr. 1992. V. 19. № 4. P. 349–354.
Skog J.E. The relationship of the fossil fern Schizaeaopsis Berry to modern genera in the Schizaeaceae // Amer. Fern J. 1993. V. 83. № 1. P. 20–29.
Strother P.K., Taylor W.A., van de Schootbrugge B. et al. Pellicle ultrastructure demonstrates that Moyeria is a fossil euglenid // Palynology. 2019. published online https://doi.org/10.1080/01916122.2019.1625457
Swiderski Z., Bakhoum A.J.S., Mlocicki D., Miquel J. Ultrastructural studies on egg envelopes surrounding the miracidia of Mediogonimus jourdanei Mas-Coma et Rocamora, 1978 (Digenea, Microphalloidea, Prosthogonimidae) // Acta Parasitol. 2010. V. 55. № 3. P. 245–253.
Swiderski Z., Poddubnaya L.G., Zhokhov A.E. et al. Ultrastructural evidence for completion of the entire miracidial maturation in intrauterine eggs of the digenean Brandesia turgida (Brandes, 1888) (Plagiorchiida: Pleurogenidae) // Parasitol. Res. 2014. V. 113. P. 1103–1111.
Taylor T.N., Krings M., Taylor E.L. Fossil fungi. Amsterdam: Acad. Press, 2015. 382 p.
Thiergart F., Frantz U. Some spores and pollen grains from a Tertiary brown coal deposit in Kashmir // Palaeobotanist. 1962. V. 10. P. 84–86.
Tiss L., Trabelsi K., Kamoun F. et al. Middle Jurassic charophytes from southern Tunisia: implications on evolution and paleobiogeography // Rev. Paleobot. Palynol. 2019. V. 263. P. 65–84.
Tryon A.F., Lugardon B. Spores of the Pteridophyta: Surface, Wall Structure, and Diversity Based on Electron Microscope Studies. N. Y.: Springer, 1991. 648 p.
Van Geel B., Van der Hammen T. Zygnemataceae in Quaternary Colombian sediments // Rev. Paleobot. Palynol. 1978. V. 25. P. 377–392.
Vigran J.O., Mork A., Forsberg A.W. et al. Tasmanites algae—contributors to the Middle Triassic hydrocarbon source rocks of Svalbard and the Barents Shelf // Polar Res. 2008. V. 27. P. 360–371.
Wesley A. Contribution to the knowledge of the flora of the Grey Limestone of Veneto. Part I // Mem. Ist. Geol. e Mineral. Univ. Padova. 1956. V. 19. P. 1–69.
Wesley A. Contribution to the knowledge of the flora of the Grey Limestone of Veneto. Part II // Mem. Ist. Geol. e Mineral. Univ. Padova. 1958. V. 21. P. 1–57.
Wurdack E.S., Gilbert J.J., Jagels R. Fine structure of the resting eggs of the Rotifers Brachionus calyciflorus and Asplanchna sieboldin // Trans. Amer. Microscop. Soc. 1978. V. 97. № 1. P. 49–72.
Zavialova N.E., Tekleva M.V., Polevova S.V., Bogdanov A.G. Electron microscopy for morphology of pollen and spores. Moscow: RIPOL Classic Press, 2018. 334 p.
Zhang L., Zhou L., Li J. The genus Hattorioceros (Notothyladaceae) new to China // The Bryologist. 2011. V. 114. № 1. P. 190–193.
Zippe P.A. Freshwater algae from the Mattagami Formation (Albian), Ontario: paleoecology, botanical affinities, and systematic taxonomy // Micropaleontol. 1998. V. 44. Suppl. 1. P. 1–78.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал