

Палеонтологический журнал, 2023, № 1, стр. 102-118
Новый представитель рода Pinus L. из юрских отложений Белгородской области, Россия
Н. В. Баженова a, *, А. В. Баженов a, М. В. Теклева a, А. С. Резвый b
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Зоологический музей Зоологического института РАН
199034 Санкт-Петербург, Россия
* E-mail: gordynat@mail.ru
Поступила в редакцию 23.12.2021
После доработки 01.08.2022
Принята к публикации 01.08.2022
- EDN: FIULLA
- DOI: 10.31857/S0031031X23010038
Аннотация
Новый вид ископаемых сосен Pinus bukatkinii sp. nov. обнаружен в среднеюрских отложениях Стойленского карьера (окрестности г. Старый Оскол, Белгородская обл.). Семенные шишки нового вида обладают четырьмя ключевыми диагностическими признаками представителей рода Pinus L.: утолщающиеся в верхней части семенные чешуи, имеющие апофизы с умбо; единый след, васкуляризующий семенную чешую и брактею; смоляные каналы входят в основание семенной чешуи абаксиально; адаксиально выпуклые проводящие пучки семенной чешуи. Отличительной особенностью нового вида является сильная склерификация основной паренхимы семенных чешуй. В шишках P. bukatkinii sp. nov. обнаружены галереи, оставленные фитофагами, что является древнейшим задокументированным случаем питания на семенных шишках сосновых.
ВВЕДЕНИЕ
Число находок семенных шишек анатомической сохранности, отнесенных к роду Pinus, чрезвычайно мало. Один из древнейших представителей этого рода, P. yorkshirensis Ryberg, Stockey, Hilton, Mapes, Riding et Rothwell, происходит из нижнемеловых (готерив–барремских) отложений Йоркшира, Англия (Ryberg et al., 2012), другой, P. belgica Alvin, – из вельда Бельгии (Alvin, 1960). Всего два вида известно из верхнего мела: P. mutoi Saiki из коньяка о-ва Хоккайдо, Япония (Saiki, 1996) и P. driftwoodensis Miller et Malinky из сантона Нью Джерси, США (Miller, Malinky, 1986). Помимо этих четырех видов, отнесенных к современному роду, в меловом периоде существовали таксоны, обнаруживавшие ряд морфолого-анатомических особенностей, характерных для семенных шишек Pinus, но в разной степени отличавшихся от них (Miller, 1976; Falder et al., 1998; Taylor et al., 2009). Такие шишки относят к родам Pityostrobus Dutt, Pseudoaraucaria Fliche и Obirastrobus T. Ohsawa, M. Nishida et H. Nishida (Alvin, 1957; Miller, 1976; Ohsawa et al., 1992). Ряд исследователей возражает против самостоятельности рода Obirastrobus и предлагает рассматривать его представителей в рамках рода Pityostrobus (Smith, Stockey, 2001, 2002). Описанный ниже новый вид является пока лишь вторым юрским представителем семейства Pinaceae, для которого известно анатомическое строение. Первый, Eathiestrobus mackenziei Rothwell, Mapes, Stockey et Hilton из кимериджских отложений Черного о-ва, Северо-Восточная Шотландия (Rothwell et al., 2012), имеет примитивное строение и существенно отличается от представителей рода Pinus.
Появившийся в триасе и широко распространенный в юре – раннем мелу род Sсhizolepidopsis Doweld, по мнению ряда исследователей, также должен быть отнесен к сосновым (Harris, 1979; Leslie et al., 2013; Matsunaga et al., 2021 и др.). Данные анатомии пока что известны только для одного раннемелового представителя данного рода (Matsunaga et al., 2021), отношение которого к более древним представителям не вполне ясно.
Семенные шишки Pinus bukatkinii sp. nov. являются самой древней находкой, демонстрирующей ключевые анатомические признаки, характерные для шишек рода Pinus, согласно Ч. Миллеру (Miller, 1976), а также самой древней находкой репродуктивных органов представителя семейства сосновых анатомической сохранности.
Авторы благодарны М.С. Рогову из Геологического ин-та РАН, Москва (ГИН РАН) за обсуждение вопросов геологии Стойленского местонахождения, и Р.А. Ракитову из Палеонтологического ин-та им. А.А. Борисяка РАН, Москва (ПИН РАН) за помощь при работе с СЭМ и компьютерным рентгеновским томографом. Работа поддержана грантом РФФИ № 19-04-00046.
МАТЕРИАЛ И МЕТОДИКА
Изученный материал происходит из Стойленского карьера, Белгородская обл. (окрестности г. Старый Оскол), где он был собран В.В. Букаткиным (Стойленский горно-обогатительный комбинат) в 2001–2002 гг. из небольшой песчанистой линзы с растительным детритом, датируемой средней юрой, предположительно, батом (Баженов, Баженова, 2022). Фрагменты семенных шишек сохранились преимущественно в виде бурого угля, но некоторые из них подверглись частичной пиритизации; у нескольких экземпляров ткани частично замещены белым глинистым матриксом. Часть углефицированных шишек имеют превосходную анатомическую сохранность.
Поверхность обнаруженных шишек была покрыта эпоксидным клеем для предупреждения разрушения. При смачивании углистое вещество шишек начинает разрушаться, что не позволило изготовить тонкие срезы. Особенности морфологии образцов были изучены при помощи стереомикроскопа Leica M165C с цифровой камерой Leica DFC 420C. Анатомическое строение изучалось на поперечных и продольных сколах в СЭМ Tescan, а также с использованием компьютерных рентгеновских томографов SkyScan 1172 и Neoscan 80. Крупные углефицированные фрагменты плохо поддаются золочению и разрушаются при использовании режима высокого вакуума, поэтому преимущественно использовался режим низкого вакуума (BSE детектор, 20 кВ), без напыления. В режиме высокого вакуума (SE детектор, 30 кВ), с напылением золотом и палладием изучались фрагменты шишечных чешуй и семена. Препараты оболочек семян изготавливались по стандартной методике, путем последовательного погружения в плавиковую и азотную кислоту, а затем в раствор КОН. Полученные препараты изучались в СЭМ Тескан и под световым микроскопом Axioplan-2. Пыльцевые зерна были изучены в СЭМ Тескан в режиме высокого вакуума (SE детектор, 30 кВ).
Коллекция № 5851 хранится в ПИН РАН, Москва.
ОПИСАНИЕ
ПОРЯДОК PINALES
СЕМЕЙСТВО PINACEAE LINDLEY, 1836
Род Pinus L., 1753
Pinus bukatkinii Bazhenova et Bazhenov, sp. nov.
Табл. IX , фиг. 1–16 ; табл. X , фиг. 1–7 (см. вклейку)
Рис. 1.
Pinus bukatkinii sp. nov.: а – паратип ПИН РАН, № 5851/8, продольная томограмма фрагмента семенной шишки; б, ж – паратип ПИН РАН, № 5851/4: б – продольная томограмма основания семенной шишки; ж – вертикальные смоляные каналы в ксилеме шишечной оси, образующие два кольца – полное на границе с сердцевиной (отмечено белыми стрелками) и дополнительное неполное (отмечено серыми стрелками), СЭМ; в–е – паратип ПИН РАН, № 5851/2, СЭМ: в – поперечный скол семенной шишки, вертикальные смоляные каналы в коре шишечной оси, расширенные в месте ветвления, отмечены стрелками; г – деталь фиг. 1, в, видны нормальные вертикальные смоляные каналы в коре оси шишки и расширенный в месте ветвления канал (отмечен белой стрелкой); д – деталь фиг. 1, в, сердцевина, окруженная кольцом ксилемы; е – вертикальные смоляные каналы в ксилеме шишечной оси, расположенные у границы с сердцевиной (отмечены белыми стрелками). Обозначения: bt – след брактеи, с – кора, g – галерея, p – сердцевина, ph – флоэма, ssc – семенная чешуя, tr – единый след брактеи и семенной чешуи, vrc – вертикальные смоляные каналы, х – ксилема.
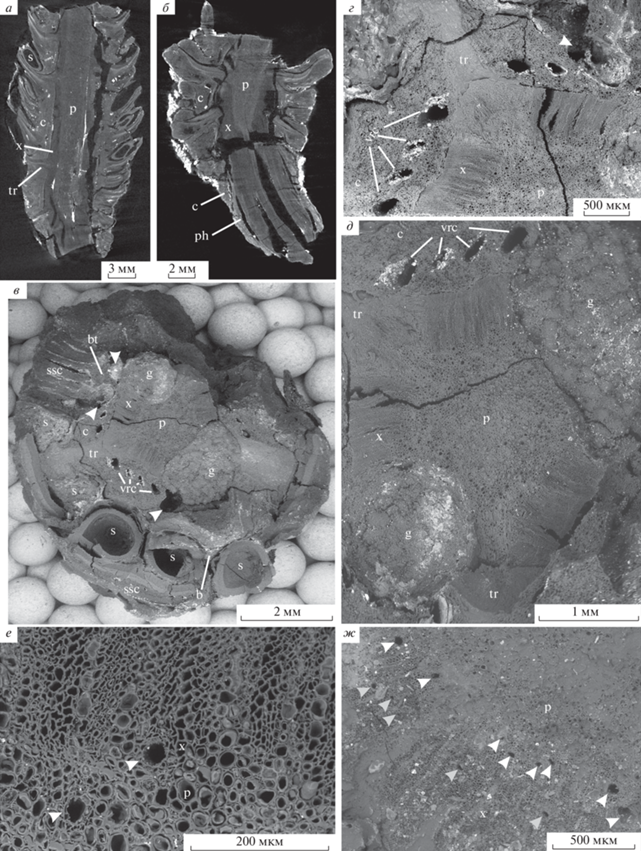
Рис. 2.
Pinus bukatkinii sp. nov., СЭМ: а – паратип ПИН РАН, № 5851/2, поперечный скол семенной чешуи на уровне семенных лунок, виден единственный уплощенный проводящий пучок и ряд смоляных каналов под ним (отмечены белыми стрелками); б, е – голотип ПИН РАН, № 5851/9: б – поперечный скол семенной чешуи в средней части, видны многочисленные проводящие пучки и смоляные каналы (отмечены белыми стрелками), утратившие однорядное расположение; е – поперечный скол семенной чешуи под апофизом, проводящие пучки (отмечены белыми стрелками) начали терять однорядное расположение, видны абаксиальные и адаксиальные смоляные каналы; в–д – паратип ПИН РАН, № 5851/1: в – фрагмент поперечного скола семенной чешуи в средней части, видны тонкая абаксиальная и адаксиальная гиподерма, небольшие гнезда склереид в абаксиальной и адаксиальной коре, а также в основной паренхиме; г – фрагмент скола семенной чешуи за семенными лунками, видны склерификация основной ткани и смоляные каналы (отмечены белыми стрелками), перемещающиеся в адаксиальное положение; д – проводящий пучок и склерифицированные ткани вокруг него. Обозначения: ir – межсеменной гребень, scl – склерификация, vb – проводящий пучок.
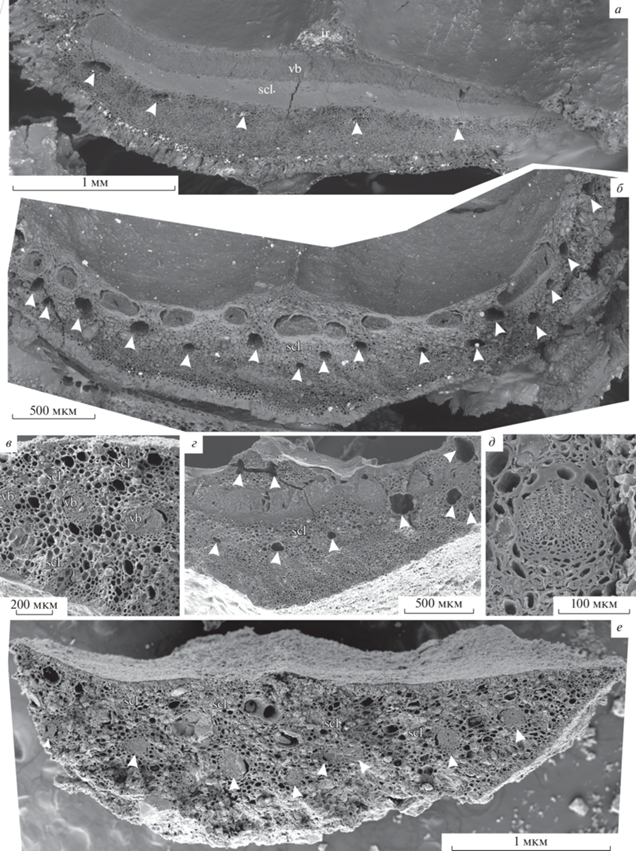
Рис. 3.
Pinus bukatkinii sp. nov., рентгеновские томограммы: а, е – паратип ПИН РАН, № 5851/2: а – поперечное сечение, демонстрирующее основные детали анатомии семенной шишки и две крупные галереи; е – радиальное сечение, демонстрирующее отделение следа брактеи от общего следа; б–г – голотип ПИН РАН, № 5851/9: б – тангентальное сечение, наблюдается отделение следа брактеи от общего следа и ветвление пары смоляных каналов (верхние ответвления входят в семенную чешую, нижние – в брактею); в – поперечное сечение, демонстрирующее основные детали анатомии семенной шишки, а также галереи различного диаметра; г – тангентальное сечение, видна брактея, отделяющаяся от семенной чешуи; д – паратип ПИН РАН, № 5851/8, тангентальное сечение, видны единые следы семенной чешуи и брактеи; ж – паратип ПИН РАН, № 5851/4, тангентальное сечение, видны стерильные чешуи в основании шишки с отделяющимися от них брактеями. Обозначения: b – брактея, bb – проводящий пучок брактеи, brc – смоляные каналы брактеи, bt – след брактеи, g – галерея, ir – межсеменной гребень, ph – флоэма, s – семя, sc – склерификация, ssc – семенная чешуя, sscrc – смоляные каналы семенной чешуи, ssct – след семенной чешуи, tr – единый след брактеи и семенной чешуи, vrc – вертикальные смоляные каналы, w – крыло.
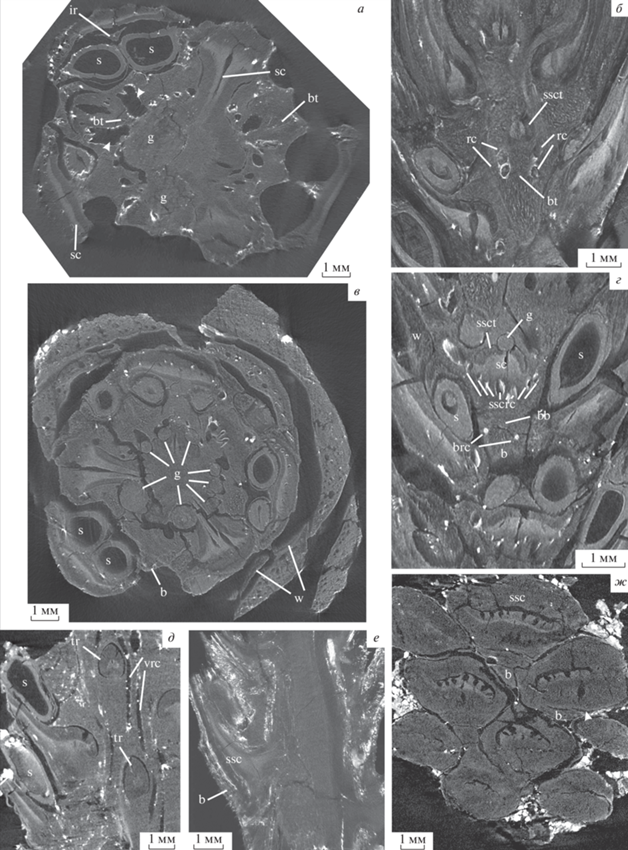
Рис. 4.
Pinus bukatkinii sp. nov.: а–в – голотип ПИН РАН, № 5851/9, СЭМ: а – семя с удаленной саркотестой, видна ребристость поверхности склеротесты; б – два крылатых семени, вид с дорсальной стороны; в – семя, вид с вентральной стороны; г–е, з–к – паратип ПИН РАН, № 5851/5: г – нуцеллус с мегаспоровой мембраной, СМ; д – нуцеллус с мегаспоровой мембраной, СЭМ; е – деталь фиг. 1, д, верхушка нуцеллуса с микропиле; з – деталь фиг. 1, е, микропиле с двумешковым пыльцевым зерном; и – кутикула нуцеллуса; к – мегаспоровая мембрана на поперечном сколе; ж – паратип ПИН РАН, № 5851/2, поперечный скол семенной чешуи с семенами, СЭМ. Обозначения: m – микропиле, po – пыльцевое зерно, s – семя, ssc – семенная чешуя.
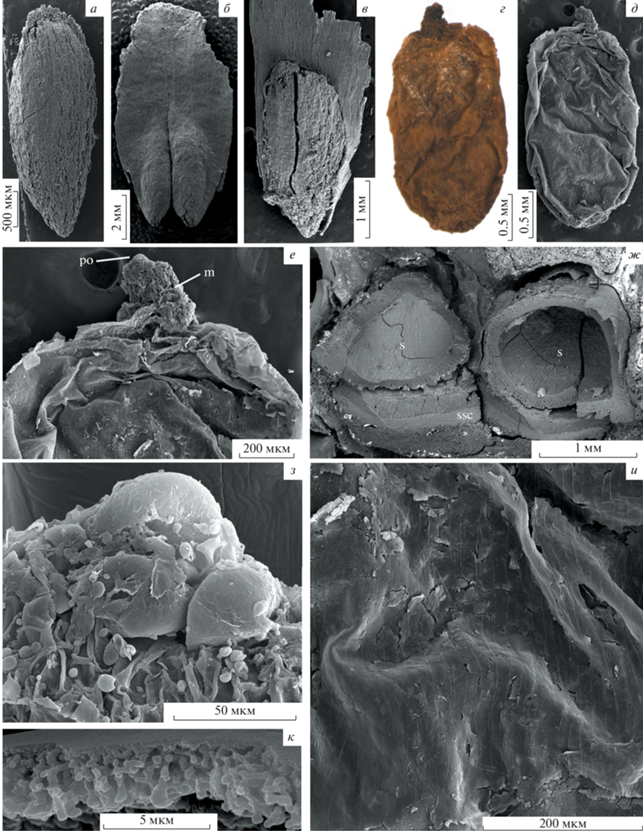
Рис. 5.
Pinus bukatkinii sp. nov.: а–и – пыльцевые зерна с поверхностей внутри голотипа ПИН РАН, № 5851/9, СЭМ: а–ж – пыльцевые зерна, обнаруженные на поверхности семян; з, и – пыльцевые зерна, обнаруженные на адаксиальной поверхности семенной чешуи; а–г, ж, з – пыльцевые зерна с проксимальной стороны, на фиг. 5, ж видна внутренняя структура (endoreticulations) воздушного мешка в разрыве; д – пыльцевое зерно в боковом положении; е, и – пыльцевые зерна с дистальной стороны, видна лептома; масштабная линейка 20 мкм; к, л – паратип ПИН РАН, № 5851/4, СЭМ: к – ксилемный луч на радиальном сколе ксилемы ножки семенной шишки; л – деталь фиг. 5, к, видны поля перекреста с пицеоидными порами; м – голотип ПИН РАН, № 5851/9, на продольной томограмме видна крупная галерея (обведена пунктирной линией) с ответвлениями, разворачивающаяся у верхушки шишки.
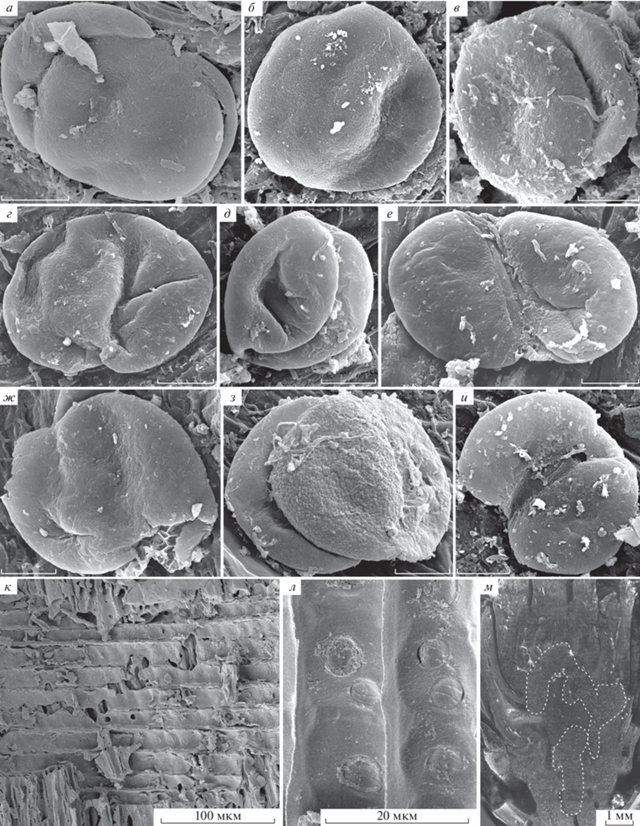
Рис. 6.
Pinus bukatkinii sp. nov.: а – голотип ПИН РАН, № 5851/9, СЭМ, краевая часть семенной чешуи, видны перпендикулярно ориентированные крупные клетки и слабая склерификация абаксиальной гиподермы, а также несклерифицированная основная ткань, смоляные каналы и окружающие их тонкостенные клетки; б, е, ж – паратип ПИН РАН, № 5851/2, СЭМ: б – след брактеи и расширенные вертикальные смоляные каналы в коре шишечной оси; е – след брактеи, только что отделившийся от единого следа и направляющийся вниз, также видны смоляные каналы, идущие в семенную чешую; ж – деталь фиг. 6, е, эллипсоидальный проводящий пучок следа брактеи; в, з–к – паратип ПИН РАН, № 5851/4: в – поперечная томограмма ножки семенной шишки, видны сердцевина, кольцо ксилемы, флоэма и кора; з – клетки сердцевины ножки семенной шишки на продольном сколе, СЭМ; и – флоэма ножки семенной шишки на поперечном сколе, видны массивы клеток флоэмы, чередующиеся с участками, содержащими крупные идиобласты, СЭМ; к – смоляной ход в коре ножки семенной шишки; г, д – паратип ПИН РАН, № 5851/1, СЭМ: г – смоляной канал на продольном сколе семенной чешуи; д – двурядно расположенные проводящие пучки на поперечном сколе основания апофиза (отмечены белыми стрелками). Обозначения: bt – след брактеи, с – кора, p – сердцевина, ph – флоэма, rc – смоляной канал, sc – склерификация, ssc – семенная чешуя, tr – единый след брактеи и семенной чешуи, vb – проводящий пучок, vrc – вертикальные смоляные каналы, х – ксилема.
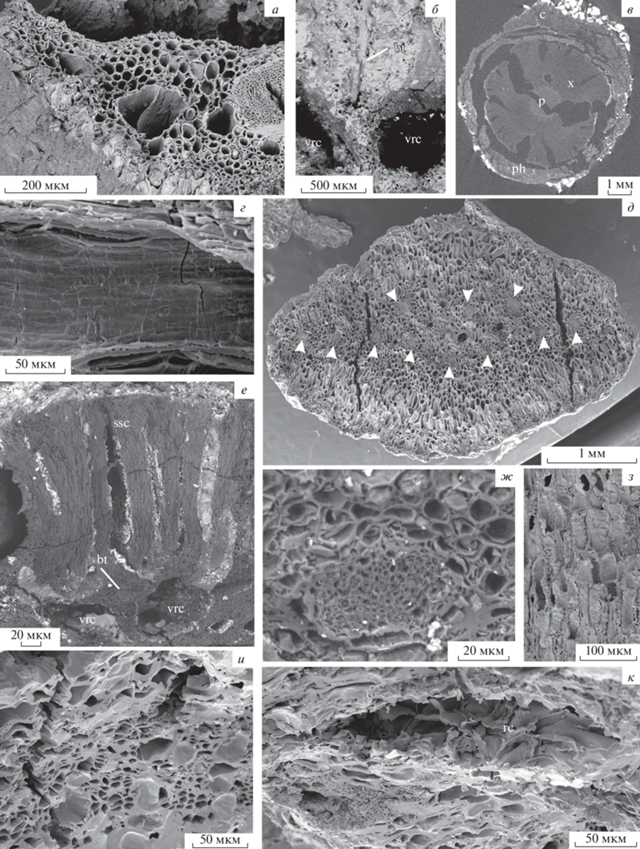
Рис. 7.
Pinus bukatkinii sp. nov.: а – отхождение единого следа брактеи и семенной чешуи; б – смоляные каналы комплекса семенной чешуи и брактеи и их связь с вертикальными смоляными каналами коры шишечной оси (показаны смоляные каналы только с одной стороны следа, идущего к брактее и семенной чешуе).
Вид назван в честь В.В. Букаткина, основателя музея “Художественно-палеонтологический центр” при Стойленском ГОКе.
Голотип – ПИН РАН, № 5851/9, лигнитизированный фрагмент шишки; Европейская часть России, Белгородская обл., окрестности г. Старый Оскол, Стойленский карьер; средняя юра, бат?; обозначен здесь, табл. IX , фиг. 3, 4.
Diagnosis. Incomplete cones oblong cylindrical, 19–33 mm in diameter when closed, more than 55 mm long; cone base broadly cuneate, symmetrical, with massive peduncle more than 10 mm long, 5–7.3 mm in diameter. Bract-scale complexes helically arranged. Seed scales oblong to obovoid-oblong, up to 17 mm long, 7–10 mm wide; interseminal ridge well-developed. Apophyses widely rhomboidal to pentagonal with distinctly rounded upper margin, 5–8 mm long, 7–11.4 mm wide, transversely keeled, bearing dorsal transversely oval umbo. Apophyses in lower third of seed cone distinctly raised (up to 3–5 mm high) with protruding slightly recurved umbo; apophysis and umbo less prominent toward apex. Pith 2–2.7 mm in diameter. Secondary xylem forming complete cylinder up to 1.10 mm thick without grows rings; resin canals near pith area, occasionally forming additional incomplete ring in middle part of xylem. Cortex about 1–2 mm thick, parenchymatose; inner zone with 16–20 vertical resin canals dilated 2–6 times at places of branching. Trace of seed scale and bract entire, with sclerified pith. Bract trace detaching from bract-scale complex trace downward then recurving to supply bract. Bract up to 2.5 mm long, parenchymatose. Seed scale vascular bundle branching in middle part of scale to form 12–15 vascular bundles. Vascular bundles initially arranged in one row, then in two rows in apical part of scale. Resin canals abaxial near seed scale base, then abaxial, adaxial, and interfascicular. Ground tissue below vascular bundles strongly sclerified in the proximal half of seed scale. Abaxial hypodermal layer thin, uneven, consisting of one–three rows of small sclerenchyma cells. Adaxial cortex in proximal part of seed scale non-sclerified; in its middle part, containing sclerenchyma nests and thin hypodermal layer of one–three cells; in distal part, non-sclerified. Seeds two per scale, inverted, winged, narrowly ellipsoidal, 2.5–4 mm long, 1.2–1.6 mm wide; wing semi-oval, thickened near seed body; seed including wing up to 9 mm long.
Описание (рис. 1–7). В коллекции имеются три фрагмента оснований шишек длиной 22 мм (№ 5851/4), 29 мм (№ 5851/8) и 44 мм (№ 5851/6), шириной 20, 15 и 33 мм соответственно, а также фрагменты средних частей шишек длиной 33–47 мм, шириной 27–32 мм, и один крупный фрагмент с сохранившейся верхушкой, но без основания (табл. IX , фиг. 1–16 ; рис. 1, а), длиной 55 мм, шириной 19 мм. Поскольку некоторые экземпляры слегка уплощены, их ширина несколько больше реального диаметра, который не превышал 30 мм. Представлены фрагменты шишек, как в закрытом виде, так и частично раскрывшиеся. Исходя из размеров и пропорций шишек, они были удлиненно-цилиндрическими, более 55 мм длиной, 16–30 мм в диаметре. Основания шишек широко-клиновидные (табл. IX , фиг. 1, 2, 9, 10 ), шишечные чешуи расположены спирально.
Ножка (хорошо сохранилась только у одного экземпляра) прямая, массивная, длиной более 10 мм, диаметром 5–7.3 мм (табл. IX , фиг. 9, 10 ; рис. 1, б; 6, в). Сердцевина ножки диаметром около 1200 мкм, сложена толстостенными паренхиматозными клетками диаметром 8–40 мкм, длиной 40–120 мкм, часто заполненными органическим содержимым (табл. X , фиг. 1, 4; рис. 6, в, з). Протоксилема эндархная (табл. X , фиг. 4). Вторичная ксилема образует сплошное кольцо толщиной около 1700 мкм; сложена трахеидами диаметром до 20 мкм. Поровость радиальных стенок трахеид однорядная, поры чаще соприкасаются, но иногда расстояние между соседними порами составляет 2–6 мкм; на концах трахеид могут встречаться поры с очередным расположением (табл. X , фиг. 6, 7). Поры округлые, диаметром 9–11 мкм, ширина окаймления 3–6 мкм, апертура округлая, диаметром 2.5–3 мкм. Поровость тангентальных стенок однорядная. Ксилемные лучи высотой 1–11 клеток, преимущественно однорядные, но встречаются двурядные участки (табл. X , фиг. 2, 5; рис. 5, к, л). Клетки лучей длиной 15–120 мкм, высотой 15–20 мкм, с относительно тонкими ровными стенками (рис. 5, л). Горизонтальные стенки лучей с простыми порами диаметром около 2 мкм, тангентальные стенки гладкие, без пор (табл. X , фиг. 3). Поля перекреста с одной–тремя пицеоидными или купрессоидными порами диаметром 4–5 мкм (табл. X , фиг. 5; рис 5, л). Лучевые трахеиды не обнаружены. Вертикальные нетравматические смоляные каналы диаметром до 180 мкм с крупными тонкостенными эпителиальными клетками рассредоточены по всей ширине кольца ксилемы, редкие (табл. X , фиг. 1, 4). Флоэма шириной около 150 мкм (рис. 6, в, и). Клетки флоэмы диаметром 4–10 мкм, выстроены нечеткими радиальными рядами внутри небольших массивов, разделенных радиальными рядами (около пяти в одном ряду) крупных идиобластов диаметром до 40 мкм. Кора шириной до 1250 мкм, сохранилась частично, сложена паренхиматозными клетками диаметром 30–50 мкм; смоляные каналы диаметром около 100 мкм, выстланы тонкостенными эпителиальными клетками (рис. 6, в, к).
Шишечная ось в средней части диаметром около 6.5 мм, с хорошо выраженной сердцевиной диаметром 2–2.7 мм, сложенной толстостенными паренхиматозными клетками диаметром 5–60 мкм и окруженной кольцом ксилемы толщиной 0.6–1.10 мм (рис. 1, а–д; 3, а, в, е). Вторичная древесина шишечной оси образует сплошной цилиндр. Трахеиды в поперечном сечении от округлых до округло-многоугольных, диаметром 5–20 мкм (рис. 1, а). Ксилемные лучи однорядные. Клетки лучей длиной до 20 мкм, шириной 40 мкм. В основании прироста видны вертикальные нетравматические смоляные каналы, диаметром до 95 мкм (в среднем 20 мкм) (рис. 1, е, ж). У двух экземпляров есть дополнительное кольцо смоляных каналов ближе к средней части кольца ксилемы (рис. 1, ж). Годичные кольца не выражены, но не исключено, что дополнительное кольцо смоляных каналов маркирует границу между двумя слоями прироста. Флоэма не сохранилась. Кора толщиной 1–2 мм, сложена преимущественно толстостенными паренхиматозными клетками диаметром 10–50 мкм, однородная. Только тонкая внешняя часть коры (до четырех клеток толщиной) сложена относительно более мелкими клетками диаметром до 25 мкм (рис. 1, г). Во внутренней зоне коры имеется кольцо из 16–20 вертикальных смоляных каналов диаметром 90–300 мкм, которые могут значительно расширяться в месте ветвления (до 800 мкм).
Проводящие пучки, васкуляризующие шишечные чешуи, отходят от оси под более или менее прямым углом и сопровождаются двумя смоляными каналами, ответвляющимися от пары вертикальных смоляных каналов во внутренней коре (рис. 1, а–г; 3, а–в, е; 7, а, б). Проводящие пучки эндархные, со склерифицированной сердцевиной, сначала округлые, диаметром 0.7–1.2 мм (рис. 1, д), но вскоре распадаются на адаксиальный аркообразный пучок, следующий к семенной чешуе, и небольшой абаксиальный округлый пучок, васкуляризующий брактею (рис. 1, б, г; 3, е; 6, б, е, ж). Пучок, васкуляризующий брактею, отходит в месте отделения вниз под углом, близким к 90°, в части коры между смоляными каналами, затем выполаживается и следует параллельно пучку, идущему в семенную чешую, а в месте отделения брактеи от семенной чешуи резко отгибается кверху и входит в основание брактеи.
Брактея у основания более или менее треугольная в поперечном сечении, шириной 2.5–3 мм, толщиной до 1.5 мм, отделяющаяся от семенной чешуи в ее основании, начиная с краев (рис. 3, г, ж), свободная часть достаточно короткая, до 2.5 мм; кора брактеи паренхиматозная, смоляные каналы расположены по бокам от проводящего пучка (рис. 3, г).
Семенные чешуи деревянистые, длиной до 17 мм, шириной 3–4 мм у основания, толщиной около 1.3–2 мм, шириной в средней части 7–10 мм, утолщающиеся ближе к апофизу (рис. 2, а, б, г, е; 3, в; 6, д). Апофизы преимущественно поперечно-ромбические до пятиугольных, c закругленным верхним краем, шириной 7–11.4 мм, высотой около 5–8 мм. В основании шишек они сильно приподнятые (3–5 мм), пирамидальные, с отчетливым поперечным килем, несущие выпуклое поперечно-ромбическое дорсальное умбо (табл. IX , фиг. 1, 2, 7, 9, 10, 13, 14 ) шириной 2–2.5 мм, слегка отогнутое книзу. Наличие/отсутствие мукро невозможно установить. Ближе к верхушечной части апофизы становятся слабовыпуклыми, конфигурация умбо неясна (табл. IX , фиг. 3, 4).
Семенные чешуи в средней части шишки отходят от шишечной оси под углом 40°–45° (рис. 1, а). В основании семенной чешуи имеется уплощенный проводящий пучок шириной 2–2.5 мм, высотой 200–270 мкм. Основная ткань под проводящим пучком почти полностью замещена сплошным массивом склеренхимы толщиной до 300 мкм, сложенным клетками диаметром 20–30 мкм (рис. 2, а; 3, а–ж). На границе основной ткани и абаксиальной коры имеются однорядно расположенные смоляные каналы диаметром до 90 мкм. По краям от проводящего пучка также имеются крупные смоляные каналы диаметром до 90 мкм (рис. 6, а). Смоляные каналы выстланы однослойным эпителием (рис. 6, г). Клетки эпителия тонкостенные, продольно вытянутые, шириной около 20 мкм. Прилегающие непосредственно к эпителию клетки не склерифицированы, 20–40 мкм в поперечном сечении, длиной до 110 мкм.
Абаксиальная кора ниже смоляных каналов паренхиматозная (диаметр клеток 10–30 мкм); внешняя часть абаксиальной коры образована одним слоем крупных, столбчатых, относительно тонкостенных клеток высотой до 150 мкм, шириной до 50 мкм, ориентированных почти перпендикулярно к поверхности семенной чешуи и обычно заполненных содержимым, в краевых зонах замещающихся тонким (толщиной в одну–две клетки) слоем гиподермальных волокон (рис. 2, а; 6, а). Адаксиальная кора тонкая, не более 50 мкм, паренхиматозная, диаметр клеток 5–20 мкм. В центральной части имеется отчетливая межсеменная перегородка, разграничивающая две семенных лунки (рис. 2, а; 3, а–в). Сначала она широкая, соединяющаяся с шишечной осью, к средней части семенных лунок отделяется от оси и становится треугольной в поперечном сечении, высотой около 400 мкм, шириной у основания около 500 мкм, сложена толстостенными паренхиматозными клетками диаметром 5–30 мкм. Постепенно перегородка становится ниже и уже, и полностью исчезает к верхней трети семенной чешуи.
Уплощенный проводящий пучок начинает расщепляться на отдельные пучки на уровне средней части семенных лунок, либо непосредственно за ними. Образовавшиеся проводящие пучки расположены однорядно, среди них обычно выделяется более широкий центральный пучок (рис. 2, б, г).
За семенными лунками, примерно в средней части, семенная чешуя шириной 7–10 мм, серпообразная в поперечном сечении; проводящие пучки семенной чешуи достигают максимального числа 12–15. Пучки коллатеральные, округлые либо почковидные, отчетливо выпуклые со стороны флоэмы, с достаточно хорошо выраженными рядами трахеид и слабо выраженными ксилемными лучами, без смоляных ходов (рис. 2, в, д). Ширина проводящих пучков в средней части семенной чешуи до 200 мкм, высота 120–150 мкм. Трахеиды толстостенные, диаметром до 10 мкм. Крупные склерифицированные массивы в основной ткани приурочены к проводящим пучкам (рис. 2, б, д, е). Смоляные каналы в абаксиальной коре становятся преимущественно мелкими, до 50–100 мкм в диаметре; адаксиальная кора семенной чешуи постепенно утолщается до 200–250 мкм (рис. 2, б, в; 3, в), в ней появляются адаксиальные смоляные каналы разного диаметра; изредка также смоляные каналы, образовавшиеся de novo, наблюдаются в семенной перегородке (рис. 2, г). По мере утолщения адаксиальной коры семенной чешуи в дистальном направлении граница между основной тканью и корой стирается, в толще семенной чешуи появляются многочисленные хаотично расположенные склеренхимные гнезда, во внешней части абаксиальной и адаксиальной коры образуется тонкий слой гиподермальных волокон толщиной в две–три клетки; смоляные каналы равномерно распределены в толще чешуи (рис. 2, в).
Ближе к основанию апофиза семенная чешуя в среднем утолщается до 1450 мкм и становится субтреугольной в сечении. Здесь адаксиальная кора утолщается до 550 мкм. В ней увеличивается количество смоляных каналов, которые расположены здесь беспорядочно. Смоляные каналы диаметром до 100 мкм. Абаксиальная кора утолщается до 900 мкм. Мелкие абаксиальные смоляные каналы в ней также присутствуют, расположены беспорядочно. Проводящие пучки начинают терять горизонтальную ориентировку, образуют небольшие аркообразные группы (рис. 2, е).
В основании апофиза склерификация не выражена (рис. 6, д). Проводящие пучки становятся немного мельче, диаметром 75–200 мкм, в центральной части расположены двурядно. Смоляные каналы, в основном, мелкие, диаметром 50–100 мкм, расположены беспорядочно, но группируются преимущественно вокруг проводящих пучков. Иногда смоляные каналы в апофизе могут расширяться до 250 мкм.
Семена узко-эллипсоидальные, длиной 2.5–4 мм, шириной 1.2–1.6 мм, с заостренным микропилярным концом, закругленной халазой и хорошо развитым полуовальным персистирующим крылом (рис. 4, а–в). Длина семени вместе с крылом до 9 мм. Крыло семени сильно утолщено у халазы (рис. 3, б; 4, б). Теста состоит из саркотесты, склеротесты и эндотесты (рис. 4, ж). Саркотеста тонкая, толщиной около 100 мкм. Склеротеста развита неравномерно, ребристая; максимальная толщина 280 мкм, на более тонких участках – 100 мкм. Проводящие пучки в тесте отсутствуют. Нуцеллус в верхней части свободен от интегумента, тонко кутинизированный, длиной 2.3 мм, шириной 1.2 мм (рис. 4, г–е). Клетки нуцеллуса продольно вытянутые, прямоугольные, длиной до 40 мкм, шириной до 20 мкм (рис. 4, и). Мегаспоровая мембрана толщиной около 3 мкм, имеет ячеистое строение (рис. 4, к).
Сравнение и замечания. Новый вид демонстрирует все признаки, характерные для шишек представителей рода Pinus L., согласно Миллеру (Miller, 1976): утолщенные апексы семенных чешуй; единый округлый след брактеи и семенной чешуи; смоляные каналы, входящие в основание семенной чешуи только абаксиально; адаксиально выпуклые проводящие пучки семенной чешуи. Также у представителей данного рода имеются вертикальные смоляные каналы в ксилеме шишечной оси и небольшая субтреугольная брактея, отделяющаяся, начиная с краев.
Признаки морфологии P. bukatkinii sp. nov. не позволяют отнести его к какому-либо из двух современных подродов рода сосна (Pinus или Strobus). Сильно приподнятые апофизы с выступающим дорсальным умбо встречаются у представителей обоих подродов, например, у Pinus gerardiana Wallich ex D. Don (подрод Strobus) и P. sabiniana Douglas ex D. Don (подрод Pinus), поэтому данный признак не является существенным. Удлиненно-цилиндрические семенные шишки также встречаются у представителей обоих подродов (Farjon, 2010).
По анатомии изученные шишки более сходны с представителями подрода Pinus. Например, проводящие пучки в средней части чешуи у P. bukatkinii sp. nov. расположены однорядно, тогда как для подрода Strobus указывается их двурядное расположение (Yamada, Yamada, 2017). Отсутствие склерификации эпителиальных клеток смоляного канала и прилегающего к ним кольца клеток в семенной чешуе характерно для представителей обоих подродов (Yamada, Yamada, 2017). Несклерифицированная кора шишечной оси в сочетании со склерифицированной корой семенной чешуи – признак, характерный для шишек подсекции Ponderosae подрода Pinus, а также для некоторых представителей Contortae и Australes, тогда как у представителей подрода Strobus склерификация шишечной оси и чешуй отсутствует (Radais, 1894; Shaw, 1914; Miller, 1976). Утолщенное основание крыла семени также характерно для некоторых представителей подрода Pinus (Shaw, 1914; Miller, 1978).
От всех ископаемых и современных сосен, для которых изучено анатомическое строение, новый вид отличается сильно развитой склерификацией основной ткани семенных чешуй непосредственно под проводящими пучками, причем признак прослеживается от самого основания семенной чешуи (склерификации подвергаются уже клетки сердцевины общего пучка шишечного комплекса сразу после отхождения следа брактеи). Сильная склерификация основной паренхимы характерна для некоторых представителей рода Pityostrobus и современного Pseudolarix Gordon (Miller, 1976). Например, у Pityostrobus hokodzensis Ratzel, Rothwell, G. Mapes, R.H. Mapes et Doguzhaeva из нижнемеловых отложений Северного Кавказа, склерификация также прослеживается еще в следе семенной чешуи и сходным образом распределяется в ее тканях (Ratzel et al., 2001). У Pityostrobus hallii Miller из верхнего мела Мериленда, США, склеренхима образует в основной паренхиме сплошной массив и дистальнее перемещается на адаксиальную сторону, формируя адаксиальную гиподерму (Miller, 1974). По данным Миллера, среди современных сосен склерификация основной паренхимы может встречаться только в расширенном апексе семенной чешуи у представителей подрода Pinus (Miller, 1976). Среди ископаемых сосен только для P. cliffwoodensis из сантона Нью-Джерси, США (Miller, Malinky, 1986) указано наличие гнезд склереид в паренхиматозной внешней коре шишечной оси и в основной паренхиме основания семенной чешуи, которые далее преобразуются в отдельные тяжи, чтобы в дистальной части образовать адаксиальную гиподерму. К сожалению, сами гнезда склереид в работе не проиллюстрированы, но, судя по описанию, характер склерификации P. cliffwoodensis существенно отличается от такового у P. bukatkinii sp. nov. В отличие от нового вида, P. cliffwoodensis имеет яйцевидно-конические шишки c сердцевиной и ксилемой примерно вдвое большего диаметра, имеющей смоляные каналы в конце прироста и выраженные годичные кольца; межсеменной гребень отсутствует, наличие/отсутствие крыла у семени не установлено.
Раннемеловые P. yorkshirensis из готерив–барремских отложений Йоркшира, Англия (Ryberg et al., 2012), и P. belgica из вельда Бельгии (Alvin, 1960) наиболее близки по возрасту к новом виду. Из них наибольшее сходство с P. bukatkinii sp. nov. демонстрирует P. yorkshirensis. Но у P. yorkshirensis шишки конические, тогда как у нового вида они цилиндрические, с более выступающими апофизами. Кроме того, у P. bukatkinii sp. nov. более частые смоляные каналы в древесине шишечной оси, иногда расположенные не только вблизи сердцевины, но и в средней части кольца прироста. Смоляные каналы в коре шишечной оси у P. yorkshirensis проходят в средней ее части, а внешняя кора склерифицирована, в отличие от коры P. bukatkinii sp. nov. Брактея у P. yorkshirensis, предположительно, очень короткая; смоляные каналы в ней сохранились плохо. Авторы указывают на их краевое положение, тогда как у нового вида они расположены близко к проводящему пучку. Адаксиальные смоляные каналы в семенной чешуе P. yorkshirensis отсутствуют. Характер склерификации семенной чешуи также иной: сильная склерификация наблюдается ниже смоляных каналов, а также в краевых зонах семенной чешуи, где склерифицированы все ткани. Для P. yorkshirensis неизвестен тип отхождения следа брактеи и семенной чешуи.
У P. belgica из вельда Бельгии (Alvin, 1960) шишки овально-конические; сердцевина и древесина несколько массивнее, чем у P. bukatkinii sp. nov. Кроме того, внешняя кора шишечной оси у P. belgica склерифицирована; склереиды встречаются также и во внутренней коре. Гиподерма семенной чешуи у этого вида была сильно склерифицирована, а межсеменной гребень предположительно отсутствовал. Кроме того, семена у P. belgica значительно крупнее, чем у нового вида (длина тела семени достигает 9 мм).
У P. mutoi из коньяка Хоккайдо, Япония (Saiki, 1996), шишки цилиндрические, с выступающими апофизами, но значительно крупнее, чем у нового вида – длиной до 20 см и диаметром 6 см. Кроме того, у P. mutoi внешняя кора шишечной оси склерифицированная, семенные чешуи без межсеменного гребня, семена в плане почти шаровидные, а длина крыла семени составляет только половину длины семенной чешуи. К сожалению, у японского вида сохранились не все признаки анатомии, что затрудняет сравнение.
Среди кайнозойских видов паренхиматозную внешнюю кору шишечной оси в сочетании со склерифицированной корой семенной чешуи имеют: P. driftwoodensis Stockey из эоцена Британской Колумбии и P. escalantensis Banks, Ortiz-Sotomayor et Hartman из олигоцена Британской Колумбии, Канада (Banks et al., 1981; Stockey, 1983), P. avonensis Miller из олигоцена Монтаны (Miller, 1969), P. buchananii Underwood et Miller из олигоцена Вашингтона (Underwood, Miller, 1980) и P. matthewsii McКown, Stockey et Schweger из плиоцена Юкона, Канада (McKown et al., 2002). Из них наибольшее сходство с P. bukatkinii sp. nov. обнаруживают P. driftwoodensis и P. escalantensis, имеющие цилиндрические шишки с сильно выступающими апофизами (Banks et al., 1981; Stockey, 1983).
Отличие нового вида от P. escalantensis заключается в том, что у последнего шишки значительно длиннее и уже, брактея склерифицирована, вертикальные смоляные каналы в ксилеме расположены не у границы с сердцевиной, а примерно на одной трети расстояния между внутренней и внешней границей ксилемы и, предположительно, маркируют границу между годовыми приростами. Кроме того, у P. escalantensis сердцевина и проводящий цилиндр втрое шире, чем у нового вида, а в ксилеме следа шишечного комплекса наблюдаются вертикальные смоляные каналы. Смоляные каналы коры не обнаруживают существенного расширения в месте ветвления. Семена P. escalantensis не сохранились.
У P. driftwoodensis шишки немного мельче, смоляные каналы в ксилеме шишечной оси намного крупнее и более частые, чем у нового вида. Смоляные каналы коры шишечной оси не расширяются в месте ветвления так сильно, как у нового вида, межсеменной гребень отсутствует, а в общем следе семенной чешуи и брактеи имеются смоляные каналы. Кроме того, смоляные каналы почти на всем протяжении семенной чешуи абаксиальные, в отличие от нового вида. Наличие/отсутствие крыльев у семян неизвестно.
Остальные виды с паренхиматозной внешней корой шишечной оси существенно отличаются от нового вида по морфологии, поскольку имеют конические шишки с невыступающими апофизами.
Другие кайнозойские виды: P. prinstonensis Stockey и P. arnoldii Miller из эоцена Британской Колумбии (Stockey, 1983; Klymiuk et al., 2011), P. wolfei Miller из эоцена Вашингтона, США (Miller, 1974), и P. burtii Miller из миоцена Массачусетса, США (Miller, 1978), имеют сильно склерифицированную внешнюю кору шишечной оси и в целом существенно отличаются от нового вида по морфологии и анатомии. Из них только у P. prinstonensis шишки цилиндрические.
Материал. 9 экз., сохранившихся в виде лигнитизированных фрагментов семенных шишек.
АССОЦИИРУЮЩАЯ ПЫЛЬЦА
Девять пыльцевых зерен обнаружены на поверхности семян голотипа P. bukatkinii sp. nov. (рис. 5, а–ж). Пыльцевые зерна двухмешковые, билатерально симметричные. Очертания тела эллиптические или округлые в полярном положении, плоско-выпуклые – в боковом положении; размер тела 57 (49–67.7) × 42.6 (41.2–58) мкм (ширина × длина). Очертания мешков эллиптические в полярном положении, двояковыпуклые – в боковом положении. Оба мешка сходного размера и формы, для удобства мы их обозначаем как левый и правый; размеры левого мешка 55.9 (48.9–66.4) × 20.6 (13.6–37.3) мкм, правого – 55.2 (51.5–57.8) × 22.2 (10.8–38.1) мкм. Лептома представлена утонченной областью на дистальной поверхности. На одном пыльцевом зерне, лежащем в дистальном положении, лептома шириной 5 мкм с гладкой апертурной мембраной со складками (рис. 5, е). Поверхность щита (проксимальная поверхность тела) гранулярная или мелкобугорчатая, скульптурные элементы размером 0.3–0.5 мкм (рис. 5, а–г, ж). Поверхность мешков ямчатая (рис. 5, д, е).
Три пыльцевых зерна обнаружены на адаксиальной поверхности семенной чешуи голотипа (рис. 5, з, и). Пыльцевое зерно, лежащее проксимальной стороной (рис. 5, з) – общее описание сходно с приведенным выше. Тело пыльцевого зерна 60.3 мкм шириной и 47.5 мкм длиной; левый мешок 65.1 × 18.3 мкм, правый – 59 × 17 мкм. Данное пыльцевое зерно значительно отличается большим размером гранул скульптуры поверхности щита, составляющим 1–1.4 мкм (рис. 5, з).
Два пыльцевых зерна, лежащих дистальной стороной (рис. 5, и) – общее описание сходно с приведенным выше. Тело пыльцевого зерна 46.8 и 49 мкм шириной и 51.6 мкм длиной (для второго зерна), левый мешок 52.9 × 23.7 мкм у одного и ? × 29.7 мкм у второго пыльцевого зерна, размер правого мешка можно измерить только у первого пыльцевого зерна – 50.8 × 32.5 мкм.
Пыльцевое зерно из микропиле (рис. 4, з). У этого пыльцевого зерна видны только мешки, сходные по морфологии с таковыми у пыльцевых зерен, обнаруженных на поверхности семян и семенных чешуй.
ГАЛЕРЕИ
В шишечных осях двух фрагментов шишек P. bukatkinii sp. nov. обнаружены следы повреждения фитофагами, представленные преимущественно продольными галереями различного диаметра (400–1000 мкм), затрагивающими, в основном, внешнюю часть ксилемы и флоэму шишечной оси и следов, васкуляризующих шишечные чешуи; семена и чешуи при этом не повреждены. Галереи заполнены плотными скоплениями пеллетов эллипсоидальной формы, смешанных с буровой мукой. В обоих фрагментах шишек наблюдаются две относительно крупные галереи, диаметром около 1000 мкм, идущие более или менее вдоль шишечной оси (рис. 1, в, д; 3, а, в). В шишке № 5851/2 наблюдается только предположительный ход имаго. В шишке № 5851/9 можно проследить, как ход следует от ее основания до верхушки, где разворачивается и направляется к основанию шишки. От крупной галереи отходят намного более узкие извилистые галереи, ориентированные как продольно, так и поперечно (рис. 3, в). Некоторые из них входят в семенные чешуи, затрагивая ксилему (рис. 3, г). Размеры пеллетов соотносятся с диаметром галерей. В крупных галереях диаметром 1000 мкм максимальная длина пеллетов составляет 200 мкм, тогда как в самых мелких галереях (диаметром до 400 мкм) максимальная длина пеллетов – 80 мкм. Крупная галерея, идущая от основания оси и разворачивающаяся к ее основанию, скорее всего, является маточным ходом, поскольку от нее расходятся более мелкие (предположительно, личиночные) ходы. Отличие заключается в том, что у Conophthorus Hopkins самка покидает шишку вблизи верхушки, тогда как в ископаемых шишках крупный ход разворачивается у верхушки и вновь направляется к основанию (рис. 5, м).
В ископаемой летописи известны сходные галереи в шишках соснового Pityostrobus milleri Falder, Rothwell, G. Mapes, R.H. Mapes et Doguzhaeva из раннего мела Кавказа, Россия, сравниваемые с ходами современного североамериканского вида Conophthorus resinosae Hopkins (Falder et al., 1998). У Pityostrobus hokodzensis из того же местонахождения также обнаружены галереи, но, по-видимому, оставленные другим фитофагом (Ratzel et al., 2001). Также известны две крупные продольные галереи в шишке Pinus escalantensis из олигоцена Британской Колумбии, Канада (Banks et al., 1981). Во всех перечисленных случаях нет прямых доказательств того, что шишки повреждены жуками-короедами, поскольку сами фитофаги не сохранились. Вместе с тем, древнейшая достоверная находка короеда известна из раннего мела Ливана (Kirejtshuk et al., 2009). Исходя из типа повреждений у шишек нового вида, мы полагаем, что наиболее вероятными вредителями этой древней сосны были именно короеды.
ОБСУЖДЕНИЕ
Отнесение анатомических семенных шишек соснового типа к роду Pinus основано на концепции Миллера, сформулированной им в результате изучения обширного материала по современным видам этого рода и его сопоставления с ископаемыми представителями сосновых; данная концепция в настоящее время является общепринятой (Miller, 1976, 1977; Banks et al., 1981; McKown et al., 2002; Ryberg et al., 2012; и др.). Несмотря на это, необходимо понимать некоторую условность отнесения ископаемых семенных шишек сосновых к роду Pinus, что отражает общую проблему отнесения ископаемого анатомического, даже детально охарактеризованного материала, представляющего собой изолированные органы, к современным таксонам (см. Баженова, Баженов, 2019). Соотношение между разными органами большинства ископаемых растений можно более или менее надежно установить только в единичных случаях. Лишь для некоторых ископаемых сосен анатомической сохранности имеются данные об ассоциирующих с шишками листьях (Banks et al., 1981; Ryberg et al., 2012), но все же нет данных о строении побегов и микростробилов. Единственный ископаемый вид, для которого была предложена организменная концепция – эоценовый P. arnoldii. По данным исследователей, семенные шишки этого вида имеют строение, характерное для представителей подрода Pinus, тогда как строение древесины и укороченных побегов с листьями указывает на принадлежность к подроду Strobus (Klymiuk et al., 2011). Такие данные указывают на невозможность отнесения ископаемых сосен, кроме наиболее молодых представителей, к тому или иному подроду в случае, если не восстановлены прижизненные связи между семенными и вегетативными органами.
В основе концепции Миллера, разработанной для идентификации шишек рода Pinus, лежит комплекс из четырех главных диагностических признаков: утолщающиеся в верхней части семенные чешуи, имеющие апофизы с умбо; единый след, васкуляризующий семенную чешую и брактею; смоляные каналы, входящие в основание семенной чешуи абаксиально; адаксиально выпуклые проводящие пучки семенной чешуи (Miller, 1976). Эти признаки иногда не признавались Миллером как единственно достаточные для отнесения семенных шишек к роду Pinus (см., напр., Miller, Robison, 1975; Miller, 1978). К ним добавляются такие характеристики, как наличие вертикальных смоляных каналов в ксилеме шишечной оси, наличие свободной брактеи, способ ее отделения от семенной чешуи (для сосен характерен краебежный способ) и степень ее васкуляризации (Miller, Robison, 1975; Miller, 1977). В то же время, наличие единого следа семенной чешуи и брактеи может являться необязательным признаком, поскольку не наблюдается у некоторых представителей современных сосен (Miller, 1978; Gernandt et al., 2011; Ryberg et al., 2012). У Pinus bukatkinii sp. nov., кроме четырех признаков, предложенных Миллером в качестве основных, имеются также дополнительные, а именно: наличие вертикальных смоляных каналов в ксилеме шишечной оси и свободная брактея, отделяющаяся краебежно. По нашему мнению, такие признаки, как способ отделения брактеи и степень ее васкуляризации, на самом деле не являются значимыми для идентификации шишек Pinus, поскольку они варьируют в пределах рода. Например, медиальное отделение брактеи характерно для P. matthewsii (McKown et al., 2002), а проводящий пучок, не доходящий до апикальной части брактеи, – для P. escalantensis (Banks et al., 1981).
В связи с находкой в средней юре семенных шишек, демонстрирующих морфолого-анатомические признаки Pinus, необходимо упомянуть гипотезу Миллера об архаичности строения семенных органов данного рода (Miller, 1976). Новая находка подтверждает предположение о том, что шишки, обладающие всеми основными морфолого-анатомическими признаками Pinus, лежат в основе разнообразия меловых Pityostrobus в понимании Миллера (Miller, 1976); соотношение этих двух родов с остальными представителями семейства до конца не ясно. Наличие у среднеюрского Pinus bukatkinii sp. nov. всего комплекса признаков рода в сформированном виде указывает на вероятное значительно более раннее возникновение отдельных признаков. Имеющиеся в настоящее время данные о репродуктивных структурах древнейших сосновых и форм, предположительно относимых к семейству, настолько скудны, что не дают сколько-нибудь надежных оснований даже для предварительных филогенетических гипотез. Древнейшими предположительными сосновыми являются Compsostrobus neotericus из верхнего триаса Северной Каролины (Delevoryas, Hope, 1973, 1987; Taylor et al., 2009) и Schizolepidopsis liasokeuperianus (С.F.W. Braun) Doweld, распространенный в среднем–верхнем триасе Евразии (достоверность пермских находок Schizolepidopsis не подтверждена: Domogatskaya, Herman, 2019), однако анатомия этих шишек неизвестна, что не дает оснований для их полноценного использования в реконструкции филогении семейства сосновых. В то же время, имеющиеся находки указывают на то, что непосредственных предков рода Pinus, вероятнее всего, следует искать в ранней юре.
Список литературы
Баженов А.В., Баженова Н.В. О возрасте древнейших семенных шишек сосны // Палеострат–2022. Годичн. собр. (научн. конф.) секции палеонтол. МОИП и Московского отд. Палеонтол. об-ва при РАН. Тез. докл. / Ред. Голубев В.К., Назарова В.М. М.: ПИН РАН, 2022. С. 8–9.
Баженова Н.В., Баженов А.В. Стебли нового осмундового папоротника из средней юры Курской области, европейская часть России // Палеонтол. журн. 2019. № 5. С. 100–108.
Alvin K.L. On Pseudoaraucaria Fliche emend., a genus of fossil pinaceous cones // Ann. Bot. N. Ser. 1957. V. 21. P. 33–51.
Alvin K.L. Further conifers of the Pinaceae from the Wealden formation of Belgium // Mem. Inst. R. Sci. Natur. Belg. 1960. V. 146. P. 1–39.
Banks H.P., Ortiz-Sotomayor A., Hartman C.M. Pinus escalantensis, sp. nov., a new permineralized cone from the Oligocene of British Columbia // Botan. Gaz. 1981. V. 142. № 2. P. 286–293.
Delevoryas T., Hope R.C. Fertile coniferophyte remains from the Late Tiassic Deep River basin, North Carolina // Amer. J. Bot. 1973. V. 60. № 8. P. 810–818.
Delevoryas T., Hope R.C., Further observations on the late Triassic conifers Compsostrobus neotericus and Voltzia andrewsii // Rev. Palaeobot. Palynol. 1987. V. 51. № 1. P. 59–64.
Domogatskaya K.V., Herman A.B. New species of the genus Schizolepidopsis (conifers) from the Albian of the Russian high Arctic and geological history of the genus // Cret. Res. 2019. V. 97. P. 73–93.
Falder A.B., Rothwell G.W., Mapes G. et al. Pityostrobus milleri sp. nov., a pinaceous cone from the Lower Cretaceous (Aptian) of southwestern Russia // Rev. Palaeobot. Palynol. 1998. V. 103. № 3–4. P. 253–261.
Farjon A. A Handbook of the World’s Conifers. V. II. Leiden–Boston: Brill, 2010. 1111 p.
Gernandt D.S., León-Gómez C., Hernández-León S. et al. Pinus nelsonii and a cladistic analysis of Pinaceae ovulate cone characters // Syst. Botan. 2011. V. 36. № 3. P. 583–594.
Kirejtshuk A.G., Azar D., Beaver R.A. et al. The most ancient bark beetle known: a new tribe, genus and species from Lebanese amber (Coleoptera, Curculionidae, Scolytinae) // Syst. Entomol. 2009. V. 34. № 1. P. 101–112.
Klymiuk A.A., Stockey R.A., Rothwell G.W. The first organismal concept for an extinct species of Pinaceae: Pinus arnoldii Miller // Int. J. Plant Sci. 2011. V. 172. № 2. P. 294–313.
McKown A.D., Stockey R.A., Schweger C.E. New species of Pinus subgenus Pinus subsection Contortae from Pliocene sediments of Ch’ijee’s Bluff, Yukon Territory, Canada // Int. J. Plant Sci. 2002. V. 163. № 4. P. 687–697.
Leslie A.B., Glasspool I., Herendeen P.S. et al. Pinaceae-like reproductive morphology in Schizolepidopsis canicularis sp. nov. from the Early Cretaceous (Aptian–Albian) of Mongolia //Amer. J. Bot. 2013. V. 100. № 12. P. 2426–2436.
Matsunaga K.K.S., Herendeen P., Herrera F. et al. Ovulate cones of Schizolepidopsis ediae sp. nov. provide insights into the evolution of Pinaceae // Int. J. Plant Sci. 2021. V. 182. № 6. P. 490–507.
Miller C.N. Pinus avonensis, a new species of petrified cones from the Oligocene of Western Montana // Amer. J. Bot. 1969. V. 56. № 9. P. 972–978.
Miller C.N. Pinus wolfei, a new petrified cone from the Eocene of Washington // Amer. J. Bot. 1974. V. 61. № 10. P. 772–777.
Miller C.N. Early evolution in the Pinaceae // Rev. Palaeobot. Palynol. 1976. V. 21. № 2. P. 101–117.
Miller C.N. Pityostrobus lynni (Berry) comb. nov., a Pinaceous seed cone from the Paleocene of Virginia // Bull. Torrey Bot. Club. 1977. V. 104. № 1. P. 5–9.
Miller C.N. Pinus burtii, a new species of petrified cones from the Miocene of Martha’s Vineyard // Bull. Torrey Bot. Club. 1978. V. 105. № 2. P. 93–97.
Miller C.N., Malinky J.M. Seed cones of Pinus from the Late Cretaceous of New Jersey, USA // Rev. Palaeobot. Palynol. 1986. V. 46. № 3–4. P. 257–272.
Miller C.N., Robison C.R. Two new species of structurally preserved pinaceous cones from the Late Cretaceous of Martha’s Vineyard Island, Massachusetts // J. Paleontol. 1975. V. 49. № 1. P. 138–150.
Ohsawa T.A., Nishida M., Nishida H. Structure and affinities of the petrified plants from the Cretaceous of northern Japan and Saghalien. XII. Obirastrobus gen. nov., petrified pinaceous cones from the Upper Cretaceous of Hokkaido // Bot. Mag. Tokyo. 1992. V. 105. P. 461–484.
Radais M. Contribution a l’étude de l’anatomie comparée du fruit des conifères // Ann. Sci. Natur. Bot. Paris. 1894. Ser. 7–19. P. 165–368.
Ratzel S.R., Rothwell G.W., Mapes G. et al. Pityostrobus hokodsensis, a new species of pinaceous cone from the Cretaceous of Russia // J. Paleontol. 2001. V. 75. № 4. P. 895–900.
Rothwell G.W., Mapes G., Stockey R.A. et al. The seed cone Eathiestrobus gen. nov.: fossil evidence for a Jurassic origin of Pinaceae // Amer. J. Bot. 2012. V. 99. № 4. P. 708–720.
Ryberg P. E., Rothwell G.W., Stockey R.A. et al. Reconsidering relationships among stem and crown group Pinaceae: oldest record of the genus Pinus from the Early Cretaceous of Yorkshire, United Kingdom // Int. J. Plant Sci. 2012. V. 173. № 8. P. 917–932.
Saiki K. Pinus mutoi (Pinaceae), a new species of permineralized seed cone from the Upper Cretaceous of Hokkaido, Japan // Amer. J. Bot. 1996. V. 83. № 12. P. 1630–1636.
Shaw G.R. The genus Pinus // Publications of Arnold Arboretum. № 5. Riverside, Cambridge: Mass, 1914. 96 p.
Smith S.Y., Stockey R.A. A new species of Pityostrobus from the Lower Cretaceous of California and its bearing on the evolution of Pinaceae // Int. J. Plant Sci. 2001. V. 162. № 3. P. 669–681.
Smith S.Y., Stockey R.A. Permineralized pine cones from the Cretaceous of Vancouver Island, British Columbia // Int. J. Plant. Sci. 2002. V. 163. № 1. P. 185–196.
Stockey R.A. Middle Eocene Pinus remains from British Columbia //Bot. Gaz. 1983. V. 145. № 2. P. 262–274.
Taylor T.N., Taylor E.L., Krings M. Paleobotany: The Biology and Evolution of Fossil Plants. N.Y.: Acad. Press, 2009. 1230 p.
Underwood C., Miller C.N. Pinus buchananii, a new species based on a petrified cone from the Oligocene of Washington // Amer. J. Bot. 1980. V. 67. № 7. P. 1132–1135.
Yamada M., Yamada T. Ovuliferous scale anatomy of Pinus species and its value for classification above the species level // Bot. J. Linn. Soc. 2017. V. 183. № 4. P. 633–643.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал