

Палеонтологический журнал, 2022, № 6, стр. 3-13
Апт–альбские устрицы юго-западных отрогов Гиссарского хребта. Статья 2. Роды Ceratostreon Bayle, Pseudogyra Mirkamalov и Costagyra Vialov
И. Н. Косенко a, b, c, *, Е. К. Метелкин a, b, **
a Институт нефтегазовой геологии и геофизики им. А. А. Трофимука СО РАН
630090 Новосибирск, Россия
b Новосибирский государственный университет
630090 Новосибирск, Россия
c Нанкинский институт геологии и палеонтологии Китайской академии наук
210008 Нанкин, Китай
* E-mail: KosenkoIN@ipgg.sbras.ru
** E-mail: MetelkinEK@ipgg.sbras.ru
Поступила в редакцию 15.02.2022
После доработки 28.02.2022
Принята к публикации 30.03.2022
- EDN: AMPRBO
- DOI: 10.31857/S0031031X2206006X
Аннотация
Представлены результаты ревизии устриц, относящихся к родам Ceratostreon Bayle, 1878, Pseudogyra Mirkamalov, 1971 и Costagyra Vialov, 1936 (Gryphaeidae, Exogyrinae), из апта и альба юго-западных отрогов Гиссарского хребта. Монографически описаны виды Ceratostreon trigonalis (Mirkamalov, 1971), Pseudogyra pennata Mirkamalov, 1971 и Costagyra luppovi (Mirkamalov, 1963).
ВВЕДЕНИЕ
Настоящая статья является продолжением серии статей, посвященных апт–альбским устрицам Гиссарского хребта. В первой статье (Метелкин, Косенко, 2021) приведены сведения по стратиграфии апта–сеномана юго-западных отрогов Гиссарского хребта, а также представлены результаты ревизии устриц рода Amphidonte Fisсher de Waldheim, 1829. Настоящая статья посвящена ревизии устриц, относящихся к родам Ceratostreon Bayle, 1878, Pseudogyra Mirkamalov, 1971 и Costagyra Vialov, 1936 (Gryphaeidae, Exogyrinae). Необходимость переописания известных видов связана с тем, что в результате изучения нового палеонтологического материала с представительными выборками удалось пересмотреть пределы изменчивости отдельных видов, в результате чего ряд видовых названий сведен в синонимику. Кроме того, уточнено стратиграфическое распространение и пересмотрена родовая принадлежность описываемых видов с учетом последних данных по классификации устриц.
Материалом для исследования послужила представительная коллекция устриц из апт–сеноманских отложений юго-западных отрогов Гиссарского хребта, собранная в разные годы сотрудниками Всероссийского научно-исследовательского геологического ин-та им. А.П. Карпинского (ВСЕГЕИ) и переданная в 2015 г. Т.Н. Богдановой для изучения в Ин-т нефтегазовой геологии и геофизики им. А.А. Трофимука (ИНГГ) СО РАН. Рисунок с картой расположения местонахождений изученных раковин устриц приведен в первой статье (Метелкин, Косенко, 2021, рис. 1 ).
Изученная коллекция передана на хранение в центр коллективного пользования “Геохрон” ИНГГ СО РАН.
ОПИСАНИЕ ТАКСОНОВ
НАДСЕМЕЙСТВО OSTREOIDEA RAFINESQUE, 1815
СЕМЕЙСТВО GRYPHAEIDAE VIALOV, 1936
ПОДСЕМЕЙСТВО EXOGYRINAE VIALOV, 1936
Триба Amphidonteini Vialov, 1983
Диагноз. Раковины от маленького до среднего, реже крупного размера, вытянутые в высоту, уховидных очертаний. Макушка завернута в слабую спираль; плоскость завивания спирали приближена к плоскости смыкания створок; ось завивания спирали приблизительно перпендикулярна плоскости смыкания створок. Лигаментные площадки экзогироидного11 типа, иногда выпрямляющиеся на поздних стадиях роста. Хоматы прямые, многочисленные, расположены по периметру (или его большей части) смычного края.
Состав трибы: роды Amphidonte, Ceratostreon, Vultogryphaea, Palaeogyra и Pseudogyra.
Замечания. Экзогиры были подразделены на таксоны надродового ранга О.С. Вяловым (1983). Он поднял ранг изначально выделенного им подсемейства Exogyrinae (Вялов, 1936) до семейства и подразделил его, в свою очередь, на три подсемейства: Exogyrinae Vialov, 1936, Amphidonteinae Vialov, 1983 и Ceratostreoninae Vialov, 1983. Вялов принял тип скульптуры раковины как основу для деления экзогир на подсемейства. Подсемейство Amphidonteinae объединило роды Amphidonte Fischer de Waldheim, 1829, Aetostreon Bayle, 1878 и Gryphaeostrea Conrad, 1865, для которых характерно полное отсутствие радиальной скульптуры (Вялов, 1984). Однако еще Г. Стензелем (Stenzel, 1971) было показано, что наружные признаки раковин, сильно зависящие от среды обитания моллюска, такие как скульптура, не могут быть использованы для систематики таксонов надродового ранга. Использование Вяловым скульптуры раковины в качестве признака для разделения на подсемейства привело к тому, что в подсемейство Amphidonteinae были объединены роды, относящиеся в современной систематике к разным подсемействам (Malchus, 1990; Carter et al., 2011).
Другой вариант классификации подсемейства Exogyrinae был предложен Н. Мальхусом (Malchus, 1990). Он разделил подсемейство на две трибы: Nanogyrini Malchus, 1990 и Exogyrini Vialov, 1936. В основу деления был положен комплекс признаков, включающий в себя форму раковины, тип хомат и степень их развития, геометрию завивания макушек, микроструктуру раковины. В трибу Nanogyrini Мальхус включил роды Nanogyra Beurlen, 1958, Aetostreon и Amphidonte (с подродами Amphidonte s. str. и Ceratostreon Bayle, 1878). Позднее на основании изучения микроструктуры раковины Мальхус сделал вывод о том, что род Aetostreon ближе к подсемейству Gryphaeostreinae, чем к Exogyrinae (Malchus, 1998). В современной систематике двустворчатых моллюсков (Carter et al., 2011) принято деление подсемейства Exogyrinae на две трибы: Exogyrini Vialov, 1936 и Amphidonteini Vialov, 1983 (=Nanogyrini Malchus, 1990). Мы полностью поддерживаем подход Мальхуса к систематике экзогир, основанный на комплексном изучении всех морфологических признаков раковин и микроструктуры, но считаем, что род Nanogyra стоит исключить из трибы Amphidonteini и рассматривать в составе монотипической трибы Nanogyrini. Основанием для этого служит отсутствие у представителей рода Nanogyra хомат, характерных для всех остальных экзогир.
Род Ceratostreon Bayle, 1878
Ceratostreon: Bayle, 1878, табл. CXXXIV , фиг. 1, 2 ; Stenzel, 1971, с. N1119; Невесская и др., 2013, с. 191.
Amphidonte (Ceratostreon): Malchus, 1990, с. 110; Aqrabawi, 1993, с. 62.
Типовой вид – Ostrea plicifera Dujardin, 1837 (=O. matheroniana d’Orbigny, 1846), сенон Франции.
Диагноз. Раковины мелких и средних размеров, обычно вытянутые в высоту, с радиально-ребристой скульптурой на обеих створках (иногда скульптура очень слабо выражена, вплоть до почти полного отсутствия), имеющие прямые хоматы, развитые вдоль всего внутреннего периметра смычного края. Лигаментная площадка экзогироидного типа, у крупных раковин может выпрямляться на поздних стадиях роста.
Видовой состав: Ceratostreon flabellatum (Goldfuss, 1833), C. tuberculiferum (Koch et Dunker, 1837), C. pliciferum (Dujardin, 1837), C. boussingaulti (d’Orbigny, 1842), C. spinosum (Matheron, 1842), C. texanum (Roemer, 1849), C. minos (Coquand, 1869), C. fragilis Bogdanova, 1980 и C. trigonalis (Mirkamalov, 1971).
Замечания. Вслед за Мальхусом (Malchus, 1990) многие исследователи рассматривают род Ceratostreon в качестве подрода рода Amphidonte. Однако на наш взгляд, эти два рода представляют собой две хорошо прослеживаемые с начала и до конца мела обособленные филетические линии, морфологические различия между которыми выражены в наличии в разной степени проявленной радиальной скульптуры у представителей рода Ceratostreon и отсутствии таковой у представителей рода Amphidonte, в связи с чем мы рассматриваем Ceratostreon как самостоятельный род. Этой же точки зрения придерживается М. Купер (Cooper, 1997).
Ceratostreon trigonalis (Mirkamalov, 1971)
Табл. I , фиг. 16–23
Exogyra trigonalis: Миркамалов, 1971а, с. 119, табл. I , фиг. 1–9 ; 1986, с. 65, табл. XVII , фиг. 1–6 .
Голотип – Ин-т геологии и разведки нефтяных и газовых месторождений (ИГИРНИГМ), № М1/202, целая раковина; Туркменистан, юго-западные отроги Гиссарского хребта, пос. Койтендаг (=Чаршанга); апт, кундалянгтауская свита (Миркамалов, 1971а, табл. I , фиг. 1, 2 ).
Объяснения к таблице I
Юго-Восточный Туркменистан и Южный Узбекистан, юго-западные отроги Гиссарского хребта. Все изображения даны в натуральную величину.
Фиг. 1–15 . Costagyra luppovi (Mirkamalov): 1 – экз. № 2105/428, левая створка; 2 – экз. № 2105/421, левая створка; 3 – экз. № 2105/438, левая створка; 4 – экз. № 2105/431, левая створка; Кундалянгтау; средний альб, зона Hoplites dentatus, лучакская свита; 5 – экз. № 2105/412, левая створка; колодец Бегляр; средний альб, зона Hoplites dentatus, лучакская свита; 6 – экз. № 2105/539, левая створка; 7 – экз. № 2105/595, левая створка; 8 – экз. № 2105/602, левая створка; Кундалянгтау; верхний альб, зона Anahoplites rossicus, лучакская свита; 9 – экз. № 2105/434, левая створка; местонахождение то же; средний альб, зона Hoplites dentatus, лучакская свита; 10 – экз. № 2105/600, левая створка; местонахождение то же; верхний альб, зона Anahoplites rossicus, лучакская свита; 11 – экз. № 2105/408, левая створка; колодец Бегляр; средний альб, зона Hoplites dentatus, лучакская свита; 12 – экз. № 2105/644, левая створка; местонахождение то же; верхний альб, зона Anahoplites rossicus, лучакская свита; 13 – экз. № 2105/418, левая створка; Кундалянгтау; средний альб, зона Hoplites dentatus, лучакская свита; 14 – экз. № 2105/590, левая створка; местонахождение то же; верхний альб, зона Anahoplites rossicus, лучакская свита; 15 – экз. № 2105/612, левая створка; ущелье Газдагана, верхний альб, зона Anahoplites rossicus, лучакская свита.
Фиг. 16–23 . Ceratostreon trigonalis (Mirkamalov): 16 – экз. № 2105/355: 16а – вид с внешней стороны левой створки, 16б – вид с внешней стороны правой створки; 17 – экз. № 2105/318: 17а – вид с внешней стороны левой створки, 17б – вид с внешней стороны правой створки; Кундалянгтау; нижний апт, кундалянгтауская свита; 18 – экз. № 2105/538: 18а – левая створка, 18б – правая створка; 19 – экз. № 2105/534: 19а – левая створка, 19б – правая створка; 20 – экз. № 2105/529: 20а – левая створка, 20б – правая створка; 21 – экз. № 2105/504: 21а – правая створка, 21б – левая створка; 22 – экз. № 2105/537: 22а – левая створка, 22б – правая створка; 23 – экз. № 2105/519, левая створка; местонахождение то же; средний альб, лучакская свита.
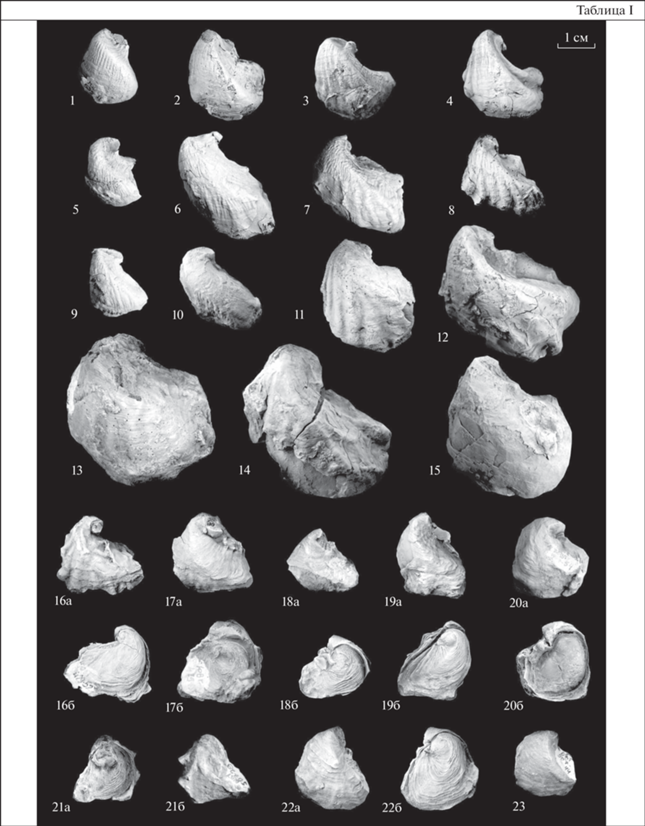
Описание. Раковина маленьких размеров (высотой до 23 мм, длиной до 21 мм), треугольно-овальной формы, неравностворчатая, неравносторонняя. Левая створка слабо выпуклая (коэффициент выпуклости Вып/Д от 0.33 до 0.64; среднее значение 0.48), треугольно-овального очертания. Передний край округло-выпуклый, задненижний край оттянут, задний край прямой, иногда вогнутый или слегка выпуклый. Макушка маленькая, спирально завернута назад, не выступает за плоскость смыкания створок. Площадка прирастания маленькая, расположена на заднем крае вблизи макушки. От макушки к задненижнему краю протягивается киль. Скульптура левой створки представлена как концентрическими линиями роста, так и радиальной ребристостью. Радиальная ребристость выражена в различной степени: для некоторых раковин она проявляется лишь в краевых частях, а ближе к килю сглаживается. Ребристость наиболее выражена на переднем крае раковины. На пересечении ребер с килем иногда образуются бугорки. Правая створка плоская, уховидного очертания. Передненижний край округлый, задний прямой, иногда с небольшим выступом – ушком. Макушка маленькая, спирально завернута назад. Наружная поверхность покрыта концентрическими линиями нарастания, расположенными достаточно плотно друг относительно друга.
Размеры в мм и отношения:
| № экз. | В | Д | Вып | В/Д | Вып/Д |
|---|---|---|---|---|---|
| 2105/311 | 19 | 17 | 7 | 1.12 | 0.41 |
| 2105/314 | 19 | 16 | 10 | 1.19 | 0.63 |
| 2105/320 | 19 | 17 | 8 | 1.12 | 0.47 |
| 2105/326 | 17 | 16 | 9 | 1.06 | 0.56 |
| 2105/333 | 21 | 18 | 10 | 1.17 | 0.56 |
| 2105/352 | 19 | 18 | 6 | 1.06 | 0.33 |
| 2105/354 | 19 | 15 | 8 | 1.27 | 0.53 |
| 2105/355 | 23 | 21 | 10 | 1.10 | 0.48 |
| 2105/356 | 17 | 14 | 9 | 1.21 | 0.64 |
| 2105/502 | 17 | 16 | 6 | 1.06 | 0.38 |
| 2105/511 | 19 | 17 | 9 | 1.12 | 0.53 |
| 2105/512 | 21 | 21 | 9 | 1.00 | 0.43 |
| 2105/513 | 14 | 15 | 6 | 0.93 | 0.40 |
| 2105/519 | 17 | 17 | 7 | 1.00 | 0.41 |
| 2105/521 | 23 | 21 | 10 | 1.10 | 0.48 |
| 2105/527 | 19 | 20 | 11 | 0.95 | 0.55 |
| 2105/529 | 21 | 19 | 8 | 1.11 | 0.42 |
| 2105/537 | 22 | 21 | 11 | 1.05 | 0.52 |
| 2105/544 | 23 | 20 | 9 | 1.15 | 0.45 |
| 2105/581 | 18 | 18 | 9 | 1.00 | 0.50 |
Изменчивость. Изменчивости в значительной степени подвержена скульптура левой створки, которая варьирует от грубых радиальных ребер и бугорков до полного их отсутствия. Кроме того, изменчивости подвержены ширина переднего поля левой створки и положение киля.
Сравнение. Описываемый вид отличается от большинства представителей рода Ceratostreon слабо проявленной радиальной скульптурой, а также более широкими очертаниями раковины.
Замечания. Определенное сходство с описываемым видом имеют экзогиры Costagyra luppovi (Mirkamalov, 1963), встречающиеся в тех же стратиграфических горизонтах юго-западных отрогов Гиссарского хребта. Ceratostreon trigonalis отличается от них меньшими размерами, меньшей вытянутостью в высоту и меньшей выпуклостью левой створки, более оттянутым задненижним краем и менее выраженным килем, а также менее развитыми радиальными ребрами, которые иногда вовсе отсутствуют. От совместно встречающихся представителей рода Pseudogyra описываемый вид отличается развитыми спирально закрученными макушками, гораздо более сильной закрученностью раковины на ранних стадиях онтогенеза, что хорошо видно по линиям роста на правых створках, и в целом большей шириной и выпуклостью раковин.
Распространение. Нижний апт–средний альб юго-западных отрогов Гиссарского хребта.
Материал. 24 раковины, 8 левых и 3 правые створки из кундалянгтауской свиты (5 раковин и 3 левые створки из Кампрекского ущелья; 8 раковин, 1 левая и 1 правая створка из Гаурдака; 11 раковин, 4 левые и 2 правые створки из Кундалянгтау); 30 раковин, 15 левых и 2 правые створки из лучакской свиты (Кундалянгтау).
Род Pseudogyra Mirkamalov, 1971
Gyrostrea (Pseudogyra) subgen. nov.: Миркамалов, 1971б, с. 37.
Pseudogyra: Миркамалов, 1986, с. 53.
Peilinia: Kues, 1997, с. 826.
Типовой вид – Gyrostrea (Pseudogyra) pennata Mirkamalov, 1971, апт юго-западных отрогов Гиссарского хребта.
Диагноз. Раковины мелких и средних размеров, уплощенные, узкие серповидные или вытянутые в высоту. Макушки маленькие, не выступающие. Радиальная скульптура развита только на левых створках и представлена выраженными в различной степени ребрами и складками. Форма лигаментных площадок варьирует от экзогироидной до гиростреоидной, с тенденцией к выпрямлению у более поздних форм. Хоматы простые, прямые, развиты вдоль всего периметра смычного края.
Видовой состав: Pseudogyra quadriplicata (Shumard, 1860), P. turkmenica (Luppov, 1960), P. pennata Mirkamalov, 1971, P. levicostata (Kues, 1997).
Замечания. Изначально род Pseudogyra был выделен Х.Х. Миркамаловым (1971б) в ранге подрода в составе рода Gyrostrea. Позднее Миркамалов (1986) повысил ранг таксона до рода и рассматривал роды Gyrostrea и Pseudogyra как регрессивную ветвь экзогирин, для которой характерно выпрямление лигаментной площадки (преобладание гиростреоидного типа). С. Френе (Freneix, 1982) на основе изучения микроструктуры раковины показала, что род Gyrostrea должен быть включен в подсемейство Crassostreinae. Более поздние исследования также показали, что род Gyrostrea должен относиться не к семейству Gryphaeidae, а к семейству Flemingostreidae (Malchus, 1990; Косенко, Метелкин, 2020). Позднее Б. Куэс (Kues, 1997) выделил в самостоятельный род Peilinia Kues, 1997 группу видов мелких удлиненных устриц с радиальной скульптурой, происходящих из альба Нью-Мексико (США), ранее рассматриваемых в составе родов Alectryonia, Rastellum, Lopha. Основные морфологические признаки рода Peilinia, данные в диагнозе, совпадают с признаками рода Pseudogyra, в связи с чем Peilinia принимается нами как младший синоним рода Pseudogyra.
Относительно положения рода Pseudogyra в системе Ostreoidea мы придерживаемся точки зрения Миркамалова (1971б, 1986) (не разделяя его взгляды на положение рода Gyrostrea в системе Ostreoidea) и считаем этот род регрессивной ветвью экзогирин. В пользу этого свидетельствует сходство морфологии ранних псевдогир из валанжина–баррема Западного Туркменистана, описанных В.А. Прозоровским (1960) (см. ниже, раздел “Сравнение”), также изученных нами, с представителями рода Ceratostreon. Причем в крупных выборках присутствуют как формы, практически не отличимые от цератостреонов, так и близкие к типичным псевдогирам, а также экземпляры с переходной морфологией раковин.
Pseudogyra pennata Mirkamalov, 1971
Табл. II , фиг. 1–15
Gyrostrea (Pseudogyra) pennata: Миркамалов, 1971б, с. 37, табл. III , фиг. 1–4 .
Gyrostrea (Pseudogyra) cornuta: Миркамалов, 1971б, с. 39, табл. III , фиг. 5, 6 .
Gyrostrea (Pseudogyra) lunata: Миркамалов, 1971б, с. 40, табл. III , фиг. 11, 12 .
Gyrostrea (Pseudogyra) sagitta: Миркамалов, 1971б, с. 38, табл. III , фиг. 7–10 .
Pseudogyra pennata: Миркамалов, 1986, с. 71, табл. X , фиг. 6–10 .
Pseudogyra sagitta: Миркамалов, 1986, с. 71, табл. X , фиг. 13, 14 .
Pseudogyra cornuta: Миркамалов, 1986, с. 72, табл. X , фиг. 11, 12 .
Голотип – Ин-т геологии и разведки нефтяных и газовых месторождений (ИГИРНИГМ), № М1/170, целая раковина; Туркменистан, юго-западные отроги Гиссарского хребта, пос. Койтендаг (=Чаршанга); нижний апт, кундалянгтауская свита.
Объяснения к таблице II
Юго-Восточный Туркменистан, юго-западные отроги Гиссарского хребта. Все изображения (кроме 15в) даны в натуральную величину.
Фиг. 1–15 . Pseudogyra pennata Mirkamalov: 1 – экз. № 2105/371: 1а – левая створка, 1б – правая створка; Кампрекское ущелье; нижний апт, кундалянгтауская свита; 2 – экз. № 2105/580, левая створка; Кундалянгтау; средний альб, лучакская свита; 3 – экз. № 2105/270: 3а – левая створка, 3б – правая створка; 4 – экз. № 2105/247, правая створка; 5 – экз. № 2105/250: 5а – левая створка, 5б – правая створка; Кансай, Гаурдак; нижний апт, кундалянгтауская свита; 6 – экз. № 2105/368, левая створка; Кампрекское ущелье; нижний апт, кундалянгтауская свита; 7 – экз. № 2105/577, левая створка; 8 – экз. № 2105/557, левая створка; Кундалянгтау; средний альб, лучакская свита; 9 – экз. № 2105/276, левая створка; Кансай, Гаурдак; нижний апт, кундалянгтауская свита; 10 – экз. № 2105/308: 10а – левая створка, 10б – правая створка; Кундалянгтау; нижний апт, кундалянгтауская свита; 11 – экз. № 2105/552, левая створка; 12 – экз. № 2105/558, левая створка; 13 – экз. № 2105/553, левая створка; 14 – экз. № 2105/574, левая створка; 15 – экз. № 2105/466: 15а – правая створка, 15б – левая створка, 15в – вид на хоматы и отпечаток мускула-замыкателя (×2); местонахождение то же; средний альб, лучакская свита.
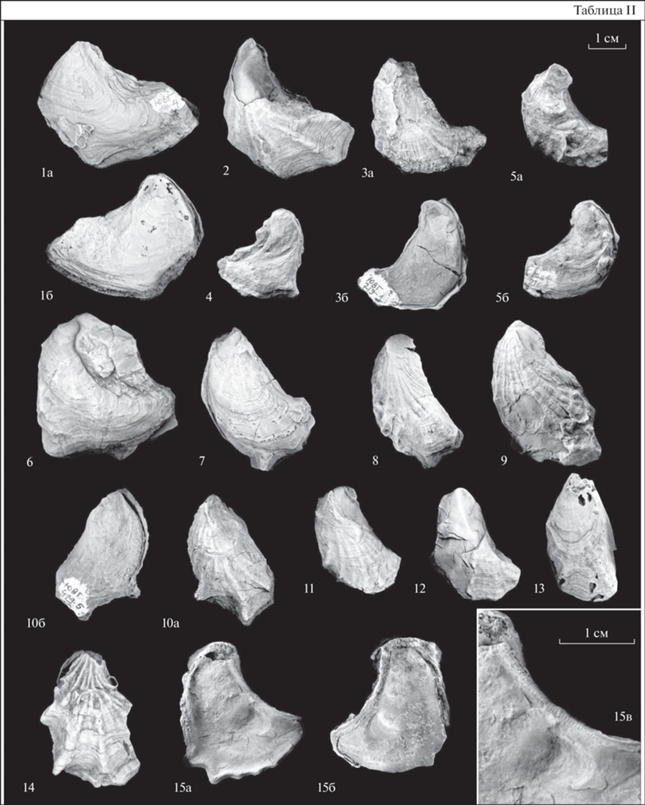
Описание. Раковина от маленьких до средних размеров (высотой до 43 мм, длиной до 38 мм), вытянутая в высоту (коэффициент вытянутости в высоту В/Д от 1 до 1.64; среднее значение 1.32), неравносторонняя, неравностворчатая, слабо выпуклая (коэффициент выпуклости от 0.26 до 0.55, среднее значение 0.41). Левая створка слабо выпуклая, от треугольного до серповидного очертания. У некоторых экземпляров на передненижнем крае створки радиальные ребра выступают за смычной край и образуют небольшое расширение. Задний край створки вогнутый. Макушка маленькая, спирально завернута назад. Площадка прирастания небольшая, расположена на макушке или отсутствует. Киль отсутствует. Наружная поверхность створки покрыта радиальными ребрами, степень выраженности которых сильно варьирует, вплоть до почти полного отсутствия, а также концентрическими линиями роста. Иногда в местах пересечения радиальных ребер и концентрических линий роста образуются бугорки. Лигаментная площадка гиростреоидного типа. Примакушечная полость отсутствует. Отпечаток мускула-замыкателя треугольно-овальной формы, смещен к нижнему краю. Смычной край очень узкий, по всему его периметру расположены простые хоматы. Отпечаток мускула Квенштедта не наблюдается. Правая створка повторяет очертания левой, плоская. Макушка маленькая, спирально завернута назад. К переднему краю створки приурочены реликтовые хоматы. Наружная поверхность покрыта концентрическими линиями роста, радиальная ребристость не проявлена. Смычной край узкий, по его периметру развиты простые хоматы. Отпечаток мускула-замыкателя треугольно-овального очертания, небольших размеров, смещен к задненижнему краю.
Размеры в мм и отношения
| № экз. | В | Д | Вып | В/Д | Вып/Д |
|---|---|---|---|---|---|
| 2105/240 | 37 | 24 | 12 | 1.54 | 0.50 |
| 2105/245 | 35 | 32 | 12 | 1.09 | 0.38 |
| 2105/270 | 32 | 28 | 10 | 1.14 | 0.36 |
| 2105/276 | 41 | 25 | 11 | 1.64 | 0.44 |
| 2105/308 | 36 | 22 | 12 | 1.64 | 0.55 |
| 2105/368 | 41 | 37 | 14 | 1.11 | 0.38 |
| 2105/371 | 37 | 37 | 10 | 1.00 | 0.27 |
| 2105/372 | 37 | 35 | 13 | 1.06 | 0.37 |
| 2105/466 | 35 | 33 | 15 | 1.06 | 0.45 |
| 2105/549 | 41 | 38 | 10 | 1.08 | 0.26 |
| 2105/550 | 43 | 35 | 14 | 1.23 | 0.40 |
| 2105/557 | 33 | 22 | 10 | 1.50 | 0.45 |
| 2105/568 | 39 | 25 | 11 | 1.56 | 0.44 |
| 2105/572 | 40 | 27 | 9 | 1.48 | 0.33 |
| 2105/574 | 35 | 27 | 12 | 1.30 | 0.44 |
| 2105/575 | 36 | 26 | 13 | 1.38 | 0.50 |
| 2105/577 | 37 | 26 | 12 | 1.42 | 0.46 |
| 2105/579 | 40 | 26 | 11 | 1.54 | 0.42 |
| 2105/580 | 41 | 32 | 11 | 1.28 | 0.34 |
Изменчивость. Изменчивости подвержены, главным образом, скульптура левой створки и очертания раковины. Скульптура левой створки варьирует от четко выраженных радиальных ребер до почти полного их отсутствия. Очертания раковины варьируют от серповидных до вытянутых в высоту треугольно-овальных. Весь диапазон изменчивости наблюдается на выборках раковин из одних и тех же слоев.
Сравнение. Наибольшее сходство описываемый вид имеет с P. levicostata (Kues, 1997) (Kues, 1997, с. 827, фиг. 4.6–4.18 ) из альба южных штатов США. Отличия заключаются в более развитой радиальной скульптуре у описываемого вида, хотя некоторые экземпляры практически неотличимы от североамериканского вида.
От P. quadriplicata (Shumard, 1860) (Kues, 1997, фиг. 4.1–4.5 ) из альба южных штатов США описываемый вид отличается более узкими раковинами и отсутствием мелких радиальных ребер на всей поверхности левых створок (хотя у некоторых экземпляров они и присутствуют между более грубыми ребрами).
От видов P. turkmenica (Luppov, 1960) (Прозоровский, 1960, табл. 18 , фиг. 2 ), P. balkhanica (Prosorovsky, 1960) (там же, табл. 18 , фиг. 3 ) и P. geokderensis (Prosorovsky, 1960) (там же, табл. 19 , фиг. 1 ) из валанжина–баррема Западного Туркменистана (изначально описанных в составе рода Exogyra), которые, на наш взгляд, являются вариациями одного вида, описываемый вид отличается меньшими размерами, меньшей выпуклостью раковины и менее развитыми радиальными ребрами.
Замечания. Изменчивость очертаний и скульптуры раковин настолько сильная, что Миркамалов (1971б) выделил четыре вида, происходящих из одного стратиграфического уровня: Pseudogyra pennata Mirkamalov, 1971, P. lunata Mirkamalov, 1971, P. sagitta Mirkamalov, 1971 и P. cornuta Mirkamalov, 1971. В нашей коллекции все выделенные Миркамаловым виды также распознаются в выборках из одних и тех же стратиграфических интервалов, при этом между всеми “видами” Миркамалова имеются переходные формы; поэтому эти виды сведены нами в синонимику вида P. pennata.
Распространение. Апт–альб юго-западных отрогов Гиссарского хребта.
Материал. 23 раковины, 25 левых и 3 правые створки из кундалянгтауской свиты (13 раковин, 11 левых и 2 правые створки из Гаурдака; 5 раковин, 9 левых и 1 правая створка из Кампрекского ущелья; 5 раковин и 5 левых створок из Кундалянгтау), 2 левые створки из каламазарской свиты (1 левая створка из Кампрекского ущелья и 1 левая створка из колодца Огулбек), 18 раковин, 18 левых и 3 правые створки из лучакской свиты (Кундалянгтау).
Триба Exogyrini Vialov, 1936
Диагноз. Раковины чаще всего треугольно-овальных очертаний, сильно неравностворчатые. Левые створки чашеобразной формы, сильно выпуклые, правые плоские или слегка вогнутые. Макушка завернута в сильно закрученную спираль, плоскость завивания которой находится под большим углом, часто близким к прямому, к плоскости смыкания створок. Хоматы развиты только вблизи лигаментной площадки. Лигаментные площадки экзогироидного типа.
Состав трибы: роды Exogyra, Costagyra, Ilymatogyra и Rhynchostreon.
Род Costagyra Vialov, 1936
Costagyra: Вялов, 1936, с. 22; 1984, с. 36.
Exogyra (Costagyra): Stenzel, 1971, с. N1117; Malchus, 1990, с. 134; Aqrabawi, 1993, с. 67.
Exogyra (pars.): Невесская и др., 2013, с. 189.
Типовой вид – Exogyra olisiponensis Sharpe, 1850, сеноман Португалии.
Диагноз. Раковины мелких и средних размеров, резко неравностворчатые, треугольно-овальных очертаний. Левые створки сильно выпуклые, правые створки плоские. Макушки левых створок закручены в геликоидальную спираль, плоскость закручивания спирали расположена под большим углом к плоскости смыкания створок. Радиальная скульптура развита на левых створках, реже на правых, и представлена грубыми ребрами, иногда осложнена более тонкими радиальными ребрами. Лигаментные площадки экзогироидного типа. Хоматы развиты вблизи лигаментной площадки.
Видовой состав: Costagyra olisiponensis (Sharpe, 1850), C. pellicoi (Verneuil et Collomb, 1853), C. luppovi (Mirkamalov, 1963), C. canensis (Mirkamalov, 1966).
Замечания. Многие исследователи рассматривают род Costagyra в качестве подрода Exogyra (Stenzel, 1971; Malchus, 1990; Aqrabawi, 1993 и др.). Главным отличием между двумя этими родами является характер скульптуры левых створок. Для рода Costagyra характерны резкие редкие радиальные ребра. Для Exogyra также характерны разные варианты радиальной скульптуры, однако резкие радиальные ребра не типичны. Кроме того, расцвет рода Exogyra пришелся на конец позднего мела (кампан–маастрихт), в то время как род Costagyra достиг своего расцвета в сеномане–туроне. На этом основании мы рассматриваем род Costagyra в качестве самостоятельного рода, морфологически близкого роду Exogyra. Такой же точки зрения придерживается Купер (Cooper, 1995).
Costagyra luppovi (Mirkamalov, 1963)
Табл. I , фиг. 1–15
Exogyra luppovi: Миркамалов, 1963, с. 134; 1966, с. 56, табл. III , фиг. 1–6 .
Exogyra localis var. subtypica: Мордвилко, 1963, с. 241, табл. VII , фиг. 8–10 .
Exogyra alaeformis: Миркамалов, 1966, с. 52, табл. 1 , фиг. 8–15 ; 1986, с. 66, табл. XVII , фиг. 7–10 .
Голотип – Ин-т геологии и разведки нефтяных и газовых месторождений (ИГИРНИГМ), № 69/102, целая раковина; юго-западные отроги Гиссарского хребта, р. Кан; верхний альб.
Описание. Раковина средних размеров (высотой до 42 мм, длиной до 38 мм), треугольно-овального, практически полулунного очертания, неравностворчатая, неравносторонняя, вытянутая в высоту (коэффициент вытянутости в высоту В/Д от 1 до 1.33, среднее значение 1.13), сильно выпуклая (коэффициент выпуклости от 0.32 до 1, среднее значение 0.59). Левая створка сильно выпуклая, с вогнутым задним краем, полукругло-выгнутым передним. Макушка массивная, сильно выступающая, спирально завернута назад и внутрь. Площадка прирастания небольшая, расположена у заднего края створки в районе макушки. Дугообразно от макушки к задненижнему краю раковины проходит закругленный киль. Иногда на заднем поле параллельно килю присутствует желобок – сулькус. Наружная поверхность створки покрыта радиальными ребрами. У многих раковин скульптура осложнена гораздо более тонкими радиальными ребрами, особенно частыми в верхней части раковины, вблизи киля, а иногда заполняющими пространство между грубыми радиальными ребрами. Имеется примакушечная полость. Отпечаток мускула-замыкателя треугольно-овальной формы, расположен ближе к заднему краю, равноудален от верхнего и нижнего краев. Смычной край узкий, в примакушечной части створки имеются хоматы. Парадонтальная выемка слабо выражена. Замочная площадка экзогироидного типа. Отпечаток мускула Квенштедта на изученном материале не наблюдался. Правая створка маленькая, плоская, треугольно-овальной формы. Макушка маленькая, спирально завернута. На торцевой стороне створок в примакушечной части имеются реликтовые хоматы. Наружная поверхность покрыта концентрическими линиями нарастания, сильнее выраженными по краям створки. Внутреннее строение из-за особенностей сохранности не изучено.
Размеры в мм и отношения
| № экз. | В | Д | Вып | В/Д | Вып/Д |
|---|---|---|---|---|---|
| 2105/395 | 12 | 9 | 9 | 1.33 | 1.00 |
| 2105/405 | 26 | 23 | 14 | 1.13 | 0.61 |
| 2105/418 | 42 | 38 | 21 | 1.11 | 0.55 |
| 2105/419 | 24 | 23 | 14 | 1.04 | 0.61 |
| 2105/421 | 27 | 22 | 15 | 1.23 | 0.68 |
| 2105/423 | 23 | 22 | 14 | 1.05 | 0.64 |
| 2105/425 | 24 | 21 | 13 | 1.14 | 0.62 |
| 2105/427 | 21 | 19 | 12 | 1.11 | 0.63 |
| 2105/431 | 25 | 21 | 13 | 1.19 | 0.62 |
| 2105/434 | 18 | 14 | 9 | 1.29 | 0.64 |
| 2105/438 | 24 | 23 | 13 | 1.04 | 0.57 |
| 2105/440 | 19 | 16 | 9 | 1.19 | 0.56 |
| 2105/444 | 14 | 12 | 8 | 1.17 | 0.67 |
| 2105/459 | 24 | 22 | 11 | 1.09 | 0.50 |
| 2105/590 | 42 | 38 | 20 | 1.11 | 0.53 |
| 2105/591 | 39 | 34 | 22 | 1.15 | 0.65 |
| 2105/592 | 39 | 32 | 17 | 1.22 | 0.53 |
| 2105/593 | 29 | 26 | 14 | 1.12 | 0.54 |
| 2105/594 | 35 | 35 | 18 | 1.00 | 0.51 |
| 2105/595 | 29 | 25 | 10 | 1.16 | 0.40 |
| 2105/608 | 38 | 37 | 12 | 1.03 | 0.32 |
Изменчивость. Очертания раковины изменяются от полулунных до треугольно-овальных, треугольных или вытянутых в высоту овальных. Сильной изменчивости подвержен характер наружной скульптуры левой створки. Варьирует частота и степень развития грубых ребер, а также область развития более тонких ребер. Кроме того, изменчивости подвержена и степень выраженности сулькуса. У большинства экземпляров он отсутствует, в то время как у некоторых он хорошо выражен.
Сравнение. Описываемый вид наиболее близок виду C. pellicoi (Verneuil et Collomb, 1853) (Dhondt, 1982, с. 858, табл. I , фиг. 9–12 ), известному из апта Испании. Обоим видам свойственна значительная изменчивость очертаний раковины и скульптуры, в результате чего найти признаки, позволяющие однозначно разграничить эти виды, не представляется возможным. В настоящее время мы разграничиваем эти виды по известным ареалам их распространения, значительно удаленным друг от друга.
От широко распространенного в сеномане–туроне Тетической надобласти вида C. olisiponensis (Sharpe, 1850) (Malchus, 1990, с. 134, табл. 10 , фиг. 1–6 ) описываемый вид отличается меньшими размерами раковины, отсутствием радиальных ребер на правой створке, более сложной скульптурой левой створки, где, кроме грубых радиальных ребер, присутствуют тонкие радиальные ребра.
Замечания. От Ceratostreon trigonalis (Mirkamalov, 1971) отличается размером раковины, большей выпуклостью левой створки, вогнутостью заднего края раковины, очертанием раковины и характером наружной скульптуры. Помимо этого вида, Миркамаловым (1966) из апт–альбских отложений Гиссара были описаны еще два вида, отнесенные им к роду Exogyra: E. alaeformis Mirkamalov, 1966 и E. luppovi Mirkamalov, 1963. Эти виды различаются между собой очертаниями раковины и характером радиальной ребристости левой створки. Эти различия укладываются в пределы внутрипопуляционной изменчивости. В изученной коллекции имеются переходные экземпляры от одного “вида” к другому по всем этим признакам. Кроме того, описанные Миркамаловым “виды” имеют схожий интервал стратиграфического распространения. Все это послужило основой для сведения описанных Миркамаловым видов в синонимику Costagyra luppovi.
Распространение. Апт–альб юго-западных отрогов Гиссарского хребта, нижний апт Большого Балхана (Западный Туркменистан).
Материал. 1 левая створка из калигрекской свиты зоны Parahoplites melchioris (возвышенность Каттаур), 11 раковин, 39 левых створок из лучакской свиты зоны Hoplites dentatus (7 левых створок из колодца Бегляр; 9 раковин и 15 левых створок из Кундалянгтау; 7 левых створок из колодца Огулбек; 2 раковины и 7 левых створок из Гаурдака; 3 левые створки из Кампрекского ущелья), 1 раковина из лучакской свиты (Кундалянгтау), 2 раковины и 21 левая створка из лучакской свиты зоны Anahoplites rossicus (1 раковина из колодца Огулбек; 2 левые створки из Кампрекского ущелья; 1 раковина и 19 левых створок из ущелья Газдагана).
* * *
Авторы благодарны Т.Н. Богдановой за переданную для изучения коллекцию устриц, а также за полезные советы и помощь в подборе литературы; С.В. Попову за рецензию рукописи статьи и конструктивные советы и замечания.
Работа выполнена при поддержке проекта ФНИ Минобрнауки РФ (№ FWZZ-2022-0004) и является вкладом в проект UNESCO-IUGS IGCP 679.
Список литературы
Вялов О.С. О классификации устриц // Докл. АН СССР. 1936. Т. 4. № 1. С. 19–22.
Вялов О.С. Общая классификация устриц // Докл. АН УССР. Сер. Б. Геол., хим. и биол. науки. 1983. № 11. С. 6–8.
Вялов О.С. Классификация устриц // Палеонтол. сборн. 1984. № 21. С. 31–36.
Косенко И.Н., Метелкин Е.К. Раннемеловые устрицы Мангышлака: таксономический состав, стратиграфическое и палеобиогеографическое распространение // Палеонтол. журн. 2020. № 3. С. 20–31.
Метелкин Е.К., Косенко И.Н. Апт–альбские устрицы юго-западных отрогов Гиссарского хребта. Статья 1: Род Amphidonte Fischer de Waldheim // Палеонтол. журн. 2021. № 6. С. 24–32.
Миркамалов Х.Х. Новые виды экзогир из верхнеальбских отложений юго-западных отрогов Гиссарского хребта // Докл. АН УзССР. 1963. № 3. С. 132–137.
Миркамалов Х.Х. Экзогиры, их систематика и значение для стратиграфии меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: Изд-во ФАН УзССР, 1966. 135 с.
Миркамалов Х.Х. Устрицы окузбулакской свиты (баррем) юго-западных отрогов Гиссарского хребта // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 1971а. Т. 46. № 1. С. 114–123.
Миркамалов Х.Х. Новые виды семейства устричных юго-западных отрогов Гиссарского хребта // Палеонтол. журн. 1971б. № 2. С. 36–40.
Миркамалов Х.Х. Стратиграфия и фауна меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: Изд-во ФАН УзССР, 1986. 104 с.
Мордвилко Т.А. Нижнемеловые отложения Северного Кавказа и Предкавказья. Ч. 2. М.–Л.: Изд-во АН СССР, 1963. 294 с.
Невесская Л.С., Попов С.В., Гончарова И.А. и др. Двустворчатые моллюски России и сопредельных стран в фанерозое. М.: Научн. мир, 2013. 524 с. (Тр. Палеонтол. ин-та РАН. Вып. 294).
Прозоровский В.А. Новые раннемеловые экзогиры Западной Туркмении // Новые виды древних растений и беспозвоночных СССР. Ч. II. М.: Госгеолтехиздат, 1960. С. 73–77.
Aqrabawi M. Oysters (Bivalvia-Pteriomorphia) of the Upper Cretaceous rocks in Jordan. Paleontology, stratigraphy and comparison with the Upper Cretaceous oysters of Northwest Europe // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1993. Bd 75. P. 1–135.
Bayle E. Explication de la Carte Géologique de la France. T. 4. Atlas. Pt 1. Fossiles principaux des terrains, pl. CXLVII–CXXVI. P.: Impr. Royale, 1878.
Carter J.G., Altaba C.R., Anderson L.C. et al. Synoptical classification of the Bivalvia (Mollusca) // Paleontol. Contrib. 2011. № 4. P. 1–47.
Carter J.G., Harries P.J., Malchus N. et al. Illustrated Glossary of the Bivalvia // Treatise online. 2012. № 48. P. 1–209.
Cooper M.R. Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Pt 1 // Durban Mus. Novit. 1995. V. 20. P. 1–48.
Cooper M.R. Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Pt 2 // Durban Mus. Novit. 1997. V. 22. P. 1–31.
Dhondt A.V. Some Spanish Cretaceous bivalves // Cuadern. Geol. Ibérica. 1982. V. 8. P. 847–865.
Freneix S. Disparité microstructurale du test entre Gryphaeostrea et Gyrostrea (huitres du Crétacé–Miocène). Novelle classification du genres // Ann. Paléontol. (Invertébr.). 1982. V. 66. № 3. P. 67–134.
Kues B.S. New bivalve taxa from the Tucumcari Formation (Cretaceous, Albian), New Mexico, and the biostratigraphic significance of the basal Tucumcari Fauna // J. Paleontol. 1997. V. 71. № 5. P. 820–839.
Malchus N. Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik) // Berlin. Geowiss. Abh. Reihe A. 1990. Bd 125. S. 1–231.
Malchus N. Multiple parallel evolution and phylogenetic significance of shell chambers and chomata in the Ostreoidea (Bivalvia) // Bivalves: An Eon of Evolution. Kansas: Univ. Calgary Press, 1998. P. 393–407.
Stenzel H.B. Oysters // Treatise on Invertebrate Paleontology. Pt N. Mollusca 6 (Bivalvia). V. 3 / Ed. R.C. Moore. Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 1971. P. N953–N1124.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал