

Палеонтологический журнал, 2023, № 1, стр. 13-31
Новый род Alexialeks gen. nov. и новые виды радиолярий ранней перми Южного Урала России
М. С. Афанасьева *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: afanasieva@paleo.ru
Поступила в редакцию 04.07.2022
После доработки 19.07.2022
Принята к публикации 19.07.2022
- EDN: FIQTFS
- DOI: 10.31857/S0031031X23010026
Аннотация
Анализ морфологических особенностей скелетов радиолярий из нижнепермских отложений Южного Урала позволил пересмотреть систематическое положение вида Helioentactinia uralica Kozur et Mostler, 1989 и двух разных морфотипов Helioentactinia sp. и отнести их к трем видам в составе нового рода Alexialeks gen. nov.: A. uralicus (Kozur et Mostler, 1989), A. alius sp. nov. и A. distinctus sp. nov. Установлено три направления в развитии Alexialeks gen. nov. на протяжении ассельского, сакмарского, артинского и кунгурского веков ранней перми. Внесены изменения и дополнения в диагнозы рода Multisphaera Nazarov et Afanasieva in Afanasieva, 2000, emend. nov. и подсемейства Multisphaerinae Nazarov et Afanasieva in Afanasieva, 2000, emend. nov. Подтверждена валидность подсемейства Multisphaerinae. Предложена гипотеза о том, что первичная внутренняя сфера с рудиментарной четырехлучевой спикулой и многочисленными стержневидными иглами может представлять собой переходную форму внутреннего каркаса скелета от n-лучевой спикулы к истинной микросфере.
ВВЕДЕНИЕ
Морфотипы скелетов радиолярий с тремя сферами встречаются крайне редко среди радиолярий палеозоя. В настоящее время удалось идентифицировать:
• пять родов радиолярий, обладающих скелетом с тремя пористыми сферами:
(1) Alexialeks gen. nov. (нижняя пермь, Южный Урал, Россия);
(2) Entactinosphaera Foreman, 1963, который является старшим синонимом рода Thecentactinia Nazarov, 1975 (Афанасьева, 2011, с. 5; Noble et al., 2017, с. 448): нижний силур, среднелландоверийский подъярус, Южный Урал, Россия (Назаров, 1975); верхний девон, фаменский ярус, Огайо, США (Foreman, 1963); франский ярус, Северные Мугоджары, Казахстан (Назаров, 1975);
(3) Guangxitrisphaera Wang in Wang et Kuang, 1993, нижний карбон, визейский ярус, Юго-Восточный Гуанси, Южный Китай;
(4) Meschedea Won, 1983, нижний карбон, верхнетурнейский подъярус, Рейнские Сланцевые горы, Германия (Won, 1983);
(5) Wuyia Feng in Feng et al., 2007, верхняя пермь, верхнечансинский подъярус, Южный Гуанси, Южный Китай;
• четыре рода среди радиолярий с губчатым, сетчатым или решетчатым типом скелетной ткани:
(1) Afanasievella Nestell et Nestell, 2010, средняя пермь, верхнекептенский подъярус, Западный Техас, США (Nestell, Nestell, 2010);
(2) Cavasphaera Won et Iams, 2015, нижний ордовик, флоский ярус, Западный Ньюфаундленд, Канада (Won, Iams, 2015);
(3) Glomispina Nestell et Nestell, 2021, средняя пермь, роудский и вордский ярусы, Западный Техас, США (Nestell, Nestell, 2021);
(4) Involutentactinia Jones et Noble, 2006, верхняя часть нижнего силура – нижний венлок, Нунавут, Канада (Jones, Noble, 2006).
Радиолярии с необычной морфологией скелета с тремя сферами впервые были описаны Х. Коцуром и Х. Мостлером (Kozur, Mostler, 1989) на Южном Урале из отложений иренского горизонта кунгурского яруса разреза Алегазово на р. Ай (рис. 1, I). Изначально эти радиолярии были отнесены Коцуром и Мостлером (Kozur, Mostler, 1989) к роду Helioentactinia Nazarov, 1975 и описаны в качестве нового вида H. uralica11 (рис. 2). Кроме этого, два разных морфотипа были приведены Коцуром и Мостлером в открытой номенклатуре как Helioentactinia sp. (рис. 2, г; 3, в, г).
Рис. 1.
Местонахождение радиолярий из рода Alexialeks на Южном Урале: I – р. Ай, с. Алегазово, разрез Алегазово (Kozur, Mostler, 1989); II – р. Юрюзань, с. Мечетлино, разрез Мечетлино (настоящая работа); III – руч. Дальний Тюлькас, пос. Красноусольский, разрез траншеи Дальний Тюлькас (Afanasieva et al., 2022); IV – р. Сакмара, с. Кондуровка, разрез Кондуровский (Afanasieva, 2021); V – р. Урал, с. Донское, разрез Донское (Назаров, 1988; Афанасьева, 2000, настоящая работа).
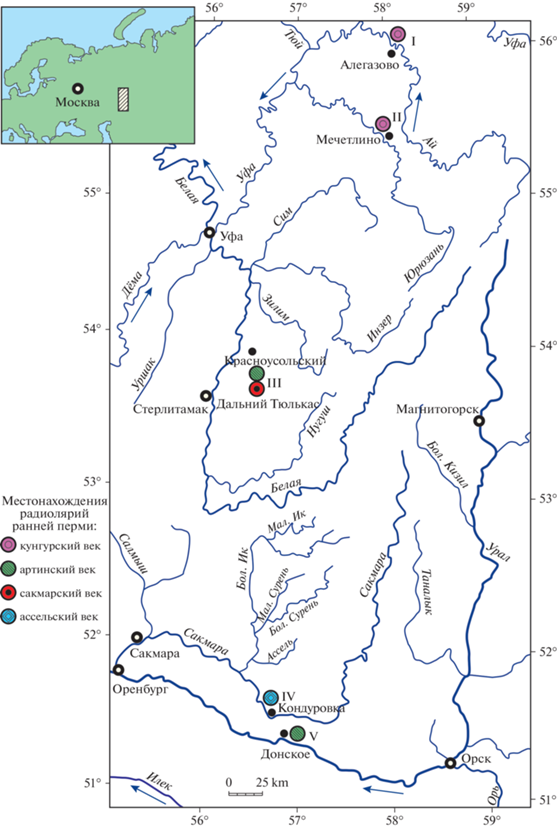
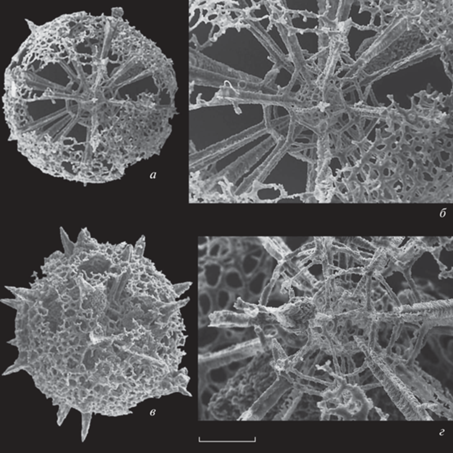
Рис. 2.
Alexialeks uralicus (Kozur et Mostler, 1989): а – экз. № KoMo-1979/I-63b, штрих = 90 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 2); б, в – голотип № KoMo-1979/I-62: б – штрих = 94 мкм (Kozur, Mostler, 1989, табл. 15 , фиг. 7a), в – фрагмент, штрих = 40 мкм (Kozur, Mostler, 1989, табл. 15 , фиг. 7б); г – экз. № KoMo-1979/I-65, штрих = 88 мкм (Kozur, Mostler, 1989, табл. 17 , фиг. 3); д, е – экз. № KoMo-1979/I-63a: д – штрих = 73 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 3a), е – фрагмент, штрих = 22 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 3b); Южный Урал, р. Ай, разрез Алегазово; нижняя пермь, кунгурский ярус, иренский горизонт, зона конодонтов Neostreptognathodus imperfectus.
Новые исследования показали ошибочность рассмотрения вида H. uralica в составе рода Helioentactinia. Морфотип рода Helioentactinia характеризуется наличием только двух сферических оболочек скелета, тогда как у Helioentactinia uralica отмечено формирование скелета из трех сфер, что позволяет уверенно отнести данный вид к новому роду Alexialeks gen. nov.
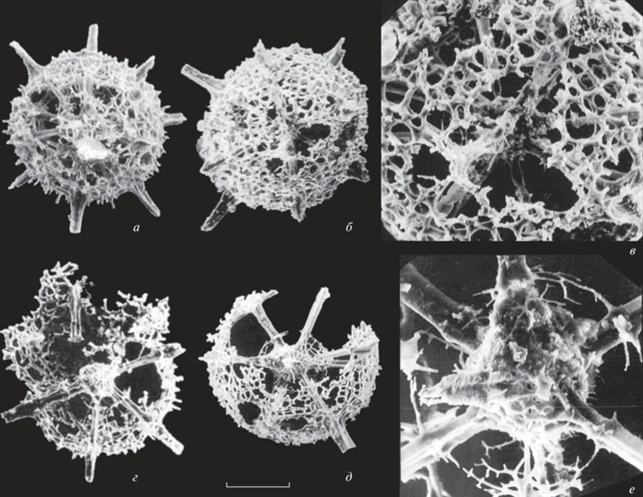
Среди изображенных Коцуром и Мостлером (Kozur, Mostler, 1989) представителей вида Helioentactinia uralica обращают на себя внимание экземпляры с относительно толстой и достаточно тонкопористой внешней оболочкой скелета (рис. 3, а), которая отличается от внешней тонкой и крупнопористой оболочки данного вида (рис. 2). Вместе с тем, подобные морфотипы широко представлены в верхнеассельских отложениях разреза Кондуровский и выделены в самостоятельный новый вид Alexialeks alius sp. nov.
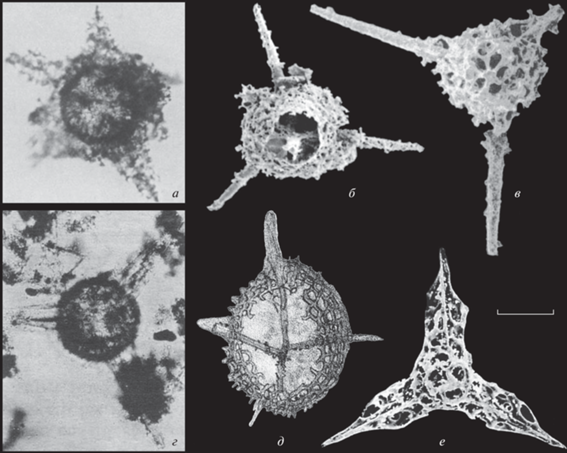
Рис. 3.
Радиолярии Alexialeks alius sp. nov. (а, б) и A. distinctus sp. nov. (в, г): а, б – экз. № KoMo-1979/I-63c: а – экземпляр, на котором видна дополнительная очень тонкая скелетная вуаль (вверху справа), опирающаяся на короткие шипики толстой внешней оболочки скелета, штрих = 108 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 1a), б – фрагмент, штрих = 22 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 1b); в, г – экз. № KoMo-1979/I-64: в – фрагмент, штрих = 14 мкм (Kozur, Mostler, 1989, табл. 17 , фиг. 2), г – штрих = 82 мкм (Kozur, Mostler, 1989, табл. 16 , фиг. 4); Южный Урал, р. Ай, разрез Алегазово; нижняя пермь, кунгурский ярус, иренский горизонт, зона конодонтов Neostreptognathodus imperfectus.
При описании вида Helioentactinia uralica Коцур и Мостлер (Kozur, Mostler, 1989) отметили, что промежуточная оболочка из-за своей хрупкости обычно не сохраняется или сохраняется лишь фрагментарно (рис. 3, г). Однако радиолярии, скелет которых отличается образованием недоразвитой или рудиментарной промежуточной сферы, достаточно часто встречаются в отложениях сакмарского, артинского и кунгурского ярусов Южного Урала. В связи с этим радиолярии с подобным морфотипом скелета выделены ниже в самостоятельный вид Alexialeks distinctus sp. nov.
Таким образом, морфотип радиолярий с тремя пористыми оболочками скелета в настоящей работе описывается в качестве нового рода Alexialeks gen. nov. с тремя видами: A. uralicus (Kozur et Mostler, 1989) и новыми видами A. alius sp. nov. и A. distinctus sp. nov.
ФИЛОГЕНИЯ РОДА ALEXIALEKS
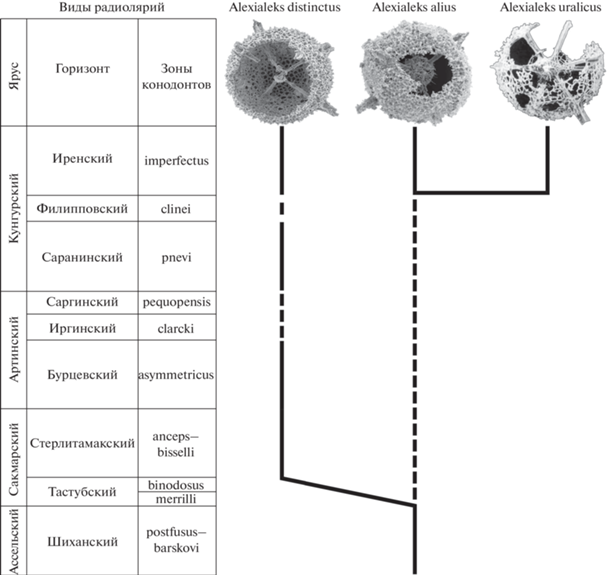
Представители нового рода Alexialeks gen. nov. известны только в нижнепермских отложениях Южного Урала России. Однако в истории этого рода даже на столь коротком временнóм интервале четко выделяются три характерных направления развития (рис. 4).
Рис. 4.
Филогенетическое древо рода Alexialeks gen. nov. Распространение радиолярий в нижнепермских отложениях Южного Урала: A. distinctus sp. nov. – кунгурский ярус: иренский горизонт, р. Ай, разрез Алегазово (Kozur, Mostler, 1989), саранинский горизонт, р. Юрюзань, разрез Мечетлино (настоящая работа); нижнеартинский подъярус, бурцевский горизонт и верхнесакмарский подъярус, стерлитамакский горизонт, пос. Красноусольский, разрез траншеи Дальний Тюлькас (Afanasieva et al., 2022); нижнеартинский подъярус, р. Урал, разрез Донское (настоящая работа); A. alius sp. nov. – кунгурский ярус, иренский горизонт, р. Ай, разрез Алегазово (Kozur, Mostler, 1989) и верхнеассельский подъярус, шиханский горизонт, р. Сакмара, разрез Кондуровский (Afanasieva, 2021); A. uralicus (Kozur et Mostler) – кунгурский ярус, иренский горизонт, р. Ай, разрез Алегазово (Kozur, Mostler, 1989).
Самые ранние представители рода Alexialeks встречены в верхнеассельских отложениях. Они относятся к виду A. alius sp. nov., который представляет основное направление в развитии рода. Данный вид характеризуется крупной толстостенной внешней оболочкой скелета и крайне тонкостенной промежуточной сферой. Последние представители A. alius sp. nov. отмечены в иренских отложениях кунгурского яруса.
В иренское время появились морфотипы с меньшими размерами скелетов, отличающихся формированием тонкостенной внешней и внутренней оболочек. Они образовали самостоятельный вид A. uralicus.
В развитии рода Alexialeks интересно ответвление A. distinctus, которое представлено морфотипами с наиболее крупным скелетом и рудиментарными остатками недоразвитой промежуточной сферы и регулярно встречающихся в отложениях нижней перми. Промежуточная сфера по каким-то причинам не развивалась полностью и утратила свое первоначальное значение; однако ее рудименты занимают примерно такое же положение (в среднем 32% от диаметра внешней сферы), что и развитая промежуточная сфера (в среднем 31–40%). Рудименты промежуточной сферы (от rudimentum лат. – зачаток, первооснова) находились в недоразвитом, зачаточном состоянии в процессе становления и существования нового вида A. distinctus sp. nov. на протяжении сакмарского, артинского и кунгурского веков.
Итак, в процессе развития рода Alexialeks наблюдается:
(1) изменение размера внешней оболочки скелета на 7–10%:
• A. alius – толстостенная большая сфера (Dсредний = 247.5 мкм),
• A. distinctus – наиболее крупный толстостенный скелет (Dсредний = 266 мкм),
• A. uralicus – тонкостенный скелет с меньшими размерами (Dсредний = 221.3 мкм);
(2) увеличение количества игл: от 8 игл у A. alius и A. distinctus, и до 8–12 игл у позднекунгурского вида A. uralicus;
(3) изменение размера промежуточной оболочки скелета:
• A. alius – крайне тонкая сфера, составляющая, в среднем, 30.9% от диаметра внешней сферы,
• A. uralicus – более толстостенная сфера, составляющая, в среднем, 40.4% от диаметра внешней сферы,
• A. distinctus – рудиментарные остатки недоразвитой промежуточной сферы, которая составляет, в среднем, 32% от диаметра внешней сферы и занимает примерно такое же положение, что и развитая промежуточная сфера.
КОНЦЕПЦИЯ КЛАССИФИКАЦИИ РАДИОЛЯРИЙ
В палеонтологической практике определение таксона связано со значительными сомнениями, поскольку практически каждый новый экземпляр привносит новую информацию, которая может изменить сложившийся ранее взгляд на классификацию. В связи с этим решение вопросов систематизации радиолярий невозможно без разработки четких критериев для таксонов различного ранга, поскольку расплывчатость понятий о таксономическом значении морфологических признаков таксонов различного иерархического уровня часто приводит к противоречивым решениям одних и тех же вопросов. Поэтому требуется однозначность трактовки различных морфологических признаков скелетов, не допускающая двусмысленного понимания диагнозов таксонов любого таксономического уровня (Афанасьева, 2000, 2002).
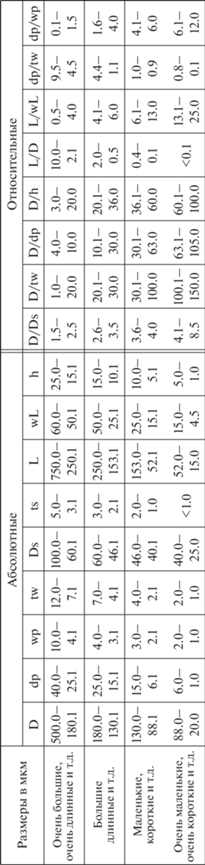
Анализ морфологических признаков скелетов радиолярий палеозоя выявил их устойчивую приуроченность к определенным иерархическим уровням и позволил разработать систему фиксированной совокупности морфологических признаков для диагноза и описания радиолярий палеозоя различного таксономического уровня. Кроме того, морфометрический анализ абсолютных и относительных значений параметров раковин радиолярий палеозоя показал устойчивую статистическую закономерность изменения размеров скелета, его частей и их соотношений. Наличие таких стабильных зависимостей позволяет формализовать процесс классификации данных и оперировать при описании видов понятиями “очень большой”, “большой”, “маленький” и “очень маленький”, которым отвечают определенные ранжированные параметры (рис. 5) (Афанасьева, 2000, 2002).
Рис. 5.
Сводная таблица абсолютных и относительных морфометрических параметров скелетов сферических радиолярий палеозоя (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006; с добавлением и уточнением). Обозначения: D − диаметры внешней (D) и промежуточной (Dm) сфер, dp − диаметры пор на внешней (dp) и промежуточной (dm) сферах, wp − ширина межпоровой перегородки на внешней (wp) и промежуточной (wm) сферах, tw − толщина стенки внешней (tw) и промежуточной (tm) сферы, Ds − диаметр первичной внутренней сферы (микросферы), ts − толщина стенки первичной сферы, L − длина основных игл, wL − ширина основания основных игл, h − высота шипиков на внешней (h), промежуточной (hm) и первичной внутренней (hs) сферах. В скобках указаны обозначения параметров внешней, промежуточной и первичной внутренней сфер, которые используются при описании видов.
Данная концепция классификации радиолярий разрешает учитывать любые сведения об ископаемых организмах и позволяет достаточно просто определить место в филогенетической схеме любого вновь рассматриваемого объекта исследования. Например, подсемейство Multisphaerinae Nazarov et Afanasieva in Afanasieva, 2000 (Афанасьева, 2000), отличающееся развитием трех и более пористых сфер скелета, а также 8–12 и более основных игл, явилось недостающим звеном в построении системы радиолярий палеозоя при выделении в семействе Astroentactiniidae трех подсемейств: Astroentactiniinae − с одной внешней сферой, Helioentactiniinae − с двумя сферами, и Multisphaerinae − с тремя и более оболочками скелета (Афанасьева, 2000, 2002; Afanasieva et al., 2005; Афанасьева, Амон, 2006).
Тем не менее, необходимо помнить известное изречение К. Линнея (1735): “Не признаки определяют род, а род определяет признаки” (цит. по: Петрушевская, 1986, с. 99). Свойства любых биологических существ, современных или вымерших, объективно существуют, т.е., они были, есть и будут вне зависимости от нашей воли или желания. Более того, в природе не существует ни абсолютного тождества, ни абсолютного различия. Следовательно, все наши суждения и выводы о сходстве или различии любых таксонов любого таксономического ранга всегда будут содержать определенную долю субъективизма (Афанасьева, 2000, с. 9).
ПОДСЕМЕЙСТВО MULTISPHAERINAE
Своеобразные морфотипы радиолярий с пятью внутренними сферами были обнаружены Б.Б. Назаровым (1988) в нижнеартинских отложениях разреза Донское (рис. 1, V; рис. 6), но не описаны, а только изображены (Назаров, 1988, табл. IV , фиг. 3, 4; табл. XXVIII , фиг. 6, 7). Более того, своеобразие строения скелетов заставило Назарова сомневаться в отнесении данных радиолярий или к Entactiniinae gen. et sp. indet. (Назаров, 1988, с. 185, табл. IV , фиг. 3, 4), или к Astoentactiniinae gen. et sp. indet. (Назаров, 1988, с. 195, табл. XXVIII , фиг. 6, 7).
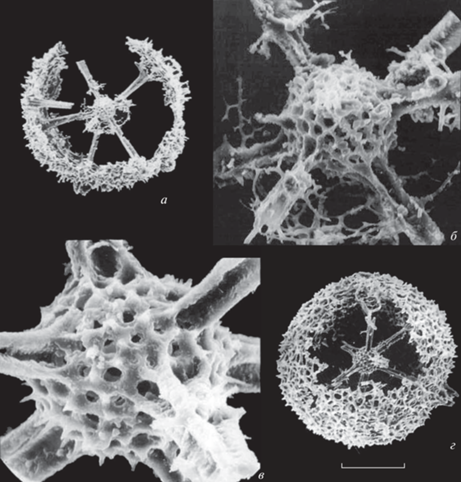
Рис. 6.
Multisphaera impersepta Nazarov et Afanasieva in Afanasieva, 2000, emend. nov.: а – голотип ГИН, № 4673/208-174, штрих = 118 мкм; б – экз. ГИН, № 4673/208-173, штрих = 125 мкм; в–д – экз. ГИН, № 4673/208-61: в – фрагмент, штрих = 33 мкм, г – фрагмент, штрих = 17 мкм; д – фрагмент, штрих = 17 мкм (из: Афанасьева, 2000, табл. 81 , фиг. 7–10 ); е, ж – экз. 3 (адаптировано из: Dumitrica, 2011, табл. 1 , фиг. 4, 5): е – штрих = 96 мкм, ж – фрагмент, штрих = 14 мкм; з, и – экз. 2, фотографии с разной глубиной резкости (адаптировано из: Dumitrica, 2011, табл. 1 , фиг. 1, 2): з – штрих = = 117 мкм, и – штрих = 80 мкм; Россия, Южный Урал, р. Урал, разрез Донское; нижняя пермь, нижнеартинский подъярус.
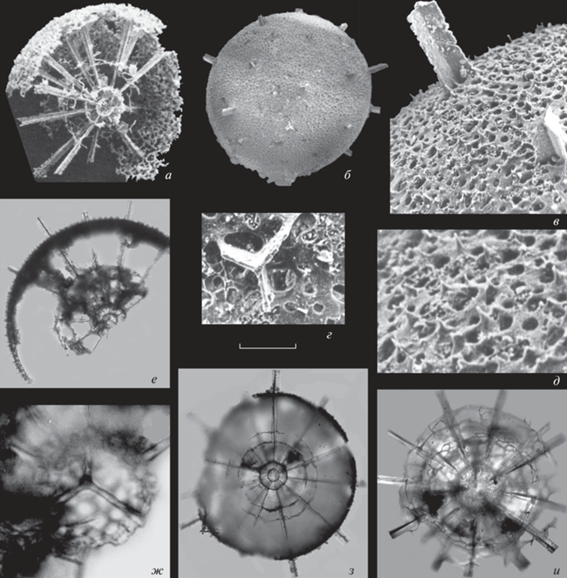
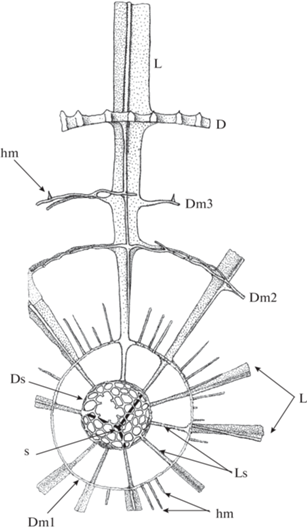
Рис. 7.
Схема строения Multisphaera impersepta Nazarov et Afanasieva in Afanasieva, 2000, emend. nov. (по: Dumitrica, 2011, рис. 1, с изменениями и дополнениями): D − внешняя сфера, Dm1, Dm2 и Dm3 − промежуточные сферы, Ds − первичная внутренняя сфера (микросфера), L − основные трехлопастные иглы, Ls – стержневидные иглы на первичной внутренней сфере (микросфере), hm – шипики на промежуточных сферах, s − четырехлучевая спикула.
Подсемейство Multisphaerinae было установлено на основании описания рода и вида Multisphaera impersepta Nazarov et Afanasieva in Afanasieva, 2000 из нижнеартинских отложений разреза Донское (Афанасьева, 2000). В настоящее время состав подсемейства Multisphaerinae дополнен новым родом Alexialeks gen. nov. из верхнеассельских и нижнесакмарских отложений разреза Кондуровский, верхнесакмарского и нижнеартинского подъярусов разреза траншеи Тюлькас, из базальных кунгурских отложений разреза Мечетлино (саранинский горизонт) и из иренского горизонта разреза Алегазово (рис. 1).
Морфотипы подсемейства Multisphaerinae характеризуются развитием пористых оболочек скелета: трех в скелетах Alexialeks и пяти у представителей Multisphaera.
Внутренний каркас у представителей рода Alexialeks полностью не установлен. Однако внутри первичной внутренней сферы иногда наблюдаются остатки спикулы (табл. II , фиг. 6в; см. вклейку), которые дают возможность предполагать реальность ее первичного присутствия, а число основных игл (от 8 до 12) позволяет сделать вывод о том, что внутренний каркас был представлен гипотетической восьми- или n-лучевой спикулой.
Морфотипы Multisphaera характеризуются развитием очень маленькой (Ds = 27–59 мкм), пористой первичной внутренней сферы (микросферы) с рудиментарной эксцентричной четырехлучевой спикулой и многочисленными стержневидными иглами (рис. 6, а, е–з; 7, Ds, s, Ls).
На дезинтегрированном экземпляре Multisphaera impersepta внутренняя спикула не наблюдается (рис. 6, а). Вместе с тем, на двух экземплярах из препаратов П. Думитрика (Dumitrica, 2011) установил наличие четырехлучевой спикулы, расположенной эксцентрично (рис. 6, е–ж; 7, s). Скорее всего, это рудимент, остаток восьми- или n-лучевой спикулы, утративший свое основное значение в процессе эволюции организма и сохранивший в силу этого эксцентричное расположение. Однако, с другой стороны, первичная внутренняя сфера с рудиментарной четырехлучевой спикулой и многочисленными стержневидными иглами, берущими свое начало на поверхности первичной сферы, может представлять собой переходную форму внутреннего каркаса скелета от n-лучевой спикулы к истинной микросфере.
Лучи спикулы и иглы микросферы связаны с многочисленными основными трехлопастными иглами:
• морфотип Multisphaera насчитывает более 24 игл с  -образным сечением (рис. 6); при этом следует особо подчеркнуть, что все (!) основные иглы Multisphaera берут
свое начало на поверхности первой промежуточной оболочки и являются продолжением стержневидных
игл первичной внутренней сферы, и только часть (!) игл является продолжением внутренней
четырехлучевой спикулы (рис. 7, L, s). Это отчетливо видно на объемном изображении морфотипов (рис. 6, а) и на фотографиях двух экземпляров из препаратов (рис. 6, е, з);
-образным сечением (рис. 6); при этом следует особо подчеркнуть, что все (!) основные иглы Multisphaera берут
свое начало на поверхности первой промежуточной оболочки и являются продолжением стержневидных
игл первичной внутренней сферы, и только часть (!) игл является продолжением внутренней
четырехлучевой спикулы (рис. 7, L, s). Это отчетливо видно на объемном изображении морфотипов (рис. 6, а) и на фотографиях двух экземпляров из препаратов (рис. 6, е, з);
• морфотип Alexialeks представлен 8–12 иглами с  -образным сечением (табл. II, III
; см. вклейку), но у некоторых экземпляров основные иглы имеют субстержневидную форму
между первичной внутренней и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
-образным сечением (табл. II, III
; см. вклейку), но у некоторых экземпляров основные иглы имеют субстержневидную форму
между первичной внутренней и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
Аналогичное внутренне строение скелета наблюдается у рода Glomispina Nestell et Nestell, 2021 (Nestell, Nestell, 2021). Внутренняя спикула или ее остатки обнаружены не были, но на очень маленькой (Ds = 30–40 мкм) пористой внутренней сфере (microsphere: Nestell, Nestell, 2021) берут свое начало многочисленные основные иглы, которые в интервале между первичной внутренней и промежуточной сферами имеют стержневидную форму, которая затем трансформируется в трехлопастную (рис. 8, б, г).
Рис. 8.
Glomispina mirifica Nestell et Nestell, 2021: а, б – голотип № NMMNHS 80661 (Nestell, Nestell, 2021, табл. 2 , фиг. 14a, b ): а – штрих – 88 мкм, б – фрагмент, штрих = 44 мкм; в – экз. № NMMNHS 80659 (Nestell, Nestell, 2021, табл. 2 , фиг. 12 ), штрих – 66 мкм; г – экз. № NMMNHS 80657 (Nestell, Nestell, 2021, табл. 2 , фиг. 10c ), штрих – 28 мкм; США, Техас, Гваделупские горы; средняя пермь (гваделупий), роудский ярус, формация Катофф, пачка Вильямс Ренч.
Все это может служить подтверждением предлагаемой автором гипотезе о переходном статусе внутреннего каркаса скелета от n-лучевой спикулы к истинной микросфере.
СЕМЕЙСТВО TETRENTACTINIIDAE
Думитрика (Dumitrica, 2011) высказал сомнение в валидности подсемейства Multisphaerinae на основании того, что все таксоны с четырехлучевой спикулой должны быть объединены в семейство Tetrentactiniidae Kozur et Mostler, 1979 (=триба Tetrentactiniini Kozur et Mostler, 1979).
Однако, согласно диагнозу Коцура и Мостлера (Kozur, Mostler, 1979), триба Tetrentactiniini объединяет радиолярий с одной решетчатой или губчатой, обычно шаровидной или тетраэдрической оболочкой22. Четыре иглы внутренней спикулы направлены к вершинам тетраэдра и продолжаются снаружи как основные иглы. Часто присутствующие вторичные иглы мало отличаются от основных игл. В состав трибы входят два рода – Tetrentactinia Foreman, 1963 и Tetragregnon Ormiston et Lane, 1976.
Думитрика (Dumitrica, 2011) объединил в семействе Tetrentactiniidae шесть родов из верхнего девона (фамен)–нижнего карбона (турне–визе) только на основании наличия четырехлучевой спикулы (рис. 9): Tetrentactinia, Tetragregnon, Tetraspongoactinia Won, 1998, Staurentactinia Schwartzapfel et Holdsworth, 1996, Ellipsostigma Hinde, 1899 и Triaenosphaera Deflandre, 1973. Кроме этого, Думитрика (Dumitrica, 2011) предложил рассматривать в составе семейства Tetrentactiniidae единственный раннепермский род Multisphaera Nazarov et Afanasieva in Afanasieva, 2000 с четырехлучевой спикулой (рис. 6).
Рис. 9.
Радиолярии семейства Tetrentactiniidae: а – Tetrentactinia barysphaera Foreman, 1963, штрих = 110 мкм (Foreman, 1963, с. 282, табл. 7 , фиг. 9); верхний девон, фаменский ярус, формация Гурон, штат Огайо, США; б – Tetraspongoactinia holdsworthi Won, 1998, штрих = 63 мкм (Won, 1998, с. 257, табл. 4 , фиг. 10 ); нижний карбон, верхнетурнейский подъярус, Рейнские Сланцевые горы, Германия; в – Tetragregnon sycamorensis Ormiston et Lane, 1976, штрих = 134 мкм (Ormiston, Lane, 1976, с. 167, табл. 2 , фиг. 6); нижний карбон, визейский ярус, формация Сикморе, горы Арбакл, штат Оклахома, США; г – Triaenosphaera sicarius Deflandre, 1973, штрих = 63 мкм (Deflandre, 1973, с. 1150, табл. 2 , фиг. 3); нижний карбон, визейский ярус, горы Монтень-Нуар, Франция; д – Ellipsostigma australe Hinde, 1899, штрих = 46 мкм (Hinde, 1899, с. 51, табл. 9 , фиг. 5); средний девон, живетский ярус, формация Яррими, Новый Южный Уэльс, Австралия; е – Staurentactinia nazarovi Schwartzapfel et Holdsworth, 1996, штрих = 65 мкм (Schwartzapfel, Holdsworth, 1996, с. 202, табл. 9 , фиг. 8); верхний девон, верхнефаменский подъярус, формация Вудфорд, Кринер-Хиллз и горы Арбакл, штат Оклахома, США.
При этом в одном семействе Tetrentactiniidae оказались объединены морфотипы с разными типами структуры скелетной ткани (пористые, губчатые, сетчато-решетчатые) и разной геометрической формой скелета (рис. 6, 9): сферические (с одной, двумя и четырьмя оболочками скелета), эллипсоидальные и ставраксонные с четырьмя лучами. Они относятся к трем разным классам радиолярий: пористые Sphaerellaria (Ellipsostigma, Multisphaera, Triaenosphaera), губчато-решетчатые Spumellaria (Tetrentactinia, Tetragregnon, Tetraspongoactinia) и Stauraxonaria с четырьмя лучами-руками (Staurentactinia).
Более того, это смешение разных морфологических признаков таксонов закреплено Думитрикой в диагнозе семейства Tetrentactiniidae:33 “Губчатые или пористые (=latticed)44 Entactinaria палеозоя, имеющие четырехлучевую начальную спикулу с четырьмя лучами, исходящими из центра или от очень короткой срединной балки” (Dumitrica, 2011, с. 212).
Безусловно, спикула является очень важным и определяющим признаком высокого таксономического ранга: отряд, надсемейство, семейство (Афанасьева, 2000, 2002; Afanasieva et al., 2005; Афанасьева, Амон, 2006). Четырехлучевая спикула является универсальным первичным скелетным элементом радиолярий, устойчивым в пространстве и времени.
Скелет радиолярий состоит из опала (SiО2·nH2O), основной структурной единицей которого является кремнекислородный тетраэдр [SiO4]4– – анион ортокремниевой кислоты (Афанасьева, Вишневская, 1992; Точилина, 1997; Афанасьева, 2000, 2007). В вершинах тетраэдра находятся атомы кислорода, а в центре – атом кремния. Особенностью биогенного аморфного кремнезема является способность создавать сложные структуры из коротких трехмерных цепочек тетраэдров [SiO3]2–, которые могут образовывать соединения двух или более анионов через общий атом кислорода.
Данный элемент ультраструктуры первичного опала лежит в основе формирования первичной четырехлучевой спикулы радиолярий, т.е. микроуровень проявляется на макроуровне. При этом внешняя форма скелета отражает общие закономерности молекулярного уровня. Возможно, именно четырехлучевая спикула была прообразом первичного скелета у всех радиолярий (Афанасьева, Вишневская, 1992; Точилина, 1997; Афанасьева, 2000, 2007; Афанасьева, Амон, 2003).
Однако объединение разных морфотипов радиолярий в одном семействе только на основании наличия четырехлучевой спикулы мне кажется неубедительным и опрометчивым. Вместе с тем, наличие четырехлучевой спикулы в морфотипах разных эволюционных ветвей хорошо отражает параллелизм в развитии радиолярий.
ОПИСАНИЕ ТАКСОНОВ
ТИП RADIOLARIA
НАДКЛАСС POLYCYSTINA
КЛАСС SPHAERELLARIA
ОТРЯД ENTACTINIATA
СЕМЕЙСТВО ASTROENTACTINIIDAE NAZAROV ET ORMISTON, 1985
ПОДСЕМЕЙСТВО MULTISPHAERINAE NAZAROV ET AFANASIEVA IN AFANASIEVA, 2000, EMEND. NOV.
Multisphaerinae: Афанасьева, 2000, с. 74; Afanasieva et al., 2005, с. S272; Афанасьева, Амон, 2006, с. 108.
Типовой род – Multisphaera Nazarov et Afanasieva in Afanasieva, 2000.
Диагноз. Astroentactiniidae, скелет которых состоит из трех и более пористых сфер;
внутренний каркас представлен n-лучевой спикулой или микросферой с рудиментарной четырехлучевой
спикулой и многочисленными стержневидными иглами; лучи спикулы и иглы микросферы связаны
с многочисленными (от 8–12 до >24) основными трехлопастными иглами с  -образным или
-образным или  -образным сечением.
-образным сечением.
Состав. Два рода: типовой и Alexialeks gen. nov.; оба из ранней перми России, Южный Урал.
Сравнение. От подсемейства Helioentactiniinae Afanasieva, 1999, которое характеризуется наличием только двух оболочек скелета (пористой внутренней и пористой или сетчатой внешней сферы), а также от семейства Glomispinidae Nestell et Nestell, 2021, характеризующегося наличием трех оболочек скелета (тонкой сетчатой внешней, решетчатой промежуточной с очень большими многоугольными порами и очень маленькой пористой внутренней) подсемейство Multisphaerinae отличается формированием скелета из трех и более пористых сфер.
Замечания. Диагноз подсемейства: (1) изменен в связи с дополнительными данными, полученными Думитрикой (Dumitrica, 2011) при исследовании в препаратах трех экз. радиолярий типового рода из разреза Донское (рис. 1, V; рис. 6, е–и), и на основании особенностей строения скелета нового рода Alexialeks gen. nov.; (2) дополнен новым элементом скелета – микросфера с рудиментарной четырехлучевой спикулой и многочисленными стержневидными иглами.
Состав подсемейства изменен за счет изъятия ошибочно включенного рода Callela Won, 1983 и добавления нового рода Alexialeks gen. nov.
Род Multisphaera Nazarov et Afanasieva in Afanasieva, 2000, emend. nov.
Entactiniinae gen. et sp. indet.: Назаров, 1988, c. 185.
Astoentactiniinae gen. et sp. indet.: Назаров, 1988, c. 195.
Multisphaera: Афанасьева, 2000, с. 75; Dumitrica, 2011, с. 208; Noble et al., 2017, с. 457.
Типовой вид – Multisphaera impersepta Nazarov et Afanasieva in Afanasieva, 2000; Россия, Южный Урал; нижняя пермь, нижнеартинский подъярус.
Диагноз. Раковина сферическая, очень большая (D = 358–390 мкм), с пятью пористыми
оболочками и многочисленными коническими шипиками (рис. 6, д). Первичная внутренняя сфера (микросфера) очень маленькая (Ds = 27–59 мкм), пористая
с многочисленными стержневидными иглами (рис. 6, а, е, з; 7, Ds, Ls). Внутренний каркас представлен рудиментарной четырехлучевой спикулой,
которая расположена эксцентрично в микросфере (рис. 6, е, ж; 7, Ds, s). Многочисленные основные иглы (более 24) имеют трехлопастную форму с  -образным сечением основания иглы и тонкими прямыми лезвиями (рис. 6, а–г, и). Все основные иглы берут свое начало на поверхности первой промежуточной оболочки
и являются продолжением стержневидных игл микросферы (первичной внутренней сферы),
при этом только часть игл является продолжением лучей внутренней спикулы (рис. 6, е, з; 7, L, s).
-образным сечением основания иглы и тонкими прямыми лезвиями (рис. 6, а–г, и). Все основные иглы берут свое начало на поверхности первой промежуточной оболочки
и являются продолжением стержневидных игл микросферы (первичной внутренней сферы),
при этом только часть игл является продолжением лучей внутренней спикулы (рис. 6, е, з; 7, L, s).
Видовой состав. Типовой вид.
Сравнение. См. род Alexialeks gen. nov.
Замечания. Диагноз рода дополнен новым элементом скелета, установленным в связи с дополнительными исследованиями Думитрики (Dumitrica, 2011) радиолярий из типового местонахождения разрез Донское – микросфера с рудиментарной четырехлучевой спикулой и многочисленными стержневидными иглами (рис. 6, е–з).
Длина основных игл за пределами внешней оболочки неизвестна, потому что все они обломаны на небольшом расстоянии от внешней поверхности скелета; но они должны были быть очень длинными, если судить по тому факту, что более-менее сохранившиеся иглы не проявляют тенденции к сужению (рис. 6, б, в, з, и).
Род Alexialeks Afanasieva, gen. nov.
Род назван в честь проф. Александра Сергеевича Алексеева; м.р.
Типовой вид – Alexialeks alius sp. nov.
Диагноз. Раковина сферическая, большая и очень большая (D = 200–295 мкм), с тремя пористыми оболочками. Внешняя оболочка с многочисленными порами разного размера и округло-овального очертания. На узлах межпоровых перегородок расположены конические шипики. На кончиках шипиков развивается дополнительная тончайшая скелетная вуаль толщиной менее 2 мкм, которая сохраняется крайне редко (рис. 3, а; табл. II , фиг. 3в). Промежуточная сфера большая (Dm = 67–95 мкм), тонкостенная и составляет в среднем 31–40% от диаметра внешней сферы. Сфера пронизана многочисленными крупными округло-полигональными порами. На узлах межпоровых перегородок расположены тонкие стержневидные шипики. В некоторых случаях наблюдаются только рудиментарные остатки недоразвитой промежуточной сферы (табл. III , фиг. 2а, 2б, 5а, 5б, 7а, 7б), которая составляет в среднем 32% от диаметра внешней сферы и занимает примерно такое же положение, что и развитая промежуточная сфера. Внутренняя первичная сфера очень маленькая (Ds = 27–59 мкм), очень толстостенная (Ds/ts = 13.3–20.5) и составляет, в среднем, 13–18% от диаметра внешней сферы. Первичная сфера пронизана большими округлыми порами. На узлах межпоровых перегородок расположены длинные тонкие стержневидные шипики.
Основные иглы (числом от 8 до 12) трехлопастные, с  -образным сечением основания иглы. Исключение составляют некоторые представители рода,
которые характеризуются формированием субстержневидной начальной части основных игл
в интервале между внутренней и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
-образным сечением основания иглы. Исключение составляют некоторые представители рода,
которые характеризуются формированием субстержневидной начальной части основных игл
в интервале между внутренней и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
Внутренний каркас полностью не установлен, но внутри первичной сферы иногда наблюдаются остатки спикулы, которые дают возможность предполагать ее реальное присутствие (табл. II , фиг. 6в). Продолжением лучей гипотетической спикулы являются основные иглы, число которых позволяет сделать вывод о том, что внутренний каркас представлен восьми- или n-лучевой спикулой.
Видовой состав. Три вида из нижней перми Южного Урала России: типовой вид из верхнеассельского и иренского горизонта кунгурского яруса; A. distinctus sp. nov. из верхнесакмарского и нижнеартинского подъярусов, саранинского и иренского горизонтов кунгурского яруса; A. uralicus (Kozur et Mostler, 1989) из иренского горизонта кунгурского яруса.
Сравнение. От рода Multisphaera новый род отличается:
(1) формированием скелета из трех сфер, в отличие от скелета Multisphaera с пятью оболочками;
(2) наличием 8–12 основных трехлопастных игл с  -образным сечением основания иглы, тогда как многочисленные (>24) иглы Multisphaera
характеризуются
-образным сечением основания иглы, тогда как многочисленные (>24) иглы Multisphaera
характеризуются 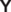 -образным сечением трехлопастных игл (рис. 6, а–г);
-образным сечением трехлопастных игл (рис. 6, а–г);
(3) основные иглы Alexialeks, как правило, имеют трехлопастную форму на всем своем протяжении, за редким исключением (табл. III , фиг. 5б), тогда как основные иглы Multisphaera отличаются трехлопастной формой основных игл в интервале между внутренними оболочками и на поверхности скелета (рис. 6, а–г, е, з, и), при этом у Multisphaera они являются продолжением стержневидных игл микросферы (рис. 6, а, е, з, и);
(4) внутренний каркас Alexialeks представлен гипотетической восьми- или n-лучевой спикулой, занимающей центральное положение в первичной внутренней сфере (табл. II , фиг. 6в), тогда как Multisphaera характеризуется внутренним каркасом в виде микросферы с рудиментарной, эксцентрично расположенной четырехлучевой спикулой и многочисленными стержневидными иглами (рис. 6, а, е–з; 7, Ds, s, Ls).
Замечания. В результате морфологического исследования пересмотрено систематическое положение Helioentactinia uralica и двух разных морфотипов Helioentactinia sp. (Kozur, Mostler, 1989). В настоящей работе они рассматривается в составе нового рода Alexialeks gen. nov. и относятся к виду A. uralicus (Kozur et Mostler, 1989) и двум новым видам – A. alius sp. nov. и A. distinctus sp. nov.
Интересно отметить конвергентное сходство нового раннепермского рода Alexialeks gen. nov. с родом Glomispina Nestell et Nestell, 2021 из роудского и вордского ярусов средней перми (гваделупий) Гваделупских гор штата Техас, США.
На основании наличия трех оболочек скелета Г. Нестелл и М. Нестелл (Nestell, Nestell, 2021) допускали возможность отнесения вида Helioentactinia uralica Kozur et Mostler, 1989 и двух неописанных морфотипов Helioentactinia sp. (Kozur, Mostler, 1989) к роду Glomispina (рис. 8). Однако среднепермские морфотипы Glomispina с сетчатой внешней и решетчатой промежуточной сферами принципиально отличаются от раннепермских морфотипов Alexialeks с тремя пористыми оболочками скелета. Кроме того, основные иглы Glomispina имеют стержневидную форму в интервале между первичной внутренней и промежуточной сферами (рис. 8, б, г), тогда как основные иглы Alexialeks на всем своем протяжении имеют трехлопастную форму, за редким исключением (табл. III , фиг. 5б).
Alexialeks uralicus (Kozur et Mostler, 1989) emend. nov.
Helioentactinia uralica: Kozur, Mostler, 1989, c. 193, табл. 15 , фиг. 7a, b; табл. 16 , фиг. 2, 3a, b.
Helioentactinia sp.: Kozur, Mostler, 1989, с. 249, табл. 17 , фиг. 3.
Голотип – № KoMo-1979/I-62; Россия, Южный Урал, р. Ай, разрез Алегазово; нижняя пермь, кунгурский ярус, иренский горизонт, зона конодонтов Neostreptognathodus imperfectus.
Описание (рис. 2). Раковина сферическая, очень большая (D = 200–250 мкм), состоящая из трех пористых оболочек. Внешняя оболочка тонкостенная (D/tw = 50–70), с многочисленными крупными порами (D/dp = 20.5–25.0). Поры разного размера и округло-овального очертания. Межпоровые перегородки относительно широкие (dp/wp = 2.5–3.3). На узлах межпоровых перегородок расположены многочисленные длинные (D/h = 21.0–33.3) конические шипики.
Промежуточная сфера очень большая (Dm = = 85–95 мкм; D/Dm = 2.4–2.6), тонкостенная (Dm/tm = 31.7–42.5) и составляет 38.4–42.5% от диаметра внешней сферы. Сфера пронизана многочисленными крупными округло-полигональными порами (Dm/dm = 12.1–15.8). Межпоровые перегородки широкие (dm/wm = 1.5–2.7).
Первичная внутренняя сфера очень маленькая (Ds = 37–45 мкм; D/Ds = 4.7–6.1), толстостенная (Ds/ts = 19) и составляет 16.4–21.4% от диаметра внешней сферы. Относительно большие поры внутренней сферы (Ds/ds = 12.7–13.7) расположены довольно равномерно и имеют форму от пятиугольника до шестиугольника. Межпоровые перегородки очень широкие (ds/ws = 2.0). На поверхности сферы расположены отдельные длинные шипики.
Основные иглы (в количестве от 8 до 12) относительно короткие (L = 52–65 мкм; L/D
= 0.2–0.3), трехлопастные, с  -образным сечением основания иглы.
-образным сечением основания иглы.
Внутренняя спикула не установлена. Однако наличие основных игл позволяет сделать вывод о том, что внутренний каркас, вероятно, представлен восьми- или n-лучевой спикулой.
Размеры:
| Абсолютные в мкм | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Промежуточная сфера | Первичная сфера | Основные иглы | |||||||||||
| D | dp | wp | tw | h | Dm | dm | wm | tm | Ds | ds | ws | ts | L | wL | |
| голотип, № KoMo-1979/I-62 |
250 | 10 | 4 | 5 | 9 | 95 | 6 | 4 | 3 | 41 | 3 | – | – | 65 | 29 |
| № KoMo-1979/I-63a | 200 | 8 | 3 | 3 | 6 | 85 | 7 | 3 | 2 | 38 | 3 | 1.5 | 2 | – | 19 |
| № KoMo-1979/I-63b | 225 | 11 | 3 | 4 | 10 | 90 | – | – | – | 37 | – | – | – | 53 | 26 |
| № KoMo-1979/I-65 | 210 | 10 | 3 | 3 | 10 | 85 | – | – | – | 45 | 3 | – | – | 54 | 25 |
| Относительные | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Промежуточная сфера | Первичная сфера | Основные иглы | |||||||||||
| D/dp | dp/wp | dp/tw | D/tw | D/h | D/Dm | Dm/dm | dm/wm | Dm/tm | D/Ds | Ds/ds | ds/ws | Ds/ts | L/D | L/wL | |
| голотип, № KoMo-1979/I-62 |
25.0 | 2.5 | 2.0 | 50.0 | 27.8 | 2.6 | 15.8 | 1.5 | 31.7 | 6.1 | 13.7 | – | – | 0.3 | 2.2 |
| № KoMo-1979/I-63a | 25.0 | 2.7 | 2.7 | 66.7 | 33.3 | 2.4 | 12.1 | 2.7 | 42.5 | 5.3 | 12.7 | 2.0 | 19.0 | – | – |
| № KoMo-1979/I-63b | 20.5 | 3.7 | 2.8 | 56.3 | 22.5 | 2.5 | – | – | – | 6.1 | – | – | – | 0.2 | 2.0 |
| № KoMo-1979/I-65 | 21.0 | 3.3 | 3.3 | 70.0 | 21.0 | 2.5 | – | – | – | 4.7 | 13.0 | – | – | 0.3 | 2.1 |
Обозначения (здесь и далее): внешняя сфера (D − диаметр, dp − диаметр пор, wp − ширина межпоровой перегородки, tw − толщина стенки, h – высота шипиков); промежуточная сфера (Dm − диаметр, dm − диаметр пор, wm − ширина межпоровой перегородки, tm − толщина стенки, hm – высота шипиков); первичная внутренняя сфера (Ds − диаметр, ds − диаметр пор, ws − ширина межпоровой перегородки, ts − толщина стенки, hs – высота шипиков); основные иглы (L − длина, wL − ширина основания).
Сравнение. От нового вида A. alius sp. nov., характеризующегося крупной толстостенной внешней оболочкой скелета и крайне тонкостенной промежуточной сферой, A. uralicus отличается меньшими размерами скелета, формированием тонкостенной внешней и промежуточной оболочек скелета; кроме того, у A. uralicus промежуточная и первичная внутренняя сфера занимают, в среднем, бóльшее внутреннее пространство скелета: 40.4 и 18.3%, соответственно, от диаметра внешней сферы.
Замечания. На основании фотоматериалов, опубликованных Коцуром и Мостлером (Kozur, Mostler, 1989, табл. 15 , фиг. 7a, b; табл. 16 , фиг. 2, 3a, b; табл. 17 , фиг. 3), было осуществлено дополнительное измерение скелетов Helioentactinia uralica и Helioentactinia sp. Выяснить с абсолютной точностью истинное значение измеряемых элементов скелета практически невозможно, однако в рамках погрешности новое измерение морфометрических параметров показало достаточную сопоставимость величин диаметров внешней, промежуточной и внутренней сферы55, следовательно, абсолютные и относительные величины других элементов скелета также можно считать валидными.
Несколько бóльшими оказались размеры промежуточной сферы: 85–95 мкм вместо указанных 79–85 мкм (Kozur, Mostler, 1989), что, вероятно, можно объяснить плохой сохранностью и трудностью идентификации промежуточной сферы, особенно у экз. №№ KoMo-1979/I-62 и KoMo-1979/I-63b (рис. 2, а–в). Однако диаметр внешней оболочки оказался меньше (D = 200–250 мкм), чем было указано ранее (D = 200–294 мкм) (Kozur, Mostler, 1989). Это связано с тем, что экз. № KoMo-1979/I-63c с большим диаметром внешней сферы (D = 294 мкм) рассматривается в составе вида Alexialeks alius sp. nov (рис. 3, а, б).
Материал. Четыре экз. из типового местонахождения (рис. 1, I; рис. 2).
Alexialeks alius Afanasieva, sp. nov.
Табл. II , фиг. 1–6
Helioentactinia uralica: Kozur, Mostler, 1989, c. 193, табл. 16 , фиг. 1a, b.
Helioentactinia sp. 258: Afanasieva, 2021, с. 857, табл. 2 , фиг. 3.
Название вида alius лат. – иной, непохожий.
Голотип – ПИН, № 16455; Россия, Южный Урал, р. Сакмара, разрез Кондуровский; нижняя пермь, верхнеассельский подъярус, шиханский горизонт, зона конодонтов Streptognathodus postfusus–Streptognathodus barskovi.
Описание (рис. 3, а, б). Раковина сферическая, очень большая (D = 212–294 мкм), состоящая из трех пористых оболочек. Внешняя оболочка толстостенная (D/tw = 19.3–28.8) с многочисленными маленькими порами (D/dp = 31.4–48.0). Поры разного размера и округло-овального очертания. Поровые каналы очень узкие (dp/tw = 0.5–0.6), а межпоровые перегородки очень широкие (dp/wp = 1.0–1.5). На узлах межпоровых перегородок расположены многочисленные короткие (D/h = 32.3–58.3) конические шипики. На кончиках шипиков внешней сферы развивается дополнительная очень тонкая скелетная вуаль толщиной менее 2 мкм с очень маленькими порами диаметром 2–3 мкм. Эта вуаль сохраняется крайне редко в связи с формированием тончайшей скелетной ткани (рис. 3, а; табл. II , фиг. 3в).
Промежуточная сфера большая (Dm = 67–81 мкм; D/Dm = 2.9–3.6), крайне тонкостенная (Dm/tm = 101.3–148.0) и составляет 27.8–34.0% от диаметра внешней сферы. Сфера пронизана многочисленными большими округло-полигональными порами (Dm/dm = 11.2–13.7). Межпоровые перегородки очень тонкие (dm/wm = 6–12). На узлах межпоровых перегородок промежуточной сферы расположены многочисленные очень длинные тонкие, стержневидные шипики (hm ≤ 22 мкм; Dm/hm = 3.6–16.8) (табл. II , фиг. 3а, 5).
Первичная внутренняя сфера пористая, очень маленькая (Ds = 27–41 мкм; D/Ds = 6.6–8.6), очень толстостенная (Ds/ts = 13.3–15.5) и составляет 11.6–14.6% от диаметра внешней сферы. Сфера пронизана большими округлыми порами (Ds/ds = 13.5–20.0). Межпоровые перегородки округлые и очень широкие (ds/ws = 0.7–1.5). На узлах межпоровых перегородок внутренней сферы расположены многочисленные очень длинные тонкие стержневидные шипики (hs = 6–8 мкм; Ds/hs = 4.5–5.2) (табл. II , фиг. 4в, 6в). Внутри первичной сферы наблюдаются остатки спикулы, расположение которых дает возможность предполагать центральное положение спикулы в первичной сфере (табл. II , фиг. 6в). Продолжением лучей гипотетической спикулы являются восемь основных игл (табл. II , фиг. 2а, 2б, 4а, 4б). Это позволяет сделать вывод о том, что внутренний каркас, вероятно, представлен восьмилучевой спикулой.
Сохранившиеся основные иглы относительно короткие (L = 53–63 мкм; L/D = 0.22), трехлопастные,
с  -образным сечением основания иглы (табл. II
, фиг. 3б, 5).
-образным сечением основания иглы (табл. II
, фиг. 3б, 5).
Размеры:
| Абсолютные в мкм | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Промежуточная сфера | Первичная сфера | Основные иглы | |||||||||||||
| D | dp | wp | tw | h | Dm | dm | wm | tm | hm | Ds | ds | ws | ts | hs | L | wL | |
| голотип, ПИН, № 16455 |
288 | 6 | 4 | 10 | 8 | 80 | 7 | 0.7 | 0.7 | 22 | 40 | 2 | 3 | 3 | – | 63 | 35 |
| ПИН, № 16492 | 220 | 7 | 5 | 8 | 6 | 74 | 6 | 0.5 | 0.5 | – | 30 | 1.5 | 2 | 2 | 6 | – | 13 |
| ПИН, № 16505 | 212 | 5 | 5 | 11 | 6 | 71 | 6 | 0.5 | 0.5 | – | 31 | 2 | 1.5 | 2 | – | – | 24 |
| ПИН, № 16521 | 233 | 6 | 5 | 12 | 4 | 67 | 6 | 0.5 | 0.5 | 4 | 27 | 2 | 1.7 | 2 | 6 | – | 16 |
| ПИН, № 16342 | 238 | 6 | 4 | 10 | 8 | 81 | 6 | 1.0 | 0.8 | 19 | 30 | – | – | – | – | 53 | 41 |
| № KoMo-1979/I-63c | 294 | – | – | 12 | 9 | 82 | 6 | 1.0 | 0.6 | 10 | 41 | 3 | 2 | 3 | 8 | – | 34 |
| Относительные | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Промежуточная сфера | Первичная сфера | Основные иглы | |||||||||||||
| D/dp | dp/wp | dp/tw | D/tw | D/h | D/Dm | Dm/dm | dm/wm | Dm/tm | Dm/hm | D/Ds | Ds/ds | ds/ws | Ds/ts | Ds/hs | L/D | L/wL | |
| голотип, ПИН, № 16455 |
48.0 | 1.5 | 0.6 | 28.8 | 36.0 | 3.6 | 11.4 | 10 | 114.3 | 3.6 | 6.6 | 20.0 | 0.7 | 13.3 | – | 0.22 | 1.8 |
| ПИН, № 16492 | 31.4 | 1.4 | 0.6 | 27.5 | 36.7 | 3.0 | 12.3 | 12 | 148.0 | – | 7.3 | 20.0 | 0.8 | 15.0 | 5.0 | – | – |
| ПИН, № 16505 | 42.4 | 1.0 | 0.5 | 19.3 | 35.3 | 3.0 | 11.8 | 12 | 142.0 | – | 6.8 | 15.5 | 1.3 | 15.5 | – | – | – |
| ПИН, № 16521 | 38.8 | 1.2 | 0.5 | 19.4 | 58.3 | 3.5 | 11.2 | 12 | 134.0 | 16.8 | 8.6 | 13.5 | 1.2 | 13.5 | 4.5 | – | – |
| ПИН, № 16342 | 39.7 | 1.5 | 0.6 | 23.8 | 29.8 | 2.9 | 13.5 | 6 | 101.3 | 4.3 | 7.9 | – | – | – | – | 0.22 | 1.3 |
| № KoMo-1979/I-63c | – | – | – | 24.5 | 32.7 | 3.6 | 13.7 | 6 | 136.7 | 8.2 | 7.2 | 13.7 | 1.5 | 13.7 | 5.2 | – | – |
Сравнение. От вида A. uralicus, характеризующегося формированием тонкостенной внешней и промежуточной оболочек скелета, новый вид A. alius отличается формированием крупной толстостенной внешней оболочки скелета и крайне тонкостенной промежуточной сферы; кроме того, промежуточная и первичная внутренняя сфера у A. alius занимают, в среднем, на 30% меньшее внутреннее пространство скелета, которое составляет 30.9 и 13.4%, соответственно, от диаметра внешней сферы.
Замечание. Размер диаметра внешней сферы изменяется в позднеассельском веке от 212 до 288 мкм и достигает максимального значения 294 мкм в позднем кунгуре (рис. 3, а).
Материал. 12 экз. из нижнепермских отложений Южного Урала России: 11 экз. из типового местонахождения (рис. 1, IV) и один экз. из кунгурских отложений (иренский горизонт, зона конодонтов Neostreptognathodus imperfectus) отложений разреза Алегазово на р. Ай (Kozur, Mostler, 1989) (рис. 1, I; рис. 3, а, б).
Alexialeks distinctus Afanasieva, sp. nov.
Табл. III , фиг. 1–7
Helioentactinia sp.: Kozur, Mostler, 1989, табл. 16 , фиг. 4, табл. 17 , фиг. 2.
Helioentactinia sp. C: Afanasieva et al., 2022, табл. 3 , фиг. 15 , табл. 5 , фиг. 15 .
Название вида distinctus лат. – отличающийся, отличный.
Голотип – КФУ, № 17719; Россия, Южный Урал, р. Юрюзань, разрез Мечетлино, слой 12; нижняя пермь, кунгурский ярус, саранинский горизонт, зона конодонтов Neostreptognathodus pnevi.
Описание (рис. 3, в, г). Раковина сферическая, большая и очень большая (D = 230–295 мкм), состоящая из двух пористых оболочек и рудиментарных остатков недоразвитой промежуточной сферы (табл. III , фиг. 2а, 2б, 5а, 5б, 7а, 7б). Недоразвитая промежуточная сфера составляет 25.8–38.3% от диаметра внешней сферы.
Внешняя оболочка толстостенная (D/tw = = 21.9–25.6), с многочисленными крупными порами (D/dp = 20.9–25.6). Поры разного размера и округло-овального очертания. Поровые каналы узкие (dp/tw = 0.9–1.2), а межпоровые перегородки широкие (dp/wp = 2.3–3.0). На узлах межпоровых перегородок расположены многочисленные длинные (D/h = 18.8–28.8) конические шипики.
Первичная внутренняя сфера пористая, очень маленькая (Ds = 40–59 мкм; D/Ds = 4.6–6.5), очень толстостенная (Ds/ts = 13.3–20.5) и составляет 14.8–21.7% от диаметра внешней сферы. Сфера пронизана большими округлыми порами (Ds/ds = 10–20). Межпоровые перегородки округлые и очень широкие (ds/ws = 1.0–1.5). На узлах межпоровых перегородок первичной сферы расположены многочисленные очень длинные тонкие стержневидные шипики (hs = 4 мкм; Ds/hs = 10).
Восемь основных игл относительно короткие (L/D = 0.2–0.5), массивные (L/wL = 1.7–4.1),
трехлопастные, с  -образным сечением основания иглы. Однако некоторые морфотипы характеризуются формированием
субстержневидной начальной части основных игл в интервале между первичной внутренней
и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
-образным сечением основания иглы. Однако некоторые морфотипы характеризуются формированием
субстержневидной начальной части основных игл в интервале между первичной внутренней
и рудиментарной промежуточной сферами (табл. III
, фиг. 5б).
Внутренняя спикула не обнаружена. Однако наличие восьми основных игл позволяет сделать вывод о том, что внутренний каркас, вероятно, был представлен восьмилучевой спикулой.
Размеры:
| Абсолютные в мкм | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Первичная внутренняя сфера | Основные иглы | |||||||||
| D | dp | wp | tw | h | Ds | ds | ws | ts | hs | L | wL | |
| голотип, КФУ, № 17719 |
263 | 11 | 4 | 12 | 14 | 41 | 3 | 2 | 2 | – | 80 | 30 |
| КФУ, № 17748 | 295 | 12 | 4 | 12 | 15 | 50 | 3 | 2 | – | – | 151 | 37 |
| КФУ, № 10-1/9-3 | 270 | 12 | 4 | 12 | 11 | 40 | 2 | 2 | 2 | – | 56 | 33 |
| КФУ, № 10-1/9-20 | 272 | 13 | 4 | 11 | 10 | 59 | 3 | 2 | – | – | 98 | 31 |
| № KoMo-1979/I-64 | 230 | 9 | 4 | 9 | 8 | 40 | 4 | 3 | 3 | 4 | – | 30 |
| Относительные | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Экземпляр | Внешняя сфера | Первичная внутренняя сфера | Основные иглы | |||||||||
| D/dp | dp/wp | dp/tw | D/tw | D/h | D/Ds | Ds/ds | ds/ws | Ds/ts | Ds/hs | L/D | L/wL | |
| голотип, КФУ, № 17719 |
23.9 | 2.8 | 0.9 | 21.9 | 18.8 | 6.4 | 13.7 | 1.5 | 20.5 | – | 0.3 | 2.7 |
| КФУ, № 17748 | 24.6 | 3.0 | 1.0 | 24.6 | 19.7 | 5.9 | 16.7 | 1.5 | – | – | 0.5 | 4.1 |
| КФУ, № 10-1/9-3 | 22.5 | 3.0 | 1.0 | 22.5 | 24.6 | 6.8 | 20.0 | 1.0 | 20.0 | – | 0.2 | 1.7 |
| КФУ, № 10-1/9-20 | 20.9 | 2.3 | 1.2 | 24.7 | 27.2 | 4.6 | 19.7 | 1.5 | – | – | 0.4 | 3.2 |
| № KoMo-1979/I-64 | 25.6 | 2.3 | 1.0 | 25.6 | 28.8 | 5.8 | 10.0 | 1.3 | 13.3 | 10.0 | – | – |
Сравнение. От видов A. uralicus и A. alius sp. nov., характеризующихся формированием внутренней оболочки скелета, A. distinctus sp. nov. отличается развитием наиболее крупного скелета (D ≤ 295 мкм) и наличием только рудиментарных остатков недоразвитой промежуточной сферы, которая составляет в среднем 32% от диаметра внешней сферы. Кроме того, отдельные морфотипы A. distinctus характеризуются формированием субстержневидной начальной части основных игл в интервале между первичной внутренней и рудиментарной промежуточной сферами (табл. III , фиг. 5б).
Замечание. Отмечено уменьшение в геологическом масштабе времени диаметра внешней сферы от 270–272 мкм в позднесакмарское–раннеартинское время и 263–295 мкм в начале кунгурского века (саранинское время) до 230 мкм в иренское время (рис. 3, г).
Материал. 21 экз. из нижнепермских отложений Южного Урала, Россия: 13 экз. из типового местонахождения (рис. 1, II); один экз. из кунгурских отложений (иренский горизонт, зона конодонтов Neostreptognathodus imperfectus) разреза Алегазово на р. Ай (рис. 1, I; 3, в, г) (Kozur, Mostler, 1989, табл. 16 , фиг. 4, табл. 17 , фиг. 2); пять экз. из отложений траншеи Дальний Тюлькас близ пос. Красноусольский (рис. 1, III) (Afanasieva et al., 2022): четыре экз. из слоя 10-1 нижнеартинских отложений (бурцевский горизонт, зона конодонтов Sweetognathus asymmetricus), один экз. из слоя 7-2 верхнесакмарских отложений (стерлитамакский горизонт, зона конодонтов Sweetognathus anceps); два экз. из слоя 8 (образец 5/21) нижнеартинских отложений разреза Донское на р. Урал.
* * *
Автор приносит свою искреннюю благодарность А.С. Алексееву, Г.П. Нестелл и В.С. Вишневской за ценные советы и конструктивные рекомендации; В.В. Черныху за консультации относительно возраста вмещающих пород; Г.М. Сунгатуллиной и Р.Х. Сунгатуллину за любезно предоставленный материал по радиоляриям из отложений нижнего кунгура разреза Мечетлино; Э.А. Гайнуллиной за любезно предоставленный материал по радиоляриям из отложений нижнеартинского подъяруса разреза Донское; Б.М. Галиуллину за фотографирование радиолярий на растровом электронном микроскопе; А.Ф. Банникову, М.А. Кнорре и Е.А. Лужной за ценные советы и помощь при подготовке статьи к печати.
Авторские права на фотографии радиолярий, восстановленные из работы Kozur, Mostler, 1989, принадлежат “©Universität Innsbruck, Institut für Geologie” и используются с любезного разрешения Prof. Dr. Diethard Sanders. Авторские права на фотографии радиолярий из работы Nestell, Nestell, 2021, принадлежат “©Micropaleontology” и используются с любезного разрешения редактора Dr. Mike Kaminski.
Коллекция радиолярий ассельского и артинского ярусов нижней перми № 5481 хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН). Коллекция радиолярий кунгурского яруса нижней перми находится в Казанском федеральном ун-те (КФУ).
Исследование морфологии радиолярий проводилось на сканирующих электронных микроскопах CamScan и TESCAN на базе ПИН РАН и на растровом электронном микроскопе XL-30 ESEM на базе КФУ.
Измерения элементов скелетов радиолярий и морфометрический анализ абсолютных и относительных значений параметров раковин радиолярий палеозоя проводились по стандартной методике (рис. 5) (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006).
Работа выполнена в рамках бюджетной программы (государственного задания) ПИН РАН.
Список литературы
Афанасьева М.С. Атлас радиолярий палеозоя Русской платформы. М.: Научный мир, 2000. 480 с.
Афанасьева М.С. Новая классификация радиолярий палеозоя // Палеонтол. журн. 2002. № 2. С. 14–29.
Афанасьева М.С. Скелет радиолярий: морфология игл, внутреннего каркаса и первичной внутренней сферы // Палеонтол. журн. 2007. № 1. С. 3–14.
Афанасьева М.С. Систематическое положение рода Entactinosphaera Foreman, 1963 и новый род Retientactinosphaera gen. nov. (радиолярии палеозоя) // Палеонтол. журн. 2011. № 2. С. 3–13.
Афанасьева М.С., Амон Э.О. Новая классификация радиолярий // Палеонтол. журн. 2003. № 6. С. 72–86.
Афанасьева М.С., Амон Э.О. Радиолярии. М.: ПИН РАН, 2006. 320 с.
Афанасьева М.С., Вишневская В.С. Возможные причины появления кремневого скелета радиолярий // Докл. Акад. наук. 1992. Т. 325. № 3. С. 590–596.
Назаров Б.Б. Радиолярии нижнего–среднего палеозоя Казахстана (методы исследований, систематика, стратиграфическое значение). М.: Наука, 1975. 202 с. (Тр. Геол. ин-та АН СССР. Вып. 275).
Назаров Б.Б. Радиолярии палеозоя. Т. 2. Л.: Недра, 1988. 232 с.
Петрушевская М.Г. Радиоляриевый анализ. Л.: Наука, 1986. 200 с.
Точилина С.В. Проблемы систематики Nassellaria. Биохимические особенности эволюции. Владивосток: ДВО РАН, 1997. 71 с.
Afanasieva M.S. Asselian and Sakmarian (Lower Permian) radiolarian ecozones in the South Urals, Russia // Paleontol. J. 2021. V. 55. № 8. P. 825–862.
Afanasieva M.S., Amon E.O., Agarkov Yu.V., Boltovskoy D.S. Radiolarians in the geological Record // Paleontol. J. 2005. V. 39. Suppl. 3. P. S135–S392.
Afanasieva M.S., Chernykh V.V., Sungatullina G.M et al. Radiolarians, conodonts, and palynomorphs from the Sakmarian–Artinskian boundary beds (Lower Permian) in the Dal’ny Tulkas section, South Urals, Russia // Paleontol. J. 2022. V. 56. № 9. Р. 975–1025.
De Wever P., Dumitrica P., Caulet J.P. et al. Radiolarians in the Sedimentary Record. Amsterdam: Gordon and Breach Sci. Publ., 2001. 533 p.
Deflandre G. Observations et remarques sur les Radiolaires Sphaerellaires du Paléozoïque, à propos d’une nouvelle espece, viséenne, du genre Foremaniella Defl., parfait intermediaire entre les Périaxoplastidiés et les Pylentonémidés // C. R. Acad. Sci. 1973. Sér. D, Sci. natur. V. 276. № 1. P. 1147–1151.
Dumitrica P. On the status of the Permian radiolarian genus Multisphaera Nazarov and Afanasieva, 2000 // Rev. micropaléontol. 2011. V. 54. P. 207–213.
Feng Q.L., Gu S., He W., Jin Y. Latest Permian Entactinaria (Radiolaria) from southern Guangxi, China // J. Micropalaeontol. 2007. V. 26. № 1. P. 19–40.
Foreman H.P. Upper Devonian Radiolaria from the Huron Member of the Ohio Shale // Micropaleontology. 1963. V. 9. № 3. P. 267–304.
Hinde G.J. On the Radiolaria in the Devonian Rocks of New South Wales // Quart. J. Geol. Soc. London. 1899. V. 55. P. 38–64.
Jones M.K., Noble P.J. Sheinwoodian (uppermost Lower Silurian) Radiolaria from the Cape Phillips Formation, Nunavut, Canada // Micropaleontology. 2006. V. 52. P. 289–315.
Kozur H., Mostler H. Beiträge zur Erforschung der mesozoischen Radiolarien. Teil III: Die Oberfamilien Actinommacea Haeckel, 1862 emend., Artiscacea Haeckel, 1882, Multiarcusellacea nov. der Spumellaria und triassische Nassellaria // Geol. Paläontol. Mitt. Innsbruck. 1979. Bd 9. S. 1–132.
Kozur H., Mostler H. Radiolarien und Schwammskleren aus dem Unterperm des Vorurals // Geol. Paläontol. Mitt. Innsbruck. 1989. Sonderband 2. S. 147–275.
Nestell G.P., Nestell M.K. Late Capitanian (latest Guadalupian, Middle Permian) radiolarians from the Apache Mountains, West Texas // Micropaleontology. 2010. V. 56. P. 7–68.
Nestell G.P., Nestell M.K. Roadian (Earliest Guadalupian, Middle Permian) radiolarians from the Guadalupe Mountains, West Texas, USA. Part II: spongy radiolarians (Entactinaria? and Spumellaria) // Micropaleontology. 2021. V. 67. P. 527–555.
Noble P., Aitchison J.C., Danelian T. et al. Taxonomy of Paleozoic radiolarian genera // Catalogue of Paleozoic radiolarian genera / Eds. Danelian T., Caridroit M., Noble P., Aitchison J.C. P., 2017. P. 419–502 (Geodiversitas. V. 39. № 3).
Ormiston A.R., Lane H.R. A unique radiolarian fauna from the Sycamore Limestone (Mississippian) and its biostratigraphic significance // Palaeontogr. Abt. A. 1976. Bd 154. P. 154–180.
Schwartzapfel J.A., Holdsworth B.K. Upper Devonian and Mississippian Radiolarian Zonation and Biostratigraphy of the Woodford, Sycamore, Caney and Goddard Formations, Oklahoma // Cushman Foundation for Foraminiferal Research. 1996. Spec. Publ. № 33. P. 1–275.
Wang Y.J., Kuang G.D. Early Carboniferous radiolarians from Qinzhou, southeastern Guangxi // Acta Micropalaeontol. Sin. 1993. V. 10. № 2. P. 275–287.
Won M.Z. Radiolarien aus dem Unterkarbon des Rheinischen Schiefergebirges (Deutschland) // Palaeontogr. Abt. A. 1983. Bd 182. № 4–6. S. 116–175.
Won M.Z. A Tournaisian (Lower Carboniferous) radiolarian zonation and radiolarians of the A. pseudoparadoxa Zone from Oese (Rheinische Schiefergebirge), Germany // J. Korean Earth Sci. Soc. 1998. V. 19. № 2. P. 216–259.
Won M.Z., Iams W.J. Early/Middle Arenig (Late Floian) Radiolarian Faunal Assemblages from Cow Head Group, Western Newfoundland // Palaeontographica. Abt. A. 2015. Vol. 304. № 1–6. P. 1–63.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал