

Палеонтологический журнал, 2022, № 2, стр. 79-88
Обзор юрских долгоносиков рода Belonotaris Arnoldi (Coleoptera: Nemonychidae) с прямой головотрубкой
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Алтайский государственный университет
656049 Барнаул, Россия
c Томский государственный университет
634050 Томск, Россия
* E-mail: fossilweevils@gmail.com
Поступила в редакцию 24.02.2021
После доработки 08.04.2021
Принята к публикации 27.04.2021
- EDN: ICAILN
- DOI: 10.31857/S0031031X22020076
Аннотация
Ревизованы виды с прямой головотрубкой, относящиеся к роду Belonotaris Arnoldi, 1977 из средней–верхней юры Казахстана и Китая. Arnoldibelus Legalov, 2009 syn. nov. и Sinoeuglypheus Yu, Davis et Shih, 2019 syn. nov. сведены в синонимы к роду Belonotaris; его диагноз дополнен и установлена его семейственная принадлежность. Составлена определительная таблица для B. daohugouensis (Yu, Davis et Shih, 2019) comb. nov., B. gratshevi (Legalov, 2009) и B. rohdendorfi Legalov, 2014. Обсуждается сходство юрских фаун долгоносикообразных жуков из Даохугоу, Каратау, Шар-Тэга и Талбрагара. Юрские Nemonychidae, вероятно, развивались как на Bennettitales, так и на голосеменных из других групп (Cycadales, Ginkgoales, Czekanowskiales и Coniferales). Виды рода Belonotaris могли быть связаны с Pterophyllum и другими беннеттитовыми.
ВВЕДЕНИЕ
Долгоносикообразные жуки (Curculionoidea) – очень разнообразная группа кукуйиформных жесткокрылых. Наиболее примитивными среди них являются семейства Nemonychidae, Anthribidae, Ithyceridae и Belidae (Legalov, 2012, 2018; Gratshev, Legalov, 2014). Самые ранние Curculionoidea найдены в средней–верхней юре (келловей–оксфорд), причем они представлены специализированными формами. До последнего времени наиболее древними считались формы из средней–верхней юры Каратау, откуда описана богатая фауна Nemonychidae (Арнольди, 1977; Грачев, Жерихин, 1995; Gratshev, Zherikhin, 1996; Легалов, 2010; Legalov, 2012, 2015; Gratshev, Legalov, 2014). Кроме того, там отмечены отдельные Anthribidae (Легалов, 2011б, 2013; Legalov, 2012, 2015) и Ithyceridae (Грачев, Легалов, 2011; Legalov, 2012, 2015). Недавно из средней–верхней юры Даохугоу был описан представитель Curculionoidea, помещенный в семейство Belidae (Yu et al., 2019). Однако в ходе изучения этого вида не было выполнено сравнение с юрскими долгоносикообразными Каратау, что привело к ошибке при определении его систематического положения.
В работе представлены уточненные реконструкции Sinoeuglypheus daohugouensis Yu, Davis et Shih, 2019 и близких к нему видов из Казахстана, что позволило уточнить структуру рода Belonotaris Arnoldi, 1977 и систематическое положение S. daohugouensis.
МАТЕРИАЛ И МЕТОДЫ
Юрские долгоносикообразные жуки обнаружены в четырех местонахождениях. Даохугоу (формация Хайфангоу) расположено в окр. дер. Даохугоу (Внутренняя Монголия, Китай), возраст принимается как келловей–оксфордский (Jiang et al., 2010; Pott, Jiang, 2017); Каратау (карабастауская свита) – обнажения на берегу р. Кашкар-Ата, отроги хр. Каратау, близ дер. Маятас (быв. Михайловка) (Толебийский р-н, Туркестанская обл., Казахстан), верхний келловей–оксфорд (Полянский, Долуденко, 1978); Шар-Тэг (шартэгская толща) – южнее г. Шap-Tэг, ю.‑в. xp. Aдж-Бoгдo (Гоби-Алтай, Монголия), верхний келловей–оксфорд (Ponomarenko et al., 2014); Талбрагар (формация Purlawaugh) – 30 км с.-в. г. Галгонг (Новый Южный Уэльс, Австралия), оксфорд–титон (Bean, 2006).
Материал по роду Belonotaris хранится в коллекциях Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН и Колледжа естественных наук Столичного педагогического ун-та в Пекине (CNU).
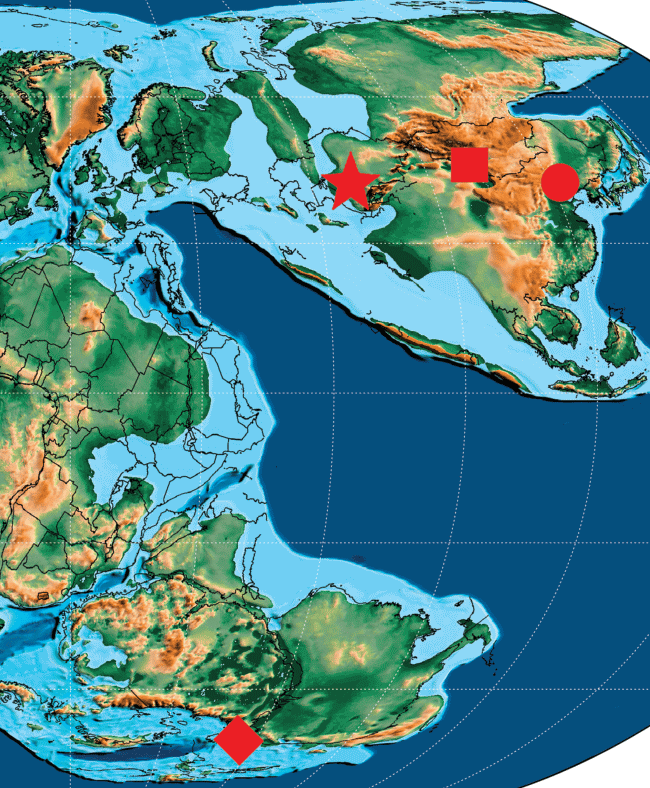
Автор выражает благодарность А.Г. Пономаренко и Е.В Яну за предоставление сравнительного материала из коллекции ПИН РАН (Москва), а также К.Р. Скотезе (Эванстон) за разрешение использовать его палеогеографическую реконстукцию поздней юры (рис. 2).
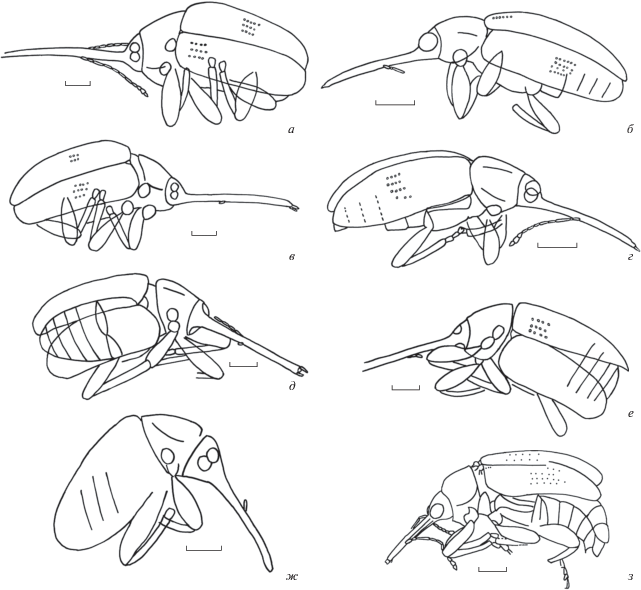
Рис. 1.
Представители рода Belonotaris: а, в – B. gratshevi, голотип ПИН, № 2904/958: а – обратный отпечаток, дорсо-латерально; в – прямой отпечаток, дорсо-латерально; б, г–ж – B. rohdendorfi: б, г – голотип ПИН, № 2784/1452: б – прямой отпечаток, вид слева, латерально; г – обратный отпечаток, вид справа, латерально; д, е – экз. ПИН, № 2784/1450: д – прямой отпечаток, дорсально; е – обратный отпечаток, дорсально; ж – экз. ПИН, № 2384/535, обратный отпечаток, вид справа, дорсолатерально; з – B. daohugouensis, голотип CNU-COL-NN2011122, прямой отпечаток, вид справа, латерально. Длина масштабного отрезка 1 мм.
ТАКСОНОМИЧЕСКАЯ ЧАСТЬ
НАДСЕМЕЙСТВО CURCULIONOIDEA LATREILLE, 1802
Описанный из юры Китая Sinoeuglypheus daohugouensis Yu et al., 2019 был помещен в семейство Belidae без указания подсемейства на основании наличия выемки на вершинном внутреннем крае передних голеней (Yu et al., 2019). Современные Belidae представлены двумя специализированными подсемействами: Belinae, имеющими удлиненную форму тела, развивающимися в стеблях и ветвях (Marvaldi, Ferrer, 2014), и уплощенными Oxycoryninae, обитающими в генеративных органах растений, реже под корой (Marvaldi et al., 2006; Marvaldi, Ferrer, 2014). Общей характеристикой этих габитуально несходных групп в первую очередь служат передние голени с широкой апикальной выемкой и густыми волосками на внутренней поверхности в месте прикрепления лапок (Marvaldi et al., 2006; Legalov, 2009, 2015). Признаки современных Belidae: верхняя губа, сросшаяся с головотрубкой; бока переднеспинки без килей; гулярных шва два, но они сросшиеся или почти сросшиеся на вершине [как исключение, гулярный шов может быть одинарный (Allocorynitae)]; усиковые бороздки не развитые; усики не коленчатые; вентриты брюшка гомономные, обычно свободные, тело сильно склеротизованное. Представители подсемейства Belinae также отличаются гранулированными переднеспинкой и надкрыльями. Однако у раннемелового подсемейства Montsecbelinae передние голени простые, хотя, как и у Belinae, киль на боках переднеспинки отсутствовал, тело было сильно склеротизованное, верхняя губа сросшаяся, а переднеспинка и надкрылья гранулированные (Legalov, 2015).
Семейство Nemonychidae является самым примитивным среди долгоносикообразных жуков. К его характеристикам относятся: свободная верхняя губа (лишь у подсемейства Idiomacerinae она сросшаяся с головотрубкой); двойные гулярные швы, широко разделенные на вершине; не коленчатые усики; довольно длинные максиллярные пальпы; гомономные стерниты брюшка; передние голени всегда без апикальной выемки на внутренней вершинной трети, и слабо склеротизованное тело. Таким образом, друг от друга семейства Nemonychidae и Belidae можно отличить по свободной или сросшейся с головотрубкой верхней губе, длине максиллярных пальп, степени склеротизованности тела и наличию гранул на переднеспинке и надкрыльях, по гулярным швам, широко разделенным или сближенным на вершине (иногда у Belidae гулярный шов одинарный или же швы редуцированы), а также по наличию или отсутствию широкой апикальной выемки передних голеней. У S. daohugouensis максиллярные пальпы, как и мандибулы, не сохранились, и строение гулярных швов не видно, но верхняя губа свободная, на что указывает четкий шов между ней и головотрубкой. Кроме того, бока переднеспинки этого вида имеют явственный латеральный киль; его тело слабо склеротизовано, переднеспинка и надкрылья не уплощенные и не гранулированные, а внимательное изучение передних голеней показало, что выемка на голенях отсутствует.
Семейства Anthribidae и Ithyceridae также были обнаружены в юре (Грачев, Легалов, 2011; Легалов, 2011б, 2013). Однако юрские представители первого характеризуются короткой широкой и уплощенной головотрубкой и часто очень длинными усиками, достигающими надкрылий или превышающими размер тела. Усики S. daohugouensis даже не достигают надкрылий, а головотрубка почти цилиндрическая, длинная. Ithyceridae в юре представлены подсемействами Mongolocarinae и Baissorhynchinae, которым свойственно сильно склеротизованное тело, короткая прекоксальная часть переднегруди и обычно суббазально прикрепленные усики. S. daohugouensis отличается от них слабо склеротизованным телом, удлиненной прекоксальной частью переднегруди и усиками, прикрепленными перед серединой переднеспинки. Все перечисленные признаки показывают, что S. daohugouensis является представителем семейства Nemonychidae.
Подсемейства Nemonychidae делятся на две группы: примитивные с килем на боках переднеспинки и более продвинутые, встречающиеся и в современной фауне – без такого киля. S. daohugouensis относится к первой группе. Передние тазики, расположенные возле основания переднегруди, и усики, прикрепленные около середины головотрубки, указывают на принадлежность S. daohugouensis к доминирующему в юре подсемейству Eobelinae, с представителями которого при первоописании сравнение поведено не было (Yu et al., 2019). Это подсемейство насчитывает четыре трибы: Eobelini, Probelini, Oxycorynoidini и Karataucarini. Длинная головотрубка и не укрупненные первые членики лапок позволяют причислить S. daohugouensis к трибе Probelini. В ее состав входят только два рода: Belonotaris и Probelus, отличающиеся по наличию или отсутствию явственного мукро на вершинах голеней. S. daohugouensis не имеет мукро; кроме того, он характеризуется длинной головотрубкой (длиннее головы и переднеспинки вместе взятых) и крупными размерами тела. Поэтому название Sinoeuglypheus Yu, Davis et Shih, 2019 здесь синонимизируется с Belonotaris.
СЕМЕЙСТВО NEMONYCHIDAE BEDEL, 1882
ПОДСЕМЕЙСТВО EOBELINAE ARNOLDI, 1977
Триба Probelini Legalov, 2009
Род Belonotaris Arnoldi, 1977
Belonotaris: Арнольди, 1977, с. 154.
Arnoldibelus: Legalov, 2009, с. 287.
Sinoeuglypheus: Yu et al., 2019, с. 2106.
Типовой вид – Belonotaris punctatissimus Arnoldi, 1977 из средней–поздней юры Каратау.
Диагноз. Крупные (4.2–10.0 мм), слабо уплощенные, довольно слабо хитинизированные, темно-коричневые жуки. Головотрубка приближается к нижней, слабо равномерно изогнутая или почти прямая, иногда сужающаяся к вершине. Верхняя губа явственная, свободная. Усики довольно короткие, не коленчатые, прикреплены возле середины головотрубки. Членики жгутика усиков удлиненные. Третий членик усиков длинный. Булава явственная, не компактная. Глаза среднего размера или крупные, круглые, выпуклые, их диаметр иногда значительно больше диаметра головотрубки. Лоб сильновыпуклый. Переднеспинка уплощенная или выпуклая, с явственным боковым килем. Диск мелко и редко пунктированный. Надкрылья более или менее уплощенные, с прищитковой бороздкой и со слабыми точечными бороздками. Наружный край надкрылий слабо вырезан возле задних тазиков. Прекоксальная часть переднегруди более или менее удлиненная. Стерниты брюшка примерно равной длины. Передние тазики смещены к заднему краю переднегруди. Бедра всех ног утолщенные. Голени почти прямые или изогнутые в вершинной части, без явственного мукро. Лапки не расширенные. Первый членик лапок не укрупненный, ýже голеней.
Видовой состав. К роду относятся шесть видов из средней–поздней юры Каратау и один вид из средней–поздней юры Китая. Виды подразделяются на две группы: с прямой головотрубкой [B. daohugouensis (Yu, Davis et Shih, 2019), B. gratshevi (Legalov, 2009) и B. rohdendorfi Legalov, 2014] и с явственно изогнутой головотрубкой (B. heeri Legalov, 2014, B. karatavicus Arnoldi, 1977, B. punctatissimus Arnoldi, 1977 и B. rasnitsyni Legalov, 2014).
Замечания. Род Belonotaris подразделялся на два подрода, отличавшихся по длине головотрубки и размерам тела: номинативный подрод и Arnoldibelus Legalov, 2009 (Gratshev, Legalov, 2014). Переизучение видов из Каратау, входящих в этот род, а также китайского вида показало, что четкой границы между Belonotaris и Arnoldibelus syn. nov. провести нельзя.
Определительная таблица видов рода Belonotaris с прямой головотрубкой
1. Головотрубка очень длинная (рис. 1, а, в), примерно в 17 раз больше ширины на середине и в 2.0 раза длиннее головы и переднеспинки вместе взятых B. gratshevi
– Головотрубка длинная (рис. 1, б, г–з), примерно в 9–15 раз больше ширины на середине и практически равна длине голове и переднеспинки, вместе взятых, или же в 1.2–1.5 раза длиннее их.
2. Головотрубка примерно в 9 раз длиннее ширины на середине, едва длиннее головы и переднеспинки, вместе взятых (рис. 1, з) B. daohugouensis
– Головотрубка примерно в 13–15 раз длиннее ширины на середине, в 1.2–1.5 раз длиннее головы и переднеспинки, вместе взятых (рис. 1, б, г–ж)............................B. rohdendorfi
Belonotaris daohugouensis (Yu, Davis et Shih, 2019), comb. nov.
Sinoeuglypheus daohugouensis: Yu et al., 2019, c. 2109, рис. 1–10 .
Голотип – CNU-COL-NN2011122, прямой отпечаток; Даохугоу; средняя юра, формация Хайфангоу.
Описание (рис. 1, з). Длина головотрубки в 12.3 раза больше ее ширины на вершине, в 9.3 раза больше ширины на середине, в 6.2 раза больше ширины у основания, в 1.7 раза больше длины переднеспинки, едва длиннее головы и переднеспинки, вместе взятых. Головотрубка прямая, слабо сужающаяся к вершине. Верхняя губа явственная, немного шире своей длины, на вершине со слабой выемкой на середине. Глаза крупные, их длина немного больше их ширины. Длина виска составляет 0.7 длины глаза. Усики прикреплены немного за серединой головотрубки, заходят за передний край переднеспинки. Членики жгутика удлиненно-трапециевидные. Длина первого членика усиков в 2.7 раза больше ширины на вершине. Длина второго членика усиков в 1.6 раза больше ширины на вершине, в два раза меньше длины первого членика. Второй членик в 1.2 раза уже первого. Длина третьего членика усиков в 7.5 раза больше ширины на вершине, в 3.8 раза больше длины второго членика. Третий членик в 1.25 раза уже второго. Длина четвертого членика усиков в 4.0 раза больше ширины на вершине, в 1.9 раза меньше длины третьего членика. Ширина четвертого членика равна ширине третьего членика. Длина пятого членика усиков в 3.2 раза больше ширины на вершине, немного меньше длины четвертого членика. Пятый членик в 1.1 раза шире четвертого. Длина шестого членика усиков в 2.4 раза больше ширины на вершине, немного меньше длины пятого членика. Шестой членик в 1.1 раза шире пятого. Длина седьмого членика усиков в 2.0 раза больше ширины на вершине, немного меньше длины шестого членика. Ширина седьмого членика равна ширине пятого членика. Длина восьмого членика усиков примерно равна длине седьмого членика. Булава явственная, не компактная, примерно в 3 раза короче предыдущих члеников, вместе взятых. Членики булавы примерно равной длины. Длина девятого членика усиков в 1.5 раза больше ширины на вершине, немного больше длины восьмого членика. Девятый членик в 1.6 раза шире восьмого. Длина десятого членика усиков в 1.2 раза больше ширины на вершине. Десятый членик немного шире девятого. Длина 11-го членика усиков в 1.5 раза больше ширины на вершине. Одиннадцатый членик немного уже десятого. Переднеспинка с явственным боковым килем, выпуклая в вершинной трети, мелко пунктированная. Щиток довольно крупный. Надкрылья слабовыпуклые, их длина в 3.2 раза больше переднеспинки. Бороздки надкрылий слабые, с мелкими точками. Прищитковая бороздка явственная. Промежутки между бороздками широкие, плоские. Прекоксальная часть переднегруди удлиненная, ее длина немного больше передних тазиков. Посткоксальная часть переднегруди короткая. Заднегрудь длинная, уплощенная, ее длина в два раза больше длины задних тазиков. Метэпистерны узкие, их длина в 6.6 раза больше ширины на середине. Вершина брюшка не скрыта надкрыльями. Стерниты свободные, равной длины. Передние тазики расположены перед основанием переднегруди. Ноги длинные. Бедра довольно широкие. Длина передних бедер примерно в 6.2 раза больше ширины на середине. Длина средних бедер примерно в 3.1 раза больше ширины на середине. Длина задних бедер примерно в 2.7 раза больше ширины на середине. Голени довольно узкие, изогнутые перед вершиной, без явственного мукро. Длина передних голеней примерно в 9.3 раза больше ширины на середине. Длина средних голеней примерно в 6.8 раза больше ширины на середине. Длина задних голеней примерно в 6 раз больше ширины на середине. Лапки длинные. Первый и второй членики удлиненно-трапециевидные. Первый членик уже голеней. Третий членик двухлопастный. Коготковый членик длинный, с крупными коготками. Длина первого членика передних лапок в 1.8 раза больше ширины на вершине. Длина второго членика задних лапок в 1.3 раза меньше длины первого членика. Длина третьего членика задних лапок в 1.2 раза меньше длины второго членика. Длина второго членика задних лапок в 1.7 раза больше длины третьего членика.
Размеры в мм. Длина тела без головотрубки 6.9; длина головотрубки 2.5.
Материал. Голотип.
Belonotaris gratshevi (Legalov, 2010)
Arnoldibelus gratshevi: Легалов, 2010, с. 54, рис. 1в; 2011а, c. 64.
Belonotaris gratshevi: Legalov, 2012, c. 687; Gratshev, Legalov, 2014, с. 900, рис. 7 с, табл. IX , фиг. 4 ; Legalov, 2015, c. 1499.
Голотип – ПИН, № 2904/958, прямой и обратный отпечатки жука; Каратау–Михайловка; средняя–верхняя юра, карабастауская свита.
Описание (рис. 1, а, в). Длина головотрубки в 50 раз больше ее ширины на вершине, в 17.5 раза больше ширины на середине, в 12.5 раза больше ширины на основании, в 2.8 раза больше длины переднеспинки, в 2.0 раза больше головы и переднеспинки, вместе взятых. Головотрубка прямая, едва сужающаяся перед вершиной. Мандибулы удлиненные. Максиллярные пальпы длинные, примерно равны по длине мандибулам. Глаза средних размеров, округлые. Диаметр глаза немного меньше ширины головотрубки у основания. Длина виска составляет 0.6 длины глаза. Усики прикреплены перед серединой головотрубки, заходят за середину переднеспинки. Членики жгутика удлиненно-трапециевидные. Первый членик усиков овальный, его длина в 3.5 раза больше ширины на вершине. Длина второго членика усиков примерно в 2.3 раза больше ширины и примерно в 1.6 раза меньше длины первого членика. Третий–седьмой членики усиков примерно равны по ширине. Третий членик усиков удлиненный, его длина в 6.1 раза больше ширины на вершине и в 2.4 раза больше длины второго членика. Третий членик немного уже второго. Четвертый–седьмой членики усиков примерно равной длины; длина четвертого членика в 3.9 раза больше ширины на вершине и в 1.6 раза меньше длины третьего членика. Длина восьмого членика усиков в 2.5 раза больше ширины на вершине и в 1.4 раза меньше длины пятого членика. Восьмой членик немного шире пятого. Булава явственная, не компактная, ее длина примерно в 4.3 раза меньше длины предыдущих члеников, вместе взятых. Длина девятого членика усиков в 1.3 раза больше ширины на вершине и немного меньше длины восьмого членика. Девятый членик в 1.5 раза шире восьмого. Длина десятого членика усиков в 1.3 раза больше ширины на вершине и немного больше длины девятого членика. Десятый членик немного шире девятого. Длина 11-го членика усиков в 1.3 раза больше ширины на вершине и немного меньше длины десятого членика. Одиннадцатый членик немного уже десятого. Переднеспинка с явственным боковым килем, уплощенная, мелко пунктированная. Щиток довольно крупный. Надкрылья уплощенные, в 3.0 раза длиннее переднеспинки. Бороздки надкрылий слабые с мелкими точками. Промежутки между бороздками широкие, плоские. Прекоксальная часть переднегруди удлиненная, ее длина примерно в 1.7 раза больше длины передних тазиков. Посткоксальная часть переднегруди примерно в 5 раз короче прекоксальной части. Заднегрудь длинная, уплощенная, ее длина примерно в два раза больше длины задних тазиков. Вершина брюшка скрыта надкрыльями. Передние тазики расположены возле основания переднегруди. Ноги длинные. Бедра довольно широкие. Длина передних бедер примерно в 3.3 раза больше ширины на середине. Длина средних бедер примерно в 3.2 раза больше ширины на середине. Длина задних бедер примерно в 3.1 раза больше ширины на середине. Голени довольно узкие, прямые, без мукро. Длина передних и средних голеней примерно в 7.5 раза больше ширины на середине. Длина задних голеней примерно в 7.3 раза больше ширины на середине. Первый членик трапециевидный, едва шире ширины голеней. Длина первого членика средних и задних лапок в 1.1 раза больше ширины на вершине.
Размеры в мм. Длина тела без головотрубки 7.6; длина головотрубки 5.0.
Замечания. Верхняя губа у голотипа плохо видна.
Материал. Голотип.
Belonotaris rohdendorfi Legalov, 2014
Belonotaris rohdendorfi: Gratshev, Legalov, 2014, с. 902, рис. 9d , табл. VIII , фиг. 4 ; Legalov, 2015, c. 1499.
Голотип – ПИН, № 2784/1452, прямой и обратный отпечатки жука; Каратау–Михайловка; средняя–верхняя юра, карабастауская свита.
Описание (рис. 1, б, г–ж). Длина головотрубки в 12–21 раз больше ее ширины на вершине, в 13–15 раз больше ширины на середине, в 7.3–9.5 раза больше ширины на основании, в 2.3–2.4 раза больше длины переднеспинки, в 1.2–1.5 раза больше головы и переднеспинки, вместе взятых. Головотрубка прямая, слабо сужающаяся к вершине. Мандибулы удлиненные. Верхняя губа явственная, почти прямоугольная. Глаза крупные, округлые. Диаметр глаза в 1.2 раза больше ширины головотрубки у основания. Длина виска составляет примерно 0.6 длины глаза. Усики прикреплены перед серединой головотрубки, заходят за передний край переднеспинки. Членики жгутика удлиненно-трапециевидные. Длина первого членика усиков в 2.0 раза больше ширины на вершине. Второй–восьмой членики усиков примерно равны по ширине. Второй членик усиков примерно равной длины и ширины. Второй членик в 3.0 раза короче первого. Третий членик усиков удлиненный. Длина третьего членика в 5.5 раза больше ширины на вершине и в 5.0 раз больше длины второго членика. Длина четвертого членика усиков в 3.5 раза больше ширины на вершине и в 1.6 раза меньше длины третьего членика. Длина пятого членика усиков в 3.3 раза больше ширины на вершине и немного меньше длины четвертого членика. Длина шестого членика усиков в 2.8 раза больше ширины на вершине и в 1.2 раза меньше длины пятого членика. Длина седьмого членика усиков в 3.0 раза больше ширины на вершине и немного меньше длины шестого членика. Восьмой членик усиков примерно равен по длине седьмому членику. Булава явственная, не компактная, ее длина примерно в 2.7 раза меньше длины предыдущих члеников, вместе взятых. Длина девятого членика усиков в 2.0 раза больше ширины на вершине и немного больше длины восьмого членика. Девятый членик в 1.5 раза шире восьмого. Длина десятого членика усиков в 1.9 раза больше ширины на вершине и немного больше длины девятого членика. Ширина десятого членика примерно в 1.2 раза больше ширины девятого членика. Длина 11-го членика усиков в 2.0 раза больше ширины на вершине и немного меньше длины десятого членика. Ширина 11-го членика примерно в 1.5 раза меньше ширины десятого членика. Переднеспинка с явственным боковым килем, слабо выпуклая, мелко пунктированная. Щиток довольно крупный. Надкрылья слабовыпуклые, их длина в 2.6–3.0 раза больше длины переднеспинки. Бороздки надкрылий слабые, с мелкими точками. Промежутки между бороздками широкие, плоские. Прекоксальная часть переднегруди удлиненная, ее длина примерно равна длине передних тазиков или больше их. Посткоксальная часть переднегруди примерно в два или более раз короче прекоксальной части. Заднегрудь длинная, уплощенная, ее длина примерно вдвое больше длины задних тазиков. Метэпистерны довольно узкие. Вершина брюшка не скрыта надкрыльями. Вентриты свободные. Длина первого вентрита в 1.3 раза больше длины задних тазиков. Длина второго вентрита составляет 0.7 длины первого вентрита. Длина третьего вентрита едва меньше длины второго вентрита или равна ей. Четвертый вентрит в 1.1–1.2 раза короче третьего. Пятый вентрит в 1.5 раза длиннее четвертого или равен ему. Передние тазики расположены возле основания переднегруди. Ноги длинные. Бедра довольно широкие. Передние бедра немного шире задних, их длина примерно в 2.4–3.3 раза больше ширины на середине. Длина средних бедер примерно в 3.3 раза больше ширины на середине. Длина задних бедер примерно в 4.0–4.1 раза больше ширины на середине. Голени довольно узкие, почти прямые, без явственного мукро. Длина задних голеней примерно в 8.8–9.8 раза больше ширины на середине. Лапки длинные. Первый и второй членики удлиненно-трапециевидные. Длина первого членика равна ширине голеней или едва больше их. Третий членик двухлопастный. Длина первого членика передних лапок в 1.4 раза больше ширины на вершине. Длина первого членика задних лапок в 1.4 раза больше ширины на вершине. Длина второго членика задних лапок в 1.6 раз больше ширины на вершине, немного меньше длины первого членика. Ширина второго членика немного меньше ширины первого членика. Длина третьего членика задних лапок в 1.3 раза больше ширины на вершине и примерно равна длине второго членика. Третий членик задних лапок в 1.3 раза шире второго.
Размеры в мм. Длина тела без головотрубки 5.7–6.7; длина головотрубки 2.9–4.0.
Материал. Голотип и два экз.: ПИН, № 2784/1450, прямой и обратный отпечатки и ПИН, № 2384/535, обратный отпечаток.
ЗАКЛЮЧЕНИЕ
Долгоносикообразные жуки обнаружены в четырех юрских местонахождениях (рис. 2). Три из них (Даохугоу, Каратау и Шар-Тэг) почти одновозрастны и датируются границей средней–верхней юры, в то время как позднеюрский Талбрагар сильно удален от азиатских точек и находится в южном полушарии. Всего в юре обнаружено 65 видов из семейств Nemonychidae, Anthribidae и Ithyceridae (Legalov, 2015; Yu et al., 2019). Наиболее разнообразная юрская фауна Curculionoidea описана из Каратау, где встречены 62 вида из трех семейств, причем ложнослоники отмечены только из этого местонахождения (Легалов, 2011б, 2013). Nemonychidae найдены во всех местонахождениях, кроме Шар-Тэга, где обнаружен только один представитель подсемейства Baissorhynchinae семейства Ithyceridae (Грачев, Жерихин, 1999; Legalov, 2015). По одному виду немонихид описано из Даохугоу (Yu et al., 2019) и Талбрагара (Oberprieler, Oberprieler, 2012). В первом местонахождении обнаружен представитель юрского рода Belonotaris из подсемейства Eobelinae, ранее известного только из Каратау, а во втором – вид рода Talbragarus Oberprieler et Oberprieler, 2012 из подсемейства Brenthorrhininae, близкий к роду Distenorrhinus Arnoldi, 1977 (триба Distenorrhinini), обитавшему в средней–поздней юре и раннем мелу Европы и Азии (Legalov, 2015). Таким образом, наибольшее сходство между фаунами юрских местонахождений, основанное на находках представителей одного рода, наблюдается в Даохугоу и Каратау. Талбрагар и Шар-Тэг обособлены от остальных фаун. Находки трибы Distenorrhinini связывают талбрагарскую и каратаускую фауны. Сходство между фаунами Каратау и Шар-Тэга проявляется в наличии представителей семейства Ithyceridae, относящихся к разным подсемействам.
Большой интерес представляют трофические связи юрских долгоносикообразных жуков. Было высказано предположение (Crowson, 1975; Gratshev, Legalov, 2014) о связи юрских Nemonychidae с беннеттитовыми (Bennettitales), основанное на совместном нахождении их остатков в захоронениях (Kuschel, 1983). Современные немонихиды, как правило, являются монофагами (Kuschel, 1989; Kuschel, Leschen, 2011). Вероятно, и ископаемые формы были приурочены к определенным видам кормовых растений. Виды в переделах рода, скорее всего, развивались на разных видах или родах кормовых растений, однако в местонахождении Каратау обнаружено всего 10 видов Bennettitales: пять видов Otozamites и по одному виду родов Williamsoniella, Weltrichia, Ptilophyllum, Zamiophyllum и Pterophyllum (Долуденко и др., 1990). Этого явно недостаточно для 62 видов Nemonychidae. Можно предположить, что возле каратауского озера произрастало больше видов беннеттитовых, поскольку существуют примеры ископаемых долгоносикообразных жуков, развивающихся на определенных растениях, которые при этом бедно представлены или отсутствуют в местонахождениях. Например, описанные из балтийского янтаря виды рода Oxycraspedus Kuschel, 1955 (Legalov, 2016a) облигатно связаны с араукариями, а виды Ceutorhynchus Germar, 1823 (Legalov, 2013, 2016b) – с крестоцветными, но эти растения не найдены в балтийском янтаре (Czeczott, 1961). Другим примером является находка шести видов рода Dorytomus Germar, 1817 (Legalov, 2016b, 2020; Bukejs, Legalov, 2019), облигатно связанного с ивовыми, которые, тем не менее, представлены в балтийском янтаре только одним видом (Czeczott, 1961). Скорее всего, виды Nemonychidae были связаны и с голосеменными других групп, например, с представителями Cycadales, Ginkgoales, Czekanowskiales и Coniferales, найденными в этих отложениях (Долуденко и др., 1990). Для талбрагарского Talbragus averyi Oberprieler et Oberprieler, 2012 предполагается обитание на Podozamites jurassica (Podocarpaceae), основанное на большом числе остатков этого растения в местонахождении (Oberprieler, Oberprieler, 2012). Возможно, это правильное предположение, но беннеттитовые были обычны в поздней юре Австралии (McLoughlin, Pott, 2009), а их остатки отмечались во флоре Талбрагара (White, 1981). Единственная немонихида из Даохугоу могла развиваться на растениях, с которыми были связаны представители Belonotaris в Каратау. Во флоре Даохугоу известны семь видов беннеттитовых (Pott, Jiang, 2017) из родов Wielandiella, Anomozamites, Cycadolepis, Pterophyllum и Jacutiella. Общим родом для двух местонахождений является Pterophyllum, который, возможно, и являлся кормовым растением для Belonotaris daohugouensis. Другими вариантами могут быть Ginkgoites и Sphenobaiera (Ginkgoales) или Pityophyllum (Pinales), также найденные в обоих местонахождениях.
Список литературы
Арнольди Л.В. Rhynchophora // Мезозойские жесткокрылые / Ред. Родендорф Б.Б. М., 1977. С. 142–176 (Тр. Палеонтол. ин-та АН СССР. Т. 161).
Грачев В.Г., Жерихин В.В. Ревизия долгоносиков-немонихид родов Distenorrhinus и Procurculio (Insecta, Coleoptera: Nemonychidae) // Палеонтол. журн. 1995. № 2. С. 83–94.
Грачев В.Г., Жерихин В.В. Gobicar, новый позднеюрский род долгоносиков-эккоптартрид из Монголии (Insecta, Coleoptera: Eccoptarthridae) // Палеонтол. журн. 1999. № 2. С. 43–45.
Грачев В.Г., Легалов А.А. Новые мезозойские итицериды (Coleoptera, Ithyceridae) // Палеонтол. журн. 2011. № 1. С. 69–74.
Долуденко М.П., Сакулина Г.В., Пономаренко А.Г. Геологическое строение района уникального местонахождения позднеюрской фауны и флоры Айлие (Каратау, Южный Казахстан). М.: Ротапринт ГИН АН СССР, 1990. 38 с.
Легалов А.А. Обзор долгоносикообразных жуков рода Arnoldibelus Leg. из юры Казахстана (Coleoptera: Nemonychidae) // Палеонтол. журн. 2010. № 6. С. 53–55.
Легалов А.А. К познанию надсемейства Curculionoidea (Coleoptera) из юры и мела Северного Полушария // Евраз. энтомол. журн. 2011а. Т. 10. № 1. С. 63–71.
Легалов А.А. Первая находка жуков-ложнослоников (Coleoptera: Anthribidae) в юре Казахстана // Палеонтол. журн. 2011б. № 6. С. 34–37.
Легалов А.А. Обзор семейства Anthribidae (Coleoptera) из юры Каратау. Подсемейство Protoscelinae. Род Protoscelis Medvedev, 1968 // Палеонтол. журн. 2013. № 3. С. 57–65.
Полянский Б.В., Долуденко М.П. Некоторые вопросы седиментогенеза верхнеюрских карбонатных флишоидных отложений хребта Каратау (Южный Казахстан) // Литол. и полезн. ископ. 1978. № 3. С. 78–88.
Bean L.B. The leptolepid fish Cavenderichthys talbragarensis from the Talbragar Fish Bed (Late Jurassic) near Gulgong, New South Wales // Rec. West. Austral. Mus. 2006. V. 23. P. 43–76.
Bukejs A., Legalov A.A. A new species of the genus Dorytomus Germar, 1817 (Coleoptera, Curculionidae) from Baltic amber // Entomol. Fennica. 2019. V. 30. № 4. P. 173–178.
Crowson R.A. The evolutionary history of Coleoptera as documented by fossil and comparative evidence // Atti 10th Congr. Naz. Ital. Entomol. 1975. P. 47–90.
Czeczott H. The flora of the Baltic amber and its age // Prace Muz. Ziemi. 1961. V. 4. P. 119–145.
Gratshev V.G., Legalov A.A. The Mesozoic stage of evolution of the family Nemonychidae (Coleoptera, Curculionoidea) // Paleontol. J. 2014. V. 48. № 8. P. 851–944.
Gratshev V.G., Zherikhin V.V. A revision of the Nemonychid weevil subfamily Brenthorrhininae (Insecta, Coleoptera: Nemonychidae) // Paleontol. J. 1995 (1996). V. 29. № 4. P. 112–127.
Jiang B., Yao X., Niu Y. et al. Outline of the Jurassic and Cretaceous systems in western Liaoning, NE China. Hefei: Univ. of Sci. & Technol. of China Press, 2010. 84 p.
Kuschel G. Past and present of the relict family Nemonychidae (Coleoptera, Curculionoidea) // Geol. J. 1983. V. 7. № 6. P. 499–504.
Kuschel G. The nearctic Nemonychidae (Coleoptera, Curculionoidea) // Entomol. Scand. 1989. V. 20. P. 121–171.
Kuschel G., Leschen R.A.B. Phylogeny and taxonomy of the Rhinorhynchinae (Coleoptera: Nemonychidae) // Invertebr. Syst. 2011. V. 24. P. 573–615.
Legalov A.A. Annotated checklist of fossil and recent species of the family Belidae (Coleoptera) from the world fauna // Amur. Zool. Zhurn. 2009. V. 1. № 4. P. 296–324.
Legalov A.A. Fossil history of Mesozoic weevils (Coleoptera: Curculionoidea) // Insect Sci. 2012. V. 19. № 6. P. 683–698.
Legalov A.A. New and little known weevils (Coleoptera: Curculionoidea) from the Paleogene and Neogene // Hist. Biol. 2013. V. 25. № 1. P. 59–80.
Legalov A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea) // Paleontol. J. 2015. V. 49. № 13. P. 1442–1513.
Legalov A.A. Two new genera and four new species of fossil weevils (Coleoptera: Curculionoidea) in Baltic amber // Entomol. Fennica. 2016a. V. 27. № 2. P. 57–69.
Legalov A.A. New weevils (Curculionidae) in Baltic amber // Paleontol. J. 2016b. V. 50. № 9. P. 970–985.
Legalov A.A. Annotated key to weevils of the World. Part 1. Families Nemonychidae, Anthribidae, Belidae, Ithyceridae, Rhynchitidae, Brachyceridae and Brentidae // Ukr. J. Ecol. 2018. V. 8. № 1. P. 780–831.
Legalov A.A. A review of the Curculionoidea (Coleoptera) from European Eocene ambers // Geosciences. 2020. V. 10. № 1(16). P. 1–74.
Marvaldi A.E., Ferrer M.S. 3.3. Belidae Schoenherr, 1826 // Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles. V. 3: Morphology and Systematics (Phytophaga) / Eds. R.A.B. Leschen, R.G. Beutel. Berlin/Boston: de Gruyter, 2014. P. 316–327.
Marvaldi A.E., Lyal C.H.C., Oberprieler R.G. et al. Phylogeny of the Oxycoryninae s. l. (Coleoptera: Belidae) and evolution of host-plant associations // Invertebr. Syst. 2006. V. 20. P. 447–476.
McLoughlin S., Pott C. The Jurassic flora of Western Australia // GFF. 2009. V. 131. № 1–2. P. 113–136.
Oberprieler R.G., Oberprieler S.K. Talbragarus averyi gen. et sp. n., the first Jurassic weevil from the Southern Hemisphere (Coleoptera: Curculionoidea: Nemonychidae) // Zootaxa. 2012. V. 3478. P. 256–266.
Ponomarenko A.G., Aristov D.S., Bashkuev A.S. et al. Upper Jurassic Lagerstätte Shar Teg, Southwestern Mongolia // Paleontol. J. 2014. V. 48. № 14. P. 1573–1682.
Pott C., Jiang B. Plant remains from the Middle-Late Jurassic Daohugou site of the Yanliao Biota in Inner Mongolia, China // Acta Palaeobot. 2017. V. 57. № 2. P. 185–222.
Scotese C.R. An Atlas of Paleogeographic Maps: The Seas Come In and the Seas Go Out // Ann. Rev. Earth Planet. Sci. 2021. V. 49. P. 669–718.
Yu Y., Davis S.R., Shih C. et al. The earliest fossil record of Belidae and its implications for the early evolution of Curculionoidea (Coleoptera) // J. Syst. Palaeontol. 2019. V. 17. P. 2105–2117.
White M.E. Revision of the Talbagar Fish Bed flora (Jurassic) of New South Wales // Rec. Austral. Mus. 1981. V. 33. № 15. P. 695–721.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал