

Палеонтологический журнал, 2022, № 1, стр. 97-106
Пещерные медведи (Ursus spelaeus sensu lato) Урала
Д. О. Гимранов a, *, П. А. Косинцев a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: djulfa250@rambler.ru
** E-mail: kpa@ipae.uran.ru
Поступила в редакцию 08.06.2021
После доработки 10.06.2021
Принята к публикации 15.06.2021
- EDN: GMFVUO
- DOI: 10.31857/S0031031X22010068
Аннотация
Приведены обобщенные результаты восьмидесятилетних исследований биологии, распространения, времени обитания и вымирания, таксономии и филогении, а также взаимодействия с человеком большого [Ursus (Spelaearctos) kanivetz Vereshchagin, 1973] и малого [Ursus (Spelaearctos) ex gr. savini–rossicus] пещерных медведей на Урале.
Пещерные медведи [Ursus (Spelaearctos) spp.] – группа типичных представителей позднеплейстоценового (мамонтового) фаунистического комплекса, широко распространенных в Европе и Северной Азии (Барышников, 2007; Sher et al., 2011). Вплоть до начала XXI в. считалось, что в позднем плейстоцене существовало два вида пещерных медведей: большой пещерный медведь Ursus spelaeus Rosenmüller, 1794 и малый пещерный медведь U. savini Andrews, 1922, или U. rossicus Borissiak, 1930 (Kurtèn, 1995; Барышников, 2007). В настоящее время по молекулярным данным (Barlow et al., 2021) в группе больших пещерных медведей различают три вида: U. spelaeus и U. eremus Rabeder, Hofreiter, Nagel et Withalm, 2004 (распространены в Западной Европе), а также U. kanivetz Vereshchagin, 1973 (распространен в Центральной и Восточной Европе и на Урале; Knapp et al., 2009; Rabeder et al., 2011; Stiller et al., 2014; Baryshnikov, Puzachenko, 2019, 2020). В группе малых пещерных медведей выделяют U. savini (Западная Европа) и U. rossicus (Северный Кавказ, Восточная Европа, Урал, Алтай, Западная и Восточная Сибирь) (Борисяк, 1930; Spassov et al., 2017; Baryshnikov, Puzachenko, 2019, 2020; Barlow et al., 2021).
На Урале в позднем плейстоцене совместно обитали большой и малый пещерные медведи. В это время здесь существовали самые северные популяции обоих видов и проходила восточная граница ареала большого пещерного медведя. Здесь их ареалы имели наибольшую протяженность с севера на юг – более 1300 км. Они проходили через районы с разными природно-климатическими характеристиками. Все это обуславливает важность Урала для изучения общих проблем микроэволюции, филогеографии, экологии и биологии большого и малого пещерных медведей.
Пещерные медведи Урала изучаются более 80 лет; за это время накоплен огромный объем данных, отражающих как частные, так и общие вопросы их биологии, морфологии, филогенеза и взаимодействия с человеком. Эти данные опубликованы в более чем 160 работах и обобщены в настоящей публикации.
ИСТОРИЯ ИЗУЧЕНИЯ ПЕЩЕРНЫХ МЕДВЕДЕЙ УРАЛА
Впервые кости пещерных медведей были найдены в Уньинской пещере на Северном Урале Э. Гофманом в 1847 г. (Гофман, 1856), но их видовая принадлежность была определена только в XX в. Первые научные коллекции костей пещерных медведей были получены в ходе раскопок пещер Южного Урала С.Н. Бибиковым в 1937 г. (Громов, 1948).
В истории изучения пещерных медведей Урала можно выделить несколько этапов. На первом этапе, который длился с конца 1940-х до конца 1980-х гг., происходило накопление данных о местонахождениях костей пещерных медведей. Были открыты и описаны десятки местонахождений на Северном, Среднем и Южном Урале (Громов, 1948; Карачаровский, 1951; Верещагин, 1957; Верещагин, Кузьмина, 1962; Гуслицер, Канивец, 1965; Кузьмина, 1971, 1975; Гуслицер, Павлов, 1987). В это время были проведены первые исследования изменчивости и был определен таксономический статус обитавших на Урале пещерных медведей (Кузьмина, 1971; Верещагин, 1973). Установлено, что на Урале обитали две формы пещерных медведей: большой пещерный медведь Ursus spelaeus kanivetz Vereshchagin, 1973 и малый пещерный медведь Ursus uralensis Vereshchagin, 1973 (Верещагин, 1973).
Второй этап длился с конца 1980-х гг. до конца 2000-х гг. В этот период начато целенаправленное изучение экологии пещерных медведей Урала (Косинцев, 1988; Смирнов и др., 1990; Косинцев, Воробьев, 2001). Появляются первые радиоуглеродные даты, полученные по костям пещерных медведей (Косинцев и др., 2003; Барышников, 2007; Pacher, Stuart, 2009). Существенно увеличивается число работ по изучению изменчивости и таксономии пещерных медведей Урала (Vereschagin, Baryshnikov, 2000; Baryshnikov, Foronova, 2001; Кузьмина, 2002; Барышников, 2003, 2007; Сатаев, 2006; Baryshnikov, 2006). Продолжают публиковаться данные о местонахождениях с остатками пещерных медведей (Смирнов и др., 1990; Сатаев, 1995; Яковлев и др., 2000; Улитко, 2003; Косинцев, Сатаев, 2005; Ражев и др., 2005). К концу этого периода были выявлены морфологические характеристики малого пещерного медведя, а также получены данные о половом и возрастном составе популяций большого и малого пещерных медведей по репрезентативным материалам из нескольких местонахождений Среднего и Южного Урала.
Последний этап начался в конце 2000-х гг. и продолжается по настоящее время. В этот период активно используются молекулярно-генетические методы (Stiller et al., 2009, 2010, 2014; Knapp et al., 2009; Gretzinger et al., 2019; Knapp, 2019; Barlow et al., 2021). В результате анализа ядерной ДНК установлена таксономическая самостоятельность и определено время дивергенции филетических линий малого и большого пещерных медведей Урала, а также оценено их морфологическое своеобразие. Для изучения изменчивости медведей начинают широко использоваться многомерные статистические методы (Baryshnikov, Puzachenko, 2011, 2017, 2019, 2020; Baryshnikov et al., 2018, 2019). Получены первые данные о взаимодействии человека и пещерных медведей Урала (Широков и др., 2011; Котов и др., 2020; Гимранов и др., 2021а). Продолжаются описания новых местонахождений остатков пещерных медведей (Kosintsev et al., 2016; Фадеева и др., 2019; Юрин, 2020; Danukalova et al., 2020; Gimranov, Kosintsev, 2020; Fadeeva et al., 2020; Гимранов и др., 2021б; Косинцев и др., 2021).
РАСПРОСТРАНЕНИЕ ПЕЩЕРНЫХ МЕДВЕДЕЙ НА УРАЛЕ
Практически все остатки пещерных медведей на Урале происходят из местонахождений пещерного типа. Имеется только одно местонахождение открытого типа – стоянка среднего палеолита Богдановка на Южном Урале (Широков и др., 2011), в которой найдены остатки малого пещерного медведя. На Полярном Урале остатков пещерных медведей не обнаружено (Bachura, Kosintsev, 2007).
На Северном Урале насчитывается шесть местонахождений (рис. 1, а, б) с костями пещерных медведей (Кузьмина, 1971; Косинцев, Воробьев, 2001). Самым северным местонахождением остатков большого пещерного медведя является грот Большая Дроватница (63°57′ с.ш., 57°37′ в.д.) (Кузьмина, 1971). В регионе известно одно местонахождение малого пещерного медведя – Медвежья пещера (62°05′ с.ш., 58°05′ в.д.) (Гимранов и др., 2021б). Находки костей пещерного медведя в гроте Шайтанский на восточном склоне Северного Урала (Косинцев, Бородин, 1990) не подтвердились.
Рис. 1.
Карта-схема местонахождений Урала с остатками пещерных медведей: а – местонахождения большого пещерного медведя, б – местонахождения малого пещерного медведя. Полужирным шрифтом выделены номера и названия местонахождений, где обнаружены остатки только малого пещерного медведя.
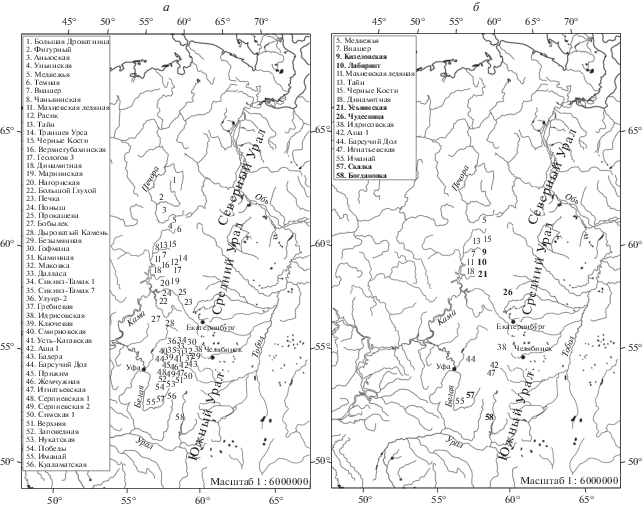
На Среднем Урале имеются 18 местонахождений с костями большого пещерного медведя и четыре местонахождения с малым пещерным медведем (рис. 1, а, б). Находки пещерных медведей ранее были указаны в пещерах Язьвинская, Першинская (Косинцев, Воробьев, 2001) и гроте Безымянном (Петрин, Смирнов, 1977; Косинцев, Воробьев, 2001). При пересмотре этих материалов нами установлено, что костные остатки из этих местонахождений в действительности принадлежат бурому медведю.
На Южном Урале насчитываются 28 местонахождений с остатками большого пещерного медведя и два местонахождения с малым пещерным медведем (рис. 1, а, б). Ранее были указаны находки пещерных медведей в пещерах Бейдинская и Улуир 2 (Юрин, 2011), Смеловская 2 (Кузьмина, 2000; Косинцев, Воробьев, 2001; Kosintsev, 2007; Юрин, 2011) и Капова (Кузьмина, Абрамсон, 1997; Косинцев, Воробьев, 2001). Ревизия этих материалов показала, что их невозможно идентифицировать ближе, чем до рода Ursus. Они могут принадлежать как пещерному, так и бурому медведю.
Анализ географического распространения находок указывает на присутствие большого пещерного медведя только в местонахождениях западного склона Урала, тогда как малый пещерный медведь найден на обоих склонах (рис. 1). Не вызывает сомнения, что Уральские горы не могли быть физическим препятствием для перехода большого пещерного медведя на восточный склон, поэтому его отсутствие на восточном склоне пока не имеет обоснованных объяснений. Оба вида пещерных медведей – растительноядные животные (Naito et al., 2020), в связи с чем симпатрия их ареалов может косвенно указывать на существенные различия их рационов питания. Можно предположить, что на восточном склоне Урала и далее в Сибири ряд основных пищевых видов растений большого пещерного медведя отсутствовал, или же продуктивность их ценопопуляций была недостаточно высокой. Это же может объяснить и ограничение продвижения обоих видов на север.
ТАКСОНОМИЯ И ФИЛОГЕНИЯ ПЕЩЕРНЫХ МЕДВЕДЕЙ УРАЛА
В настоящее время пещерных медведей Урала относят к двум видам: большому U. kanivetz и малому U. ex gr. savini–rossicus. Они хорошо различаются по строению бакулюма (Верещагин, 1973), по размерам и пропорциям черепа (Baryshnikov, Puzachenko, 2011), нижней челюсти (Baryshnikov et al., 2018), щечных зубов (Vereschagin, Baryshnikov, 2000; Baryshnikov, Puzachenko, 2019, 2020; Гимранов и др., 2021б) и метаподий (Baryshnikov, Puzachenko, 2017). Достоверность этих различий недавно была подтверждена анализом ядерной ДНК (Barlow et al., 2021).
С течением времени представления о таксономической принадлежности пещерных медведей с Урала неоднократно менялись. Первые находки были отнесены к большому пещерному медведю U. spelaeus (Громов, 1948). Все последующие находки крупной формы пещерного медведя также относили к этому виду. В 1973 г. по материалам из Медвежьей пещеры был описан самостоятельный подвид большого пещерного медведя U. spelaeus kanivetz (Верещагин, 1973). Это название использовалось и в дальнейшем (Барышников, 1981, 2003, 2007; Абрамов, Барышников, 1990; Baryshnikov, Puzachenko, 2011). Позднее по материалам из пещеры Тайн был описан подвид U. s. bliznetshovi Kuzmina, 2002 (Кузьмина, 2002), который впоследствии был сведен в младшие синонимы U. s. odessanus von Nordmann, 1858, наряду с U. spelaeus kanivetz (Барышников, 2003). Впоследствии самостоятельность подвида U. s. kanivetz была восстановлена (Барышников, 2007).
Изучение митохондриальной ДНК по образцам из пещеры Медвежья показало, что большой пещерный медведь с Северного Урала относится к гаплогруппе “ingressus” (Knapp et al., 2009; Stiller et al., 2009), представители которой ранее были выделены в самостоятельный вид U. ingressus Rabeder et al., 2004 (Rabeder et al., 2004). Позднее это подтвердилось новыми исследованиями митохондриальной ДНК медведей (Stiller et al., 2014; Gretzinger et al., 2019). Пещера Медвежья представляет собой типовое местонахождение для U. spelaeus kanivetz Vereshchagin, 1973. Таким образом, согласно “Международному кодексу зоологической номенклатуры”, последнее название имеет приоритет, и вид должен быть обозначен как U. kanivetz Vereshchagin, 1973 (=U. ingressus Rabeder et al., 2004) (Baryshnikov, Puzachenko, 2017). Пещерные медведи из местонахождений “грот Большой Глухой” на Среднем Урале и “Серпиевская пещера” на Южном Урале также относятся к гаплогруппе “ingressus” (Stiller et al., 2009). Таким образом, все большие пещерные медведи Урала относятся к U. kanivetz. Анализ ядерной ДНК подтвердил генетическое своеобразие и видовую самостоятельность большого пещерного медведя Урала (Barlow et al., 2021).
Малый пещерный медведь из Кизеловской пещеры на Среднем Урале первоначально был отнесен к U. rossicus Borissiak, 1930 (Верещагин, 1959), а позднее был выделен в самостоятельный вид U. uralensis (Верещагин, 1973). В дальнейшем эта форма обозначалась либо как U. rossicus uralensis (Барышников, 1981; Верещагин, 1982), либо как U. rossicus (Vereschagin, Baryshnikov, 2000; Baryshnikov, Foronova, 2001; Кузьмина, 2002; Косинцев, 2003; Яковлев и др., 2005). В 2003 г. вид U. rossicus Borissiak, 1930 был сведен в синонимы U. savini Andrews, 1922, а подвид U. r. uralensis Vereshchagin, 1973, соответственно, стал обозначаться как U. savini uralensis (Барышников, 2003). На протяжении ряда последующих лет остатки малого пещерного медведя с территории Урала относили к U. savini Andrews, 1922 (Косинцев, Сатаев, 2005; Барышников, 2007; Kosintsev, 2007; Широков и др., 2011; Sher et al., 2011; Kosintsev, Bachura, 2013; Kosintsev et al., 2016; Косинцев, 2019; Gimranov, Kosintsev, 2020; Fadeeva et al., 2020; Danukalova et al., 2020; Силаев и др., 2020). В то же время, другие исследователи продолжали использовать название U. rossicus, основываясь, в том числе, и на молекулярно-генетических данных (Маркова и др., 2008; Pacher, Stuart, 2009; Danukalova et al., 2009; Baryshnikov, Puzachenko, 2011, 2017, 2019, 2020; Фадеева и др., 2011; Stiller et al., 2014; Baryshnikov et al., 2018; Бачура, Косинцев, 2019; Knapp, 2019; Puzachenko et al., 2020; Barlow et al., 2021). До проведения ревизии морфологических и молекулярно-генетических данных по малым пещерным медведям Урала, Восточной и Западной Европы мы относим малого пещерного медведя Урала к группе Ursus ex gr. savini–rossicus (Котов и др., 2020; Гимранов и др., 2021а, б; Павлова и др., 2021).
Неопределенное таксономическое положение имеет мелкая форма пещерного медведя из пещеры Верхней на Южном Урале. Ее определяли как Spelaearctos cf. rossicus (Sataev, 1996), S. rossicus (Яковлев и др., 2005) или Ursus (Spelaearcos) cf. spelaeus deningeroides Mottl, 1964 (Р.М. Сатаев, личн. сообщ.). В итоге было сделано заключение о невозможности точного видового определения, и эта форма была обозначена как U. (Spelaearcos) sp. (Сатаев, 2008). Тем не менее, в ряде публикаций эта находка включена в списки фаун как U. cf. deningeri hercynicus (Danukalova et al., 2008), U. savini (Kosintsev et al., 2016) и U. cf. deningeri (Danukalova et al., 2020).
Представления о филогенетических отношениях пещерных медведей Урала первоначально строились на результатах морфологического анализа. Большой пещерный медведь Урала по результатам многомерного анализа размеров черепа (Baryshnikov, Puzachenko, 2011), нижней челюсти (Baryshnikov et al., 2018), верхних зубов (Baryshnikov, Puzachenko, 2019), нижних зубов (Baryshnikov, Puzachenko, 2020) и метаподий (Baryshnikov, Puzachenko, 2017) объединяется с другими выборками большого пещерного медведя (U. spelaeus s.l.). По результатам анализа размеров зубов U. rossicus из пещеры Кизеловской наряду с медведями из Краснодара и Южной Сибири, а также U. savini Andrews, 1922 из местонахождения “Bacton Forest Bed” объединились в одну кладу, отдельную от U. spelaeus (Baryshnikov, Foronova, 2001). Объединенная выборка малого пещерного медведя, с включением выборки уральской формы, по результатам анализа размеров черепа, нижней челюсти, зубов и метаподий (Baryshnikov, Puzachenko, 2011, 2017, 2019, 2020; Baryshnikov et al., 2018) формирует самостоятельную кладу, отдельную от других пещерных медведей.
В последние годы по результатам анализа митохондриальной ДНК были реконструированы следующие группировки медведей: клада “U. spelaeus”, включающая в себя “spelaeus”, “ladinicus”, “eremus”, а также клада U. kanivetz (=U. ingressus), включающая в себя U. rossicus с Урала (Stiller et al., 2014; Knapp, 2019). Последнее плохо согласуется с морфологическими данными.
Анализ ядерной ДНК медведей Европы и Урала дал совершенно другую картину филогенетических взаимоотношений пещерных медведей (Barlow et al., 2021). Были выявлены две клады, одна из которых включает в себя всех больших пещерных медведей (U. spelaeus s.l.) Европы и Урала, а вторая – только малого пещерного медведя (U. rossicus) из пещеры Кизеловской на Урале. Малый пещерный медведь Урала представляет собой глубоко дивергировавшую изолированную филогенетическую линию и третью крупную группу пещерных медведей, наряду с европейской (U. spelaeus s.l.) и кавказской (U. kudarenis s.l.). Анализ ядерного генома подтверждает валидность таксона U. kanivetz, который реконструируется как сестринская группа к европейским линиям spelaeus и ingressus. Результаты анализа ядерного генома полностью подтверждают результаты морфологического анализа (Baryshnikov, Puzachenko, 2011, 2017, 2019, 2020; Baryshnikov et al., 2018). Таким образом, филогения пещерных медведей, построенная ранее по результатам анализа митохондриальной ДНК, представляется некорректной.
В последнее время получены оценки времени дивергенции разных форм пещерных медведей. Медианная оценка дивергенции малого пещерного медведя Урала (U. rossicus) и большого пещерного медведя (U. spelaeus s.l.) по ядерной ДНК составляет около 880 тыс. лет назад, а отделение U. kanivetz от группы “spelaeus–ingressus”, согласно этим данным, произошло около 400 тыс. л. н. (Barlow et al., 2021).
БИОЛОГИЯ ПЕЩЕРНЫХ МЕДВЕДЕЙ УРАЛА
Наиболее репрезентативные данные получены по биологии большого пещерного медведя. По выборкам из нескольких местонахождений определены половая и возрастная структуры части популяции, погибшей во время зимней спячки. Определение пола проводилось по размерам костей (пещеры Медвежья, Тайн, Геологов 3 и Аша 1) и клыков (пещера Игнатиевская). Соотношение самцов и самок в пещере Тайн составляет примерно 1 : 3, в пещерах Медвежьей, Геологов 3 и Игнатиевской – 1 : 2 и в пещере Аше 1 – 1 : 1 (Косинцев, Воробьев, 2001; Кузьмина, 2002). Во всех выборках, кроме выборки из Аши 1, соотношения полов достоверно отличаются от соотношения 1 : 1. Очевидно, что в большинстве случаев самки чаще погибали во время спячки, чем самцы. Этому дано объяснение на основании аналогии с биологией бурого медведя, у которого беременные самки и самки с детенышами обычно залегают в спячку раньше самцов и прохолостовавших самок, занимая наиболее удобные места для спячки. Вероятно, такая черта поведения была свойственна и пещерным медведям. Если это так, то пещерные медведи залегали в спячку поодиночке (или же самка с детенышами), а не группами, иначе различия соотношения самцов и самок были бы менее выражены (Косинцев, Воробьев, 2001).
Соотношение возрастных групп оценивалось различными критериями. В выборках из пещер Тайн и Геологов 3 оно было определено по размерам костей и прирастанию эпифизов, в результате чего было выделено несколько возрастных групп (0–1 год, 1–2 года, 2–3 года, 3–4 года, старше 4 лет). В обеих пещерах доминируют особи в возрасте 1–2 и 2–3 лет (58 и 75% соответственно), новорожденные особи (0–1 год) составляют 9.5 и 2.3%, а взрослые особи (старше 4 лет) – 18 и 34%. В пещерах Игнатиевской и Аше 1 возрастной состав был определен по степени стертости коронки резцов (I1-2 и i1-2), для которых было выделено семь стадий стирания (Смирнов и др., 1990). В пещере Аше 1 среди погибших доминировали полувзрослые (21%) и взрослые особи (63%). В трех древних слоях пещеры Игнатиевской преобладали молодые (19–28%) и полувзрослые (22–26%) особи, а в позднем слое доминировали молодые особи (47%). Различия между выборками из пещер Игнатиевской и Аши 1 статистически достоверны (Смирнов и др., 1990), при этом причины таких существенных различий возрастной структуры неясны.
Паразитологический анализ копролитов большого пещерного медведя из Игнатиевской пещеры показал, что он был инвазирован нематодой Baylisascaris transfuga Rudolphi, 1819 (Сивкова, Косинцев, 2021), специфичной для семейства Ursidae.
Данных по биологии малого пещерного медведя получено меньше. На основе анализа размеров костей установлено, что соотношение самцов и самок в пещере Иманай составляет 3 : 1 (Gimranov, Kosintsev, 2020), а в Кизеловской – 1 : 2 (Vereschagin, Baryshnikov, 2000). Численность взрослых особей в пещере Иманай составляет 84% (Gimranov, Kosintsev, 2020), в Кизеловской – 52% (Верещагин, 1982). Получены первые данные о возрасте и сезоне гибели единичных особей малого пещерного медведя по регистрирующим структурам в зубах (Прилепская, Барышников, 2019; Гимранов и др., 2021а).
По результатам морфологического анализа отмечено, что малый пещерный медведь был формой еще более пастбищного типа, чем большой пещерный медведь (Верещагин, 1973), что подтверждается данными анализа изотопных подписей C13 и N15 (Силаев и др., 2020). Патологические изменения отмечены на единичных костях пещерных медведей Урала (Косинцев, Воробьев, 2001).
Выявленные характеристики биологии большого и малого медведей Урала существенно различаются не только между видами, но и для отдельных изученных палеопопуляций. Подобные значительные внутривидовые различия пока не находят объяснения.
ХРОНОЛОГИЯ СУЩЕСТВОВАНИЯ ПЕЩЕРНЫХ МЕДВЕДЕЙ НА УРАЛЕ
Самые древние остатки большого и малого пещерных медведей на Урале найдены в Игнатиевской (раскоп V, слой 10) и Серпиевской 1 (слой 3) пещерах (Смирнов и др., 1990). Они входили в состав Серпиевской фауны, датируемой концом среднего [конец морской изотопной стадии (МИС) 6] или началом позднего (МИС 5е) плейстоцена (Фадеева и др., 2019; Danukalova et al., 2020). Остатки обоих видов найдены в местонахождениях последнего межледниковья (микулинское, казанцевское, эемское, МИС 5е; Fadeeva et al., 2020). За исключением Серпиевской фауны, остатки пещерных медведей на Урале найдены в фаунах, которые включают в себя только виды, типичные для позднеплейстоценового (мамонтового) фаунистического комплекса (Kosintsev et al., 2016).
Более точно время обитания и вымирания пещерных медведей на Урале позволяют определить радиоуглеродные даты. До получения радиоуглеродных дат по костям пещерных медведей время вымирания определялось в диапазоне 104–120 тыс. л. н. (Верещагин, 1971; Кузьмина, 1971, 2002; Косинцев, Воробьев, 2001). К настоящему времени по костям большого пещерного медведя из местонахождений Урала получено 26 конечных радиоуглеродных дат и 11 запредельных, а по костям малого пещерного медведя – 14 конечных дат и две запредельные даты (Косинцев и др., 2003; Барышников, 2007; Pacher, Stuart, 2009; Косинцев и др., 2020). Среди дат, полученных по костям большого пещерного медведя, есть несколько относительно поздних. Две даты из пещеры Виашер (19550 ± 230, SOAN-4526; 22650 ± 670, SOAN-4515) и одна из пещеры Верхней (22 750 ± ± 1210, LU-3714) (Косинцев и др., 2003; Барышников, 2007) совпадают с самыми поздними датами, полученными по костям большого пещерного медведя Европы (Baca et al., 2016; Gretzinger et al., 2019). Вероятно, вымирание большого пещерного медведя произошло синхронно на всем его ареале в течение последнего ледникового максимума (начало МИС 2, LGM). Дат по костям малого пещерного медведя получено значительно меньше. Все они соответствуют МИС 3 (Косинцев и др., 2003; Gimranov, Kosintsev, 2020), и только одна дата из Кизеловской пещеры (18 800 ± ± 340, ИГАН-340) попадает в МИС 2. По образцам из этой пещеры получено еще 11 дат, все из которых древнее 30 тыс. лет (Pacher, Stuart, 2009; Barlow et al., 2021). Вероятно, наиболее поздняя дата омоложена. Косвенно это подтверждает тот факт, что ни в одном из многочисленных местонахождений крупной териофауны на Урале, датируемых периодом МИС 2, не найдено костей малого пещерного медведя. Таким образом, анализ хронологии находок большого и малого пещерных медведей показывает, что в конце среднего плейстоцена (конец МИС 6?) и на протяжении большей части позднего плейстоцена (МИС 5–МИС 3) большой и малый пещерные медведи сосуществовали на территории Урала.
ЧЕЛОВЕК И ПЕЩЕРНЫЕ МЕДВЕДИ НА УРАЛЕ
Высказывавшееся ранее предположение о существовании систематической охоты на пещерных медведей в палеолите Урала (Кузьмина, 1971; Верещагин, 1973) не подтвердилось. На палеолитических стоянках Урала не найдено костей большого пещерного медведя (Kosintsev, Plasteeva, 2015), за исключением переотложенных (Гуслицер, Канивец, 1965). На более чем 50 тыс. просмотренных костных остатках большого пещерного медведя из 52 местонахождений не найдено следов от орудий и искусственных повреждений. Из 14 тыс. костей малого пещерного медведя только на пяти костях (мустьерская стоянка Богдановка на Южном Урале; Широков др., 2011) имеются следы от орудий. Также в пещере Иманай на Южном Урале найден череп малого пещерного медведя с отверстием, пробитым каменным орудием (Котов и др., 2020; Гимранов и др., 2021а) Таким образом, на Урале имеются свидетельства лишь единичных случаев охоты только на малого пещерного медведя.
ПРИЧИНЫ ВЫМИРАНИЯ ПЕЩЕРНЫХ МЕДВЕДЕЙ НА УРАЛЕ
Предложено несколько причин вымирания пещерных медведей: в частности, из-за изменения режима весенних паводков, в результате чего во время ранних оттепелей вода могла заливать пещеры и затапливать спящих животных, а также в результате дегенеративного вырождения изолированных популяций (Верещагин, 1971, 1973, 1982). Вымирание могло произойти в результате действия нескольких факторов: морфофункциональной сверхспециализации (пасторальная специализация и низкий уровень обмена веществ); исчезновение ландшафта холодных степей в начале голоцена и смена погодных условий; истребление человеком и конкуренция с ним за “удобные” пещеры (Верещагин, 1973).
Причиной вымирания также мог быть биоценотический фактор. Пещерные медведи были зональными элементами мамонтовой биоты и вымерли в результате ее распада в конце плейстоцена. Указан механизм вымирания – недостаточное воспроизводство популяций в результате повышенной смертности неполовозрелых особей во время зимней спячки. При этом объяснении исключаются как антропогенный фактор, так и фактор дегенеративного вырождения (Косинцев, Воробьев, 2001).
В последнее время для выяснения причин вымирания представителей “мамонтовой” фауны используют результаты анализа древней ДНК сопряженно с данными по динамике климата, деятельности человека и другим параметрам (Lorenzen et al., 2011; Murray et al., 2017; Knapp, 2019). В то же время, для палеопопуляций медведей Урала такие данные пока отсутствуют.
ЗАКЛЮЧЕНИЕ
Научное изучение пещерных медведей Урала продолжается более 80 лет. За это время собрано и изучено более 50 тыс. костей большого пещерного медведя (U. kanivetz) из 52 местонахождений и около 14 тыс. костей малого пещерного медведя (U. ex gr. savini–rossicus) из 17 местонахождений. Установлено их совместное обитание на западном склоне Урала на протяжении большей части позднего плейстоцена (МИС 5–МИС 3). Показано, что большой пещерный медведь не населял восточный склон Урала. В результате анализа ядерной ДНК установлена таксономическая самостоятельность и определено время дивергенции филетических линий малого и большого пещерных медведей Урала. Получены морфометрические характеристики костей и зубов обоих видов. Изучены возрастная и половая изменчивость размеров и пропорций ряда костей и зубов. Получены оценки возрастного и полового состава погибших особей в пещерах Кизеловской, Тайн и Геологов 3 на Среднем Урале, а также в пещерах Игнатиевской, Аше 1 и Иманай на Южном Урале. Фактов добычи человеком большого пещерного медведя не обнаружено. Выявлены единичные факты добычи малого пещерного медведя. Вымирание на Урале малого пещерного медведя произошло около 26 тыс., а большого пещерного медведя – около 20 тыс. л. н.
Следует отметить различную степень изученности пещерных медведей Урала. Значительно лучше изучен большой пещерный медведь, о малом пещерном медведе данных значительно меньше. Остаются не полностью изученными крупные коллекции остатков большого и малого пещерных медведей из пещер: Махневская ледяная, Тайн и Геологов 3 на Среднем Урале, Барсучий Дол, Заповедная и Победа на Южном Урале. Имеется недостаточное число радиоуглеродных дат и данных по стабильным изотопам (C13, N15, O18, 87Sr/86Sr). Генетическое разнообразие пещерных медведей Урала также остается почти неизученным.
Авторы благодарны рецензентам за замечания и рекомендации, которые позволили улучшить качество публикации и обсудить ряд дискуссионных вопросов. Благодарим музей Института экологии растений и животных УрО РАН за предоставленную информацию. Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 20-14-50162 и финансовой поддержке РНФ, проект № 20-74-00041.
Список литературы
Абрамов А.В., Барышников Г.Ф. Каталог типовых экземпляров коллекции Зоологического института АН СССР. Млекопитающие (Mammalia). Вып. 2. Хищные (Carnivora), хоботные (Proboscidea), десмостилии (Desmostilia). Л.: Наука, 1990. 23 с.
Барышников Г.Ф. Отряд Хищные – Carnivora, Bowdich, 1821 // Каталог млекопитающих СССР / Ред. И.М. Громов, Г.И. Баранова. Л.: Наука, 1981. С. 236–292.
Барышников Г.Ф. Систематика и филогения семейства медвежьих (Carnivora, Ursidae). Автореф. дис. … докт. биол. наук. СПб., 2003. 52 с.
Барышников Г.Ф. Семейство медвежьих (Carnivora, Ursidae). СПб.: Наука, 2007. 542 с.
Бачура О.П., Косинцев П.А. Ареалы млекопитающих на Урале в конце позднего плейстоцена // Зоол. журн. 2019. Т. 98. С. 933–948.
Борисяк A.A. Ursus spelaeus rossicus nov. n. // Докл. АН СССР. 1930. Т. 8. С. 102–104.
Верещагин Н.К. “Следы” древней жизни на Северном Урале // Природа. 1957. № 4. С. 103–104.
Верещагин Н.К. Млекопитающие Кавказа. М.-Л.: Изд-во АН СССР, 1959. 704 с.
Верещагин Н.К. Охоты первобытного человека и вымирание плейстоценовых млекопитающих в СССР // Тр. Зоол. ин-та АН СССР. 1971. Т. 69. С. 200–232.
Верещагин Н.К. Краниологическая характеристика современных и ископаемых медведей // Зоол. журн. 1973. Т. 52. С. 920–930.
Верещагин Н.К. Кизеловская пещера – ловушка зверей на Среднем Урале // Тр. Зоол. ин-та АН СССР. 1982. Т. 111. С. 37–44.
Верещагин Н.К., Кузьмина И.Е. Раскопки в пещерах Северного Урала // Природа. 1962. № 3. С. 76–78.
Гимранов Д.О., Косинцев П.А., Бачура О.П. и др. Малый пещерный медведь (U. ex gr. savini-rossicus) как объект охоты древнего человека // Вестн. археол. антропол. этногр. 2021а. Т. 21. № 2. С. 5–14.
Гимранов Д.О., Павлова М.В., Нанова О.Г. и др. Новые данные о распространении малого пещерного медведя (Ursus ex gr. savini-rossicus) на Урале // Докл. РАН. Науки о жизни. 2021б. Т. 499. С. 291–295.
Гофман Э. Северный Урал и береговой хребет Пай-Хой. СПб.: Тип. Имп. Акад. наук, 1856. 376 с.
Гуслицер Б.И., Канивец В.И. Пещеры Печорского Урала. М.: Наука, 1965. 134 с.
Гуслицер Б.И., Павлов П.Ю. О первоначальном заселении северо-востока Европы. Сыктывкар: Коми фил-л АН СССР, 1987. 24 с.
Громов В.И. Палеонтологическое и археологическое обоснование стратиграфии континентальных отложений четвертичного периода на территории СССР (млекопитающие, палеолит). М.: Изд-во АН СССР, 1948. 521 с. (Тр. Геол. ин-та АН СССР. Вып. 64).
Карачаровский В.В. Позднечетвертичная фауна бассейна р. Юрюзань (Южный Урал) // Матер. иссл. археол. СССР. 1951. Т. 2. № 21. С. 244–269.
Косинцев П.А. О структуре популяции пещерного медведя Южного Урала // Экология популяций. Новосибирск, 1988. С. 120–121.
Косинцев П.А. Крупные млекопитающие Урала в плейстоцене и голоцене // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Урал. ун-та, 2003. С. 55–72.
Косинцев П. А. Палеонтологические находки в пещерах // Атлас пещер России. М.: Русс. геогр. об-во, 2019. С. 54–59.
Косинцев П.А., Бородин А.В. Териофауна восточного склона Северного Урала в позднем плейстоцене и голоцене // Тр. Зоол. ин-та АН СССР. 1990. Т. 212. С. 120–134.
Косинцев П.А., Воробьев А.А. Биология большого пещерного медведя (Ursus spelaeus Ros. et Hein.) на Урале // Мамонт и его окружение: 200 лет изучения. М.: ГЕОС, 2001. С. 266–278.
Косинцев П.А., Воробьев А.А., Орлова Л.А. Абсолютные даты по ископаемым медведям (род Ursus) Среднего Урала // Териофауна России и сопредельных территорий. Матер. Междунар. совещ. М.: РФФИ, 2003. С. 174–175.
Косинцев П.А., Гимранов Д.О., Лавров И.А. и др. Новое массовое захоронение пещерных медведей (Carnivora, Ursidae, Ursus kanivetz Vereshchagin, 1973) на Среднем Урале // Докл. РАН. Науки о жизни. 2021. Т. 498. С. 245–248.
Косинцев П.А., Сатаев Р.М. Фауна млекопитающих из местонахождения Аша I (Юж. Урал) // Фауна Урала и Сибири в плейстоцене и голоцене. Челябинск: Рифей, 2005. С. 113–147.
Котов В.Г., Румянцев М.М., Гимранов Д. О. Стоянка среднего палеолита в пещере Иманай–1 на Южном Урале: предварительные итоги археологических исследований // Orient. Stud. 2020. Т. 13. № 5. С. 1271–1291.
Кузьмина И.Е. Формирование териофауны Северного Урала в позднем антропогене // Тр. Зоол. ин-та АН СССР. 1971. Т. 49. С. 44–122.
Кузьмина И.Е. Некоторые данные о млекопитающих Среднего Урала в позднем плейстоцене // Бюлл. комисс. изуч. четвертич. пер. АН СССР. 1975. № 43. С. 63–77.
Кузьмина И.Е. Пещерные медведи Урала // Фауна Урала в плейстоцене и голоцене: сб. науч. тр. Екатеринбург: Изд-во “Университет”, 2002. С. 136–145.
Кузьмина И.Е., Абрамсон Н.И. Остатки млекопитающих в Каповой пещере на Южном Урале // Пещерный палеолит Урала. Междунар. науч. конф. Уфа: Изд-во Ин-та истории, языка и лит-ры УфНЦ РАН, 1997. С. 124–127.
Кузьмина С.А. Фаунистические данные по позднепалеолитической стоянке Смеловская II на Южном Урале // Плейстоценовые и голоценовые фауны Урала. Челябинск: Рифей, 2000. С. 137–153.
Маркова А.К., ван Кольфсхотен Т., Бохнкке Ш. и др. Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н.). М.: КМК, 2008. 549 с.
Павлова М.В., Гимранов Д.О., Нанова О.Г. и др. Распространение малого пещерного медведя (Ursus ex gr. savini-rossicus) на Урале // Теоретические и прикладные аспекты палеонтологии. Матер. LXVI сесс. Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2020. С. 121–122.
Петрин В.Т., Смирнов Н.Г. Палеолитические памятники в гротах Среднего Урала и некоторые вопросы палеолитоведения Урала // Вопр. археол. Урала. 1977. Вып. 14. С. 56–71.
Прилепская Н.Е., Барышников Г.Ф. Определение возраста и сезона гибели пещерных медведей по цементу и дентину зубов // Морфологическая эволюция и стратиграфические проблемы. Матер. LXV сесс. Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2019. С. 262–263.
Ражев Д.И., Косинцев П.А., Улитко А.И. Фауна крупных млекопитающих позднего плейстоцена и голоцена из грота Бобылек (Средний Урал) // Фауна Урала и Сибири в плейстоцене и голоцене. Челябинск: Рифей, 2005. С. 190–211.
Сатаев Р.М. Сравнительная характеристика ископаемых фаун из пещер “Победа” и “Заповедная” // Ежегодник – 1994. Информ. материалы. Уфа: УНЦ РАН, 1995. С. 40–42.
Сатаев Р.М. Остатки медведя (Spelaearctos) sp. и носорога Stephanorhinus sp. из пещеры Верхняя (Южный Урал) // Фауна и флора Северной Евразии в позднем кайнозое. Екатеринбург–Челябинск: Рифей, 2008. С. 137–146.
Сивкова Т.Н., Косинцев П.А. Первая находка яиц нематоды Baylisascaris transfuga Rudolphi, 1819 (Ascaridoidea, Nematoda) в позднем плейстоцене // Докл. РАН. Науки о жизни. 2021. Т. 499. С. 26–28.
Силаев В.И., Паршукова М.Н., Гимранов Д.О. и др. Минералого-геохимические особенности пещерной фоссилизации костного детрита на примере плейстоценовой пещеры Иманай (Южный Урал) // Вестн. Перм. ун-та. Геол. 2020. № 4. С. 323–358.
Смирнов Н.Г., Большаков В. Н., Косинцев П.А. и др. Историческая экология животных гор Южного Урала. Свердловск: УрО АН СССР, 1990. 245 с.
Улитко А.И. Локальные фауны крупных млекопитающих из отложений пещеры Дыроватый Камень на реке Серга (Средний Урал) // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского ун-та, 2003. С. 185–192.
Фадеева Т.В., Косинцев П.А., Гимранов Д.О. Млекопитающие горной части Южного Урала в последнее межледниковье // Зоол. журн. 2019. Т. 98. С. 1304–1322.
Фадеева Т.В., Косинцев П.А., Кадебская О.И. и др. Результаты исследований зоогенных отложений пещеры Махневская ледяная (Пермский край) // Пещеры. 2011. Вып. 34. С. 71–99.
Широков В.Н., Волков Р.Б., Косинцев П.А. и др. Палеолитическая стоянка Богдановка (Южный Урал) // Росc. археол. 2011. № 1. С. 111–125.
Юрин В.И. Животные и пещеры Южного Урала // Из опыта исследований спелеообъектов Южного Урала и Зауралья. Челябинск: Цицеро, 2011. С. 126–144.
Юрин В.И. Биостратиграфические исследования отложений верхнего плейстоцена и голоцена пещеры Сикияз-Тамак 7 (Южный Урал, Россия) // Пещеры челябинской области: история открытий и результаты исследований 2011–2020. Челябинск: Край Ра, 2020. С. 225–243.
Яковлев А.Г., Данукалова Г.А., Алимбекова Л.И. и др. Биостратиграфическая характеристика геологического памятника природы “Пещера Нукатская” // Плейстоценовые и голоценовые фауны Урала. Челябинск: Рифей, 2000. С. 81–104.
Яковлев А.Г., Данукалова Г.А., Алимбекова Л.И. и др. Биостратиграфическая характеристика отложений позднего неоплейстоцена–голоцена в районе памятника природы “водопад Атыш” (Южный Урал) // Фауна Урала и Сибири в плейстоцене и голоцене. Челябинск: Рифей, 2005. С. 260–304.
Baca M., Popović D., Stefaniak K. et al. Retreat and extinction of the Late Pleistocene cave bear (Ursus spelaeus sensu lato) // Sci. Natur. 2016. V. 103. P. 1–17.
Bachura O., Kosintsev P. Late Pleistocene and Holocene small- and large-mammal faunas from the Northern Urals // Quatern. Intern. 2007. V. 160. P. 121–128.
Barlow A., Paijmans J., Alberti F. et al. Middle Pleistocene genome calibrates a revised evolutionary history of extinct cave bears // Curr. Biol. 2021. V. 31. P. 1771–1779.
Baryshnikov G. Morphometrical variability of cheek teeth of cave bears // Sci. Ann., School Geol. Aristotle Univ. Thessaloniki. 2006. V. 98. P. 81–102.
Baryshnikov G.F., Foronova I.V. Pleistocene small cave bear (Ursus rossicus) from the South Siberia, Russia // Cadernos Lab. Xeolóxico Laxe Coruña. 2001. V. 26. P. 373–398.
Baryshnikov G., Gimranov D., Kosintsev P. Variability of the upper incisors in the cave bears (Carnivora, Ursidae) from the Caucasus and Urals // C. R. Palevol. 2019. V. 18. P. 209–222.
Baryshnikov G., Puzachenko A. Craniometrical variability in the cave bears (Carnivora, Ursidae): multivariate comparative analysis // Quatern. Intern. 2011. V. 245. P. 350–368.
Baryshnikov G., Puzachenko A. Morphometric analysis of metacarpal and metatarsal bones of cave bears (Carnivora, Ursidae) // Foss. Impr. 2017. V. 73. P. 7–47.
Baryshnikov G.F., Puzachenko A.Y. Morphometry of upper cheek teeth of cave bears (Carnivora, Ursidae) // Boreas. 2019. V. 48. P. 581–604.
Baryshnikov G.F., Puzachenko A.Y. Morphometry of lower cheek teeth of cave bears (Carnivora, Ursidae) and general remarks on the dentition variability // Boreas. 2020. V. 49. P. 562–593.
Baryshnikov G.F., Puzachenko A.Y., Baryshnikova S.V. Morphometric analyses of cave bear mandibles (Carnivora, Ursidae) // Rev. Paléobiol. 2018. V. 37. P. 379–393.
Danukalova G., Kosintsev P., Yakovlev A. et al. Quaternary deposits and biostratigraphy in caves and grottoes located in the Southern Urals (Russia) // Quatern. Intern. 2020. V. 546. P. 84–124.
Danukalova G., Yakovlev A., Alimbekova L. et al. Biostratigraphy of the Upper Pleistocene (Upper Neopleistocene) – Holocene deposits of the Lemeza River valley of the Southern Urals region (Russia) // Quatern. Intern. 2008. V. 190. P. 38–57.
Danukalova G., Yakovlev A., Kosintcev P. et al. Quaternary fauna and flora of the Southern Urals region: (Bashkortostan Republic) // Quatern. Intern. 2009. V. 201. P. 13–24.
Fadeeva T., Kosintsev P., Lapteva E. et al. Makhnevskaya Ledyanaya Cave (Middle Urals, Russia): Biostratigraphical reconstruction // Quatern. Intern. 2020. V. 546. P. 135–151.
Gimranov D.O., Kosintsev P.A. Quaternary large mammals from the Imanay Cave // Quatern. Intern. 2020. V. 546. P. 125–134.
Gretzinger J., Molak M., Reiter E. et al. Large-scale mitogenomic analysis of the phylogeography of the Late Pleistocene cave bear // Sci. Rep. 2019. V. 9: 10700.
Knapp M. From a molecules’ perspective – contributions of ancient DNA research to understanding cave bear biology // Histor. Biol. 2019. V. 31. P. 442–447.
Knapp M., Rohland N., Weinstock J. et al. First DNA sequences of Asian cave bear fossils reveal deep divergences and complex phylogeographic patterns // Mol. Ecol. 2009. V. 18. P. 1225–1238.
Kosintsev P.A. Late Pleistocene large mammal faunas from the Urals // Quatern. Intern. 2007. V. 160. P. 112–120.
Kosintsev P.A., Bachura O.P. Late Pleistocene and Holocene mammal fauna of the Southern Urals // Quatern. Intern. 2013. V. 284. P. 161–170.
Kosintsev P.A., Gasilin V.V., Gimranov D.O. et al. Carnivores of the Ural in the late Pleistocene and Holocene // Quatern. Intern. 2016. V. 420. P. 145–155.
Kosintsev P.A., Plasteeva N.A. Paleolithic hunting in the Urals (Russia) // Forgotten times and spaces: New perspectives in paleoanthropological, paleoetnological and archeological studies / Ed. S. Sázelová, M. Novák, A. Mizerová. Brno: Inst. Archeol. Czech Acad. Sci., Masaryk Univ., 2015. P. 91–97.
Kurtén B. The cave bear story: life and death of a vanished animal. N.Y.: Columbia Univ. Press, 1995. 163 p.
Lorenzen E.D., Nogues-Bravo D., Orlando L. et al. Species-specific responses of Late Quaternary megafauna to climate and humans // Nature. 2011. V. 479. P. 359–364.
Murray G.G.R., Soares A.E.R., Novak B.J. et al. Natural selection shaped the rise and fall of passenger pigeon genomic diversity // Science. 2017. V. 954. P. 1–14.
Naito Y., Meleg I., Robu M. et al. Heavy reliance on plants for Romanian cave bears evidenced by amino acid nitrogen isotope analysis // Sci. Rep. 2020. V. 10: 6612. https://doi.org/10.1038/s41598-020-62990-0
Pacher M., Stuart A.J. Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus) // Boreas. 2009. V. 38. P. 189–206.
Puzachenko A.Y., Titov V.V., Kosintsev P.A. Evolution of the European regional large mammals assemblages in the end of the Middle Pleistocene – the first half of the Late Pleistocene (MIS 6–MIS 4) // Quatern. Intern. 2020. https://doi.org/10.1016/j.quaint.2020.08.038
Rabeder G., Hofreiter M., Nagel D. et al. New taxa of Alpine Cave Bears (Ursidae, Carnivora) // Actes 9e Symp. Intern. sur l’ours des cavernes, Entremont-le-Vieux (Savoie, France). V. 2. Lyon: Museum Lyon, 2004. P. 49–67.
Rabeder G., Hofreiter M., Stiller M. Chronological and systematic position of cave bear fauna from Ajdovska jama near Krško (Slovenia) // Mitt. Komm. Quartärforsch. Österr. Akad. Wiss. 2011. V. 20. P. 79–85.
Sataev R.M. Question on a geological age Spelaearctos cf. rossicus out of the cave “Verchnaya” in South Ural // INQUA–SEQS Symposium. Abstracts. Netherlands, Kerkrade, 1996. P. 86.
Sher A.V., Weinstock J., Baryshnikov G.F. et al. The first record of “spelaeoid” bears in Arctic Siberia // Quatern. Sci. Rev. 2011. V. 30. P. 2238–2249.
Spassov N., Hristova L., Ivanova S. et al. First record of the “small cave bear” in Bulgaria and the taxonomic status of bears of the Ursus savini Andrews – Ursus rossicus Borissiak group // Foss. Impr. 2017. V. 73. P. 275–291.
Stiller M., Baryshnikov G., Bocherens H. et al. Withering away–25.000 years of genetic decline preceded cave bear extinction // Mol. Biol. Evol. 2010. V. 27. P. 975–978.
Stiller M., Knapp M., Stenzel U. et al. Direct multiplex sequencing (DMPS) a novel method for targeted high–throughput sequencing of ancient and highly degraded DNA // Gen. Res. 2009. V. 19. P. 1843–1848.
Stiller M., Molak M., Prost S. et al. Mitochondrial DNA diversity and evolution of the Pleistocene cave bear complex // Quatern. Intern. 2014. V. 339–340. P. 224–231.
Vereschagin N., Baryshnikov G. Small cave bear Ursus (Spelearctos) rossicus uralensis from the Kizel Cave in the Ural (Russia) // Geoloski zborn. 2000. № 15. P. 53–66.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал