

Палеонтологический журнал, 2023, № 1, стр. 54-62
Вид Palmatolepis tatarica Ovnatanova et Gatovsky, 2020: история его выделения и интерпретации в мировой литературе по конодонтам позднего девона
Н. С. Овнатанова a, *, Л. И. Кононова b, **
a Всероссийский научно-исследовательский геолого-разведочный нефтяной институт
105118 Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: ovnatanova@gmail.com
** E-mail: 2216614@list.ru
Поступила в редакцию 10.02.2022
После доработки 25.08.2022
Принята к публикации 25.08.2022
- EDN: FJVOMK
- DOI: 10.31857/S0031031X23010105
Аннотация
Статья посвящена одному из широко развитых в фаменских отложениях мира представителю конодонтов группы Palmatolepis subperlobata, название которого в течение более полувека менялось от Pa. subperlobata subsp. a Helms, 1963 до Pa. subperlobata helmsi Ovnatanova, 1976 и затем Pa. subperlobata tatarica Ovnatanova et Gatovsky, 2020. В статье этот подвид рассматривается как самостоятельный вид, обсуждаются сходство и отличия его голотипа и голотипов видов, предложенных в качестве замены преоккупированного названия. Предлагается авторская схема филогенетического развития группы Pa. subperlobata.
ВВЕДЕНИЕ
Более чем полуторавековая история изучения конодонтов, в т.ч. представителей позднедевонского рода Palmatolepis Ulrich et Bassler, приоткрыла многие тайны возникновения и происхождения видов этого рода, что нашло отражение в многочисленных публикациях (Müller, 1956; Ziegler, 1962, 1969, 1971 и др.). По недавним подсчетам, в этом роде предложено свыше 200 видовых и подвидовых названий (Becker, Hartenfels, 2016), и их число продолжает увеличиваться. Неоднократные попытки выделения подродов не сыскали множества сторонников, как и система мультиэлементных родов Е. Дзика (Dzik, 2006); видимо, разукрупнение этого рода еще впереди. Несомненным достижением явилась разработанная Й. Хелмсом во второй половине ХХ в. филоморфогенетическая схема развития видов рода Palmatolepis в привязке к аммонитовой шкале (Helms, 1963), а в последующем – и к конодонтовой зональности. В этой схеме уже наметилось выделение ряда ветвей, которые Хелмс считал подродами, но они продолжают рассматриваться как группы близких друг к другу видов.
Последующий прогресс в изучении конодонтов был связан, в первую очередь, с установлением филогенетических cвязей в развитии рода Palmatolepis в фаменское время. Подробные результаты этих исследований освещены в монографии К. Цзы и В. Циглера (Ji, Ziegler, 1993) на основании обширной коллекции, полученной из разреза Лали (Lali) в Южном Китае. Этими исследователями впервые рассматривается происхождение и филогения группы Pa. subperlobata Branson et Mehl, 1934. Хотя виды этой группы относительно немногочисленны, они имеют существенное стратиграфическое значение, а Pa. subperlobata недавно предложено использовать в качестве маркера нижней границы фаменского яруса (Spaletta et al., 2017). Один из таксонов, входящих в состав этой группы, – это подвид Pa. subperlobata tatarica Ovnatanova et Gatovsky, 2020. Название и таксономический статус этого подвида вызывают разногласия, в связи с чем мы здесь попытаемся более обстоятельно рассмотреть его характеристику и место в общей филоморфогенетической схеме группы Pa. subperlobata.
Виды рода Palmatolepis различаются на основании особенностей морфологии Ра-элементов. В статье при их характеристике использованы морфологические термины, объяснение которых дано ранее (Барсков и др., 1987; Ovnatanova, Kononova, 2008). В аппарат входят два симметричных Ра-элемента, правый и левый, не отличающиеся по форме и скульптуре друг от друга, что упрощает анализ морфологии и сравнение таксонов этого рода.
ВИД PALMATOLEPIS TATARICA
Первым обратил внимание на формы, похожие на Pa. subperlobata, но отличающиеся пережимом внутренней лопасти, Хелмс (Helms, 1963). В его статье название P. subperlobata n. subsp. a появляется при описании филоморфогенеза рода Palmatolepis (Helms, 1963, с. 458); кратко приведены признаки, отличающие его от Pa. subperlobata. Затем название P. (Pand.) subperlobata n. subsp. a находим в подписи к филоморфогенетической схеме (Helms, 1963, с. 468) и в объяснениях к фототаблицам к форме Palmatolepis (Pand.) subperlobata subsp. a (Helms, 1963, с. 478, 480, 482). Таким образом, Хелмс не дал этому подвиду формального описания, рассматривая такие формы только как уклоняющиеся от основного морфотипа. Аналогичные “подвиды” с буквенными названиями он предложил выделять и у ряда других видов рода Palmatolepis, например, Pa. (Panderolepis) serrata n. subsp. a, считая их не заслуживающими статуса самостоятельного таксона.
Н.С. Oвнатанова (1976), учитывая накопившийся за прошедшие годы материал, пришла к выводу о том, что такие формы заслуживают выделения в качестве самостоятельного подвида, и дала ему название Pa. subperlobata helmsi Ovnatanova, 1976, сделав его валидным. Это решение нашло поддержку среди части конодонтологов, и название Pa. subperlobata helmsi присутствует в ряде публикаций (Кононова, 1979; Наседкина, Плотникова, 1979; Барсков и др., 1987; Çapkinoğlu, 1997, 2005; Стрельченко, Кручек, 2013). Одновременно автоматически появился номинативный подвид Pa. subperlobata subperlobata Branson et Mehl, его упоминание можно найти в целом ряде работ (напр., Деулин, 2000; Плотицын и др., 2015), но многие продолжали считать выделение подвидов у этого вида нецелесообразным, ограничиваясь только видовым названием. Поэтому элементы Pa. subperlobata helmsi нередко фигурируют как принадлежащие Pa. subperlobata s.l. (Халымбаджа, Чернышева, 1978; Халымбаджа, 1981; Wang, 1989; Кузьмин, Мельникова, 1991; Ji, Ziegler, 1993; Metzger, 1994; Savage, Yudina, 1999).
Позднее, при изучении фаменских отложений Северо-Западного Таиланда, Н. Сэведж (Savage, 2013) выделил подвид Pa. subperlobata lapoensis, который отличается от номинативного подвида резко заостренной треугольной лопастью и заостренным задним концом; при этом, как и у Pa. subperlobata subperlobata, присутствует свободный лист и платформа практически гладкая (тонкошагреневая). Нами он рассматривается как самостоятельный вид.
В начале 2000-х гг. выяснилось, что, в соответствии со статьей 52.1 МКЗН (Международный…, 2000), название Pa. subperlobata helmsi должно считаться преокуппированным, поскольку В. Циглером на год ранее был назван в честь Хелмса вид Palmatolepis helmsi Ziegler, 1962. Г. Клаппер с соавт. (Klapper et al., 2004) посчитали возможным использовать в качестве замещающего названия (повысив ранг подвида до вида) Pa. lobicornis Schülke, 1995. Этому предложению последовали, например, Г. Сильверио и др. (Silvério et al., 2021, c. 213), хотя у них в синонимике он фигурирует со старым названием Pa. (Panderolepis) subperlobata subsp. a.
В 2006 г. Е. Дзик (Dzik, 2006, с. 138) описал в составе аппаратного рода Conditolepis новый вид C. linguiloba, рассматриваемый им как синоним Pa. superlobata helmsi, но не как формальное замещающее название. В описании этого нового вида отсутствует синонимика, а для желающих ознакомиться с ней приведен перечень некоторых статей (Wolska, 1967; Wang, 1989; Ji, Ziegler, 1993; Metzger, 1994) без указания номеров таблиц и фотографий с изображением вида.
Таким образом, перечисленные выше действия не устранили омонимии названия Pa. subperlobata helmsi, поэтому Н.С. Овнатанова и Ю.А. Гатовский (2020) предложили для него новое название Pa. subperlobata tatarica Ovnatanova et Gatovsky, nom. nov. с сохранением его голотипа (Овнатанова, 1976, табл. IX , фиг. 7 ), что удовлетворяет требованиям МКЗН.
Род Palmatolepis Ulrich et Bassler, 1926
Palmatolepis tatarica Ovnatanova et Gatovsky, 2020
Palmatolepis (Panderolepis) subperlobata subspecies а: Helms, 1963, с. 458, 468, 478, 480, 482, Taf. 1, fig. 19 ; Taf. 2, figs. 23, 24 ; Taf. 3, fig. 10 ; text-fig. 2.17.
Palmatolepis subperlobatа: Spasov, Filipović, 1966, pl. 3, figs. 1, 2; Glenister, Klapper, 1966, рl. 92, fig. 6 (только); Wolska, 1967, рl. 11, fig. 10 (только); Druce, 1976, pl. 63, fig. 3 a, b (только); Халымбаджа, Чернышева, 1978, с. 35, табл. 1 , фиг. 21 ; Наседкина, Плотникова, 1979, табл. 3 , фиг. 2; Халымбаджа, 1981, табл. 9 , фиг. 11 ; Кузьмин, Mельникова, 1991, табл. 1 , фиг. 19 ; Metzger, 1994, pl. 17, fig. 9 (только); Savage, Yudina, 1999, pl. 5, figs. 15, 16 (только).
Рис. 1.
Объем вида Palmatolepis tatarica Ovnatanova et Gatovsky, 2020 в понимании авторов статьи. На рисунке приведены изображения, заимствованные из работ разных авторов с использованными ими названиями: а–г – Pa. subperlobata subsp. a Helms, 1963: а, б – Helms, 1963, табл. 1 , фиг. 19 , табл. 3 , фиг. 10 : а – вид с оральной стороны; б – вид с аборальной стороны; в, г – Druce, 1976, табл. 58 , фиг. 2b, c: в – вид с оральной стороны, г – вид с аборальной стороны; д–з – Pa. subperlobata helmsi Ovnatanova, 1976: д, е – Овнатанова, 1976, табл. 9 , фиг. 7 а, b (голотип): д – вид с оральной стороны, е – вид с аборальной стороны; ж – Барсков и др., 1987, табл. 4 , фиг. 30 , вид с оральной стороны; з – Çapkinoğlu, 1997, табл. 3 , фиг. 25 , вид с оральной стороны; и–л – Pa. subperlobata Branson et Mehl, 1934: и – Glenister, Klapper, 1966, табл. 92 , фиг. 6 , вид с оральной стороны, к – Халымбаджа, 1981, табл. 9 , фиг. 11 , вид с оральной стороны; л – Metzger, 1994, табл. 17 , фиг. 9 , вид с оральной стороны; м – Pa. lobicornis Schülke, 1995: Klapper et al., 2004, рис. 7.30 , вид с оральной стороны
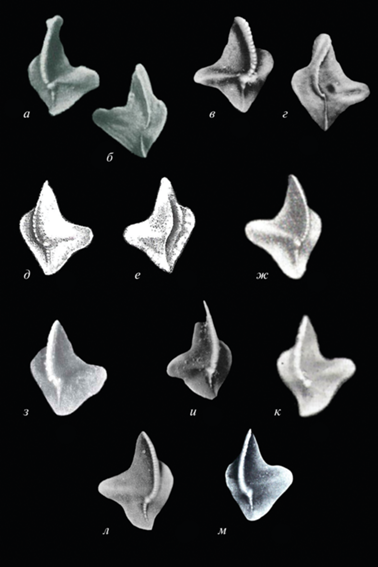
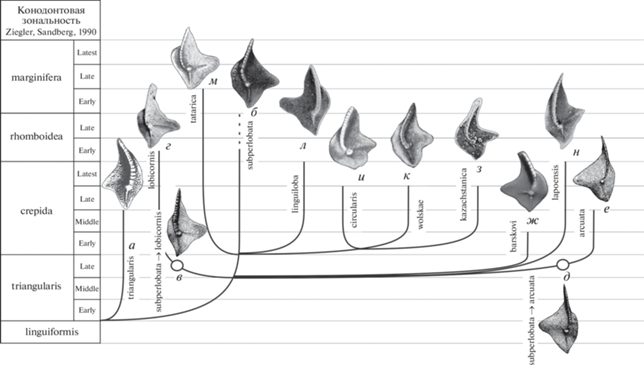
Рис. 2.
Филоморфогенез группы Palmatolepis subperlobata: а – Pa. triangularis: Sannemann, 1955, табл. 24 , фиг. 3 (голотип); б – Pa. subperlobata: Branson, Mehl, 1934, табл. 18 , фиг. 11 (лектотип) (выбран: Ziegler, 1958, с. 64; изображение из: Over, 1997, табл. 10 , фиг. 2); в – Pa. subperlobata Branson et Mehl, переходная форма к Pa. lobicornis Schülke (Schülke, 1995, рис. 26 , фиг. 11 ); г – Pa. lobicornis Schülke: Schülke, 1995, рис. 26 , фиг. 2 (голотип); д – Pa. subperlobata Branson et Mehl, переходная форма к Pa. arcuata Schülke (Schülke, 1995, рис. 26 , фиг. 14 ); е – Pa. arcuata Schülke: Schülke, 1995, рис. 26 , фиг. 5 (голотип); ж – Pa. barskovi Tagarieva et Artyushkova: Тагариева, Артюшкова, 2020, рис. 4 , фиг. 1а (голотип); з – Pa. kazachstanica Kuzmin: Кузьмин, 1992, табл. 1 , фиг. 4 (голотип); и – Pa. circularis Szulczewski: Szulczewski, 1971, табл. 15 , фиг. 6 а (голотип); к – Pa. wolskae Ovnatanova: Овнатанова, 1969, табл. 1 , фиг. 6 (голотип); л – Pa. linguiloba (Dzik): Dzik, 2006, рис. 101 N (голотип); м – Pa. tatarica Ovnatanova et Gatovsky: Овнатанова, 1976, табл. 9 , фиг. 7 (голотип), н – Pa. lapoensis Savage: Savage, 2013, фиг. 11.8 (голотип).
Palmatolepis subperlobata subspecies а: Druce, 1976, с. 170, pl. 58, figs. 2 а–c; 5 a, b.
Palmatolepis subperlobata helmsi: Овнатанова, 1976, с. 113, табл. 9 , фиг. 7 а, б; Ovnatanova, 1979, pl. 1, figs. 29, 30 (только) ; Барсков и др., 1987, с. 29, табл. 4 , фиг. 26 а, б; 28 и 30; Çapkinoğlu, 1997, pl. 3, fig. 25 ; Cтрельченко, Кручек, 2013, табл. 1 , фиг. 13 .
Palmatolepis lobicornis: Klapper et al., 2004, fig. 7.30 ; Çapkinoğlu, 2005, табл. 4 , фиг. 14, 17 (только); Бушнев и др., 2017, рис. 3 , фиг. 23 .
Palmatolepis linguiloba: Фортунатова и др., 2018, рис. 13 , фиг. 6 .
Palmatolepis subperlobata tatarica: Овнатанова, Гатовский, 2020, с. 111.
Голотип – ВНИГНИ, № 40/68, левый Ра-элемент; Татарстан, Приказанская площадь, скв. 116, инт. 1633.6–1630.1 м; фаменский ярус, нижнефаменский подъярус, зона crepida; рис. 1, д, е (Овнатанова, 1976, табл. 9 , фиг. 7a, 7б ).
Описание (рис. 1, а–м; 2, м). Ра-элемент с относительно широкой, гладкой (тонкошагреневой) платформой треугольных очертаний. Передний край наружной стороны платформы приподнятый, выпуклый, имеет округлые очертания. Длинная округлая лопасть расположена на уровне главного бугра и направлена вбок. Задний и передний синусы, оконтуривающие лопасть, неглубокие. Передний синус глубже заднего. Задний синус иногда чрезвычайно слабо выражен (рис. 1, а, б). Задний конец платформы заострен и приподнят. Свободный лист отсутствует. Осевой гребень зубчатый, сигмоидально изогнут, позади главного бугра выражен слабо. Края боковых сторон платформы подходят к осевому гребню на разных расстояниях.
Размеры голотипа в мм: l – 0.7 мм, w – 0.5 мм, где l – длина элемнта, w – ширина элемента с учетом лопасти.
Изменчивость. Проявляется в вариациях длины лопасти от умеренной (рис. 1, з, и, м) до достаточно длинной (рис. 1, а–ж, к), а также в присутствии у некоторых экземпляров небольшого рудимента свободного листа. Меняется в процессе роста и задний конец платформы – от заостренного (рис. 1, д, к) до округлого (рис. 1, ж, з, м).
Сравнение. Сходен с Pa. subperlobata гладкой поверхностью платформы и четко выраженной лопастью, отличается относительно длинной, округлой лопастью и отсутствием свободного листа, тогда как у Pa. subperlobata свободный лист составляет 1/5–1/10 длины платформы.
Распространение. Россия: Волго-Уральский регион, восточный склон Северного Урала, Тимано-Печорская провинция, зоны сrepida – Early marginifera; Белоруссия, петриковский горизонт, зона Late rhomboidea; Германия, Босния: фаменские отложения, зона Cheiloceras, возможно Platyclymenia; Польша, Свентокшиские горы, разрез Яблона, зонa crepida; Турция, формация Айинебуму, зона crepida; США, известняки Вест Рейндж и сланцы Пайлот в Неваде; Канада, формация Лонг Рапидс; Западная Австралия, формация Вирджин Хиллс.
СРАВНЕНИЕ ГОЛОТИПА PA. TATARICA С ГОЛОТИПАМИ PA. LOBICORNIS SCHÜLKE И CONDITOLEPIS LINGUILOBA DZIK
Голотип Pa. tatarica сходен с голотипом вида Pa. lobicornis Schülke треугольными очертаниями платформы, гладкой ее поверхностью, соединением боковых краев платформы с осевым гребнем на разных расстояниях, а также присутствием длинной лопасти. Различия выражаются в отсутствии свободного листа и рога у Pa. tatarica. Очертания лопасти волнистые, но без выпуклости (или рога).
Сходство голотипа Pa. tatarica с голотипом Conditolepis linguiloba Dzik проявляется в треугольных очертаниях платформы, ее гладкой поверхности, длинной округлой лопасти, отсутствии свободного листа, а также в соединении боковых сторон платформы со срединным гребнем на разных расстояниях. Основное различие проявляется в приподнятом, выпуклом, округлом очертании переднего края наружной стороны платформы у Pa. tatarica [этот признак был отмечен еще Хелмсом (Helms, 1963) как его утолщение в виде вздутия или выпуклости]. У голотипа Co. linguiloba этот признак (вздутие, выпуклость) не наблюдается; край передней части наружной стороны платформы пологий, плавно приближается к осевому гребню, не образуя выпуклости. Различия выражаются также в разных соотношениях длины платформы к ее ширине (эта особенность становится очевидной, если мысленно убрать лопасть, искажающую эти параметры): у Pa. tatarica платформа относительно широкая (отношение длины платформы к ее ширине равно 2 : 1), тогда как у Со. linguiloba платформа вытянута в длину (отношение длины к ширине составляет 3 : 1). Положение лопасти у голотипа Pa. tatarica фиксируется на уровне главного бугра, тогда как у Со. linguiloba лопасть направлена к переднему концу платформы. Различия выражаются и в конфигурации переднего синуса, оконтуривающего лопасть: у Pa. tatarica передний синус неглубокий, пологий, тогда как у голотипа Co. linguiloba он глубокий. Задний конец платформы у голотипа Ра. tatarica cлегка заострен и приподнят вверх; угол заостренности его равен 85°, тогда как у Со. linguiloba он составляет 60°.
Таким образом, Pa. tatarica, несмотря на достаточно заметное сходство с Pa. lobicornis и Co. linguiloba, является самостоятельной таксономической единицей.
СОСТАВ И ФИЛОГЕНЕТИЧЕСКИЕ СВЯЗИ ПРЕДСТАВИТЕЛЕЙ ГРУППЫ PALMATOLEPIS SUBPERLOBATA
Вопросы филоморфогенеза Pa. subperlobata и связанных с ним родством других видов, как известно, обсуждаются в литературе, начиная с публикации Циглера (Ziegler, 1962), в которой анализировалась ветвь Pa. subperlobata–Pa. glabra, а также в первой филоморфогенетической схеме всего рода Palmatolepis (Helms, 1963), в которой принималось возникновение Pa. subperlobata s. str. от Pa. triangularis Sannemann, 1955, также показан подвид Pa. (Pand.) subperlobata n. subsp. a и его филогенетические связи.
Первая филогенетическая схема группы Pa. subperlobata, а также другие группы фаменских пальматолепид, рассматривались в монографии Цзы и Циглера (Ji, Ziegler, 1993) на основании коллекций, полученных при изучении разреза Лали (Lali) в Южном Китае. Позднее раннефаменские пальматолепиды, в т.ч. и группа Pa. subperlobata, анализировались И. Шульке (Schülke, 1995), которым были выделены новые отнесенные к этой группе виды: Pa. arcuata Schülke и Pa. lobicornis Schülke.
В предлагаемую нами филоморфогенетическую схему группы Pa. subperlobatа включены: Pa. subperlobata Branson et Mehl, 1934 и возникший от него вид Pa. tatarica Ovnatanova et Gatovsky, 2020 (=Pa. subperlobata helmsi Ovnatanova, 1976), Pa. arcuata Schülke, 1995, Pa. lobicornis Schülke, 1995, Pa. wolskae Ovnatanova, 1969, Pa. circularis Szulczewski, 1971, Pa. linguiloba (Dzik, 2006), Pa. barskovi Tagarieva et Artyushkova, 2020, Pa. kazachstanica Kuzmin, 1992 (=Pa. angulata Johnston et Chatterton, 2001) и Pa. lapoensis Savage, 2013 (=Pa. subperlobata lapoensis Savage, 2013). Для облегчения восприятия морфологических особенностей нами приведены изображения голотипов всех видов этой группы (рис. 2), филогенетические связи которых рассматриваются в настоящей статье в привязке к конодонтовой зональности шкалы Циглера и Ч. Сандберга (Ziegler, Sandberg, 1990).
Вид Pa. tenuipunctata Sannemann, 1955, являющийся предковой формой для группы Pa. glabra, возможно, является самым поздним, терминальным членом группы Pa. subperlobata, так как у него чрезвычайно слабо выражена лопасть, но при этом сохраняется гладкая поверхность платформы – характерный признак этой группы. Авторы настоящей статьи разделяют мнение о родственной связи видов Pa. tenuipunctata и Pa. glabra, высказанное Циглером (Ziegler, 1962) и Хелмсом (Helms, 1963).
Родоначальник рассматриваемой группы, вид Pa. subperlobata (рис. 2, б), произошел от Pa. triangularis Sannemann (рис. 2, а) во время зоны Early triangularis, унаследовав от предка треугольные очертания платформы, хорошо выраженную лопасть и свободный лист. Существенным изменениям подверглась поверхность платформы за счет исчезновения бугорковой скульптуры, в результате чего она стала гладкой. В дальнейшем у большинства видов, произошедших от Pa. subperlobata, исчезает свободный лист.
Во время зоны Late triangularis от Pa. subperlobata возникает Pa. arcuata (рис. 2, е). Он связан с Pa. subperlobata предполагаемой переходной формой, изображенной Шюльке (Schülke, 1995, рис. 26 , фиг. 14 ), она приведена здесь на рис. 2, д. У этой формы есть сходство с предком, выраженное в треугольных очертаниях платформы, гладкой поверхности, четко выраженной лопасти и наличии свободного листа. В процессе эволюционного перехода у Pa. arcuata происходило выпрямление переднего синуса и удлинение лопасти. Вид этот возник во время зоны Late triangularis и завершил развитие в конце зоны Middle crepida (Schülke, 1995).
Во время зоны triangularis (возможно, Late triangularis, когда одновременно от Pa. subperlobata появляются Pa. lobicornis, Pa. arcuata и Pa. barskovi) возникает вид Pa. lapoensis (рис. 2, н), унаследовавший от предка треугольные очертания платформы, свободный лист и почти гладкую поверхность платформы. У него произошло углубление переднего синуса и выпрямление заднего синуса, а также изменились очертания лопасти – она стала заостренно-треугольной. Край передней части наружной стороны платформы становится близким к прямому, в отличие от предка, у которого эта часть платформы округлая. Pa. lapoensis распространен в зонах triangularis и crepida и до сих пор обнаружен только в Таиланде (Savage, 2013).
Вид Pa. lobicornis (рис. 2, г) произошел от Pa. subperlobata во время зоны Late triangularis и связан с предком переходной формой, которая сходна с Pa. subperlobata треугольными очертаниями платформы, гладкой поверхностью и хорошо выраженной лопастью, а также присутствием свободного листа и сигмоидально изогнутого осевого гребня, заканчивающегося вблизи заднего конца платформы. По-видимому, именно такая переходная форма приведена Шюльке на рисунке филогенетической схемы развития группы Pa. subperlobata (Schülke, 1995, рис. 26 , фиг. 4 ). Она приведена нами здесь на рис. 2, в. У этой формы наблюдается небольшая извилистость края внутренней стороны платформы. В процессе развития вида Pa. lobicornis на переднем крае лопасти образуется утолщение в виде небольшого “рога” (рис. 2, г). Стратиграфическое распространение Pa. lobicornis охватывает зону Late triangularis–начало зоны rhomboidea.
От Pa. subperlobata во время зоны Late triangularis происходит Pa. barskovi (рис. 2, ж). От предка он унаследовал треугольные очертания платформы и ее гладкую поверхность, но исчез свободный лист. Отличительная особенность вида проявляется в высокой передней части осевого гребня, состоящей из узких заостренных зубцов, постепенно понижающихся и резко заканчивающихся до достижения ими главного бугра. Отличительная особенность Pa. barskovi, а именно – прерывистость осевого гребня, не была отмечена нами ранее у франских пальматолепид (Ovnatanova, Kononova, 2008), не зафиксирована она также и у фаменских видов. Это позволяет условно cчитать ветвь Pa. barskovi тупиковой. Подобная прерывистость осевого гребня изредка проявляется у некоторых других конодонтов, например, у Polygnathus corpulentus Gagiev et Kononova, у которого осевой гребень в средней части прерывается, становится едва заметным и в виде цепочки мелких бугорков прослеживается вновь ближе к заднему концу платформы (Гагиев и др., 1987). Распространение Palmatolepis barskovi охватывает зоны Late triangularis–Early crepida.
В начале времени зоны Early crepida от Pa. subperlobata возникают Pa. tatarica, Pa. wolskae и, возможно, Pa. linguiloba. Вид Pa. tatarica (рис. 2, м) сохранил от предка гладкую поверхность платформы, ее треугольные очертания и сигмоидальный осевой гребень, но исчез свободный лист, углубился передний синус, и удлинилась лопасть. Передний край наружной стороны платформы стал приподнятым, выпуклым и округлым. Этот вид существовал в диапазоне зон Early crepida–Early marginifera включительно.
Вид Pa. linguiloba (рис. 2, л) унаследовал от Pa. subperlobata треугольные очертания платформы и ее гладкую поверхность, однако произошло сокращение длины свободного листа, вплоть до его полного исчезновения, удлинение лопасти и углубление переднего синуса (угол, образованный краями синуса, становится равным 90°). Нельзя исключить происхождение Pa. linguiloba и от Pa. tatarica за счет удлинения платформы, приведшего к исчезновению выпуклости и приподнятости переднего края наружной стороны платформы. Диапазон распространения вида охватывает зону сrepida.
Вид Pa. wolskae (рис. 2, к) ответвился от Pa. subperlobata и сохранил гладкую поверхность платформы и небольшую лопасть, но абрис платформы стал неправильно-овальным, исчез свободный лист, передний и задний синусы стали более глубокими. Задний конец платформы приобрел языковидные очертания. Распространен в зонах Early crepida–Late? crepida.
Во время зоны Early crepida от Pa. wolskae возник Pa. circularis (рис. 2, и), также с гладкой поверхностью платформы, небольшой лопастью и без свободного листа. Но у него платформа приобретает округлые очертания, сокращаются размеры лопасти, заостряется ее окончание. Лопасть очерчена относительно глубокими передним и задним синусами. Позади заднего синуса увеличивается степень выпуклости края платформы. Осевой гребень заканчивается, достигнув главного бугра, а задний гребень исчезает. Распространен в зонах Early–Late–Latest? crepida.
Во время зоны Early crepida от Pa. wolskae возникает и вид Pa. kazachstanica (рис. 2, з). От предка он унаследовал гладкую поверхность платформы, языковидный контур заднего конца, отсутствие свободного листа. Однако произошло удлинение лопасти, углубление переднего и заднего синусов; края переднего синуса сходятся под углом 90°. Этот вид встречается в интервале зон Early crepida–Late crepida. К Pa. kazachstanica можно отнести экземпляры, определенные под различными названиями: Pa. aff. circularis (Sandberg, Ziegler, 1973, табл. 1 , фиг. 1–12 ), Pa. wolskae [Klapper, Lane, 1985, рис. 15.1, 15 . 2?, 15.4, 15.5; Johnston, Chatterton, 1991, табл. 1 , фиг. 22 ; Metzger, 1994, рис. 7.8, 7.9, 7.13 (только)]. Позднее Д. Джонстон и Б. Чаттертон описали аналогичные формы под новым названием Palmatolepis angulata из разрезов Канады (Johnston, Chatterton, 2001, с. 22, 23; табл. 5 , фиг. 15–19 ; табл. 6 , фиг. 4, 5 ), так как статья А.В. Кузьмина (1992) им была неизвестна. Авторы этого вида предполагали его возникновение от Pa. wolskae или форм близких к нему за счет удлинения лопасти. Вид Pa. kazachstaniсa описан из фаменских отложений (зона crepida) Атасуйского рудного района в Центральном Казахстане (Кузьмин, 1992). В Канаде вид Pa. angulata известен из пачки Морро формации Пализер (Paliser), охватывающей верхнюю часть зоны Lower crepida и нижнюю часть зоны Upper crepida (Johnston, Chatterton, 2001, с. 23). В США Pa. angulata обнаружен в штате Невада в известняках Вест Рэндж в Бактрийских горах под названием Pa. wolskaе (Metzger, 1994). Мы считаем, что все эти экземпляры должны быть отнесены к Pa. kazachstaniсa, как опубликованному раньше, чем Pa. angulata, и, поэтому, имеющему приоритет.
ЗАКЛЮЧЕНИЕ
Сравнение характеристик голотипов видов Pa. tatarica Ovnatanova et Gatovsky, 2020, Pa. lobicornis Schülke, 1995 и Pa. linguiloba (Dzik, 2006) позволило доказать, что вид Pa. tatarica является самостоятельной систематической единицей. Намечены филогенетические связи между видами группы Pa. subperlobata, куда входит и вид Pa. tatarica (рис. 2).
Состав группы Pa. subperlobata расширен: наряду с подвидом Pa. subperlobata subsp. a Helms, 1963 (= Pa. tatarica) и видами Pa. wolskae и Pa. circularis, ранее рассматриваемыми в составе этой группы (Ji, Ziegler, 1993), в группу Pa. subperlobata включены также виды Pa. linguiloba, Pa. kazachstanica (=Pa. angulata), Pa. arcuata, Pa. lobicornis, Pa. lapoensis, наряду с Pa. barskovi Tagarieva et Artyushkova, 2020, установленным недавно в фаменских отложениях Южного Урала (Тагариева, Артюшкова, 2020).
Список литературы
Барсков И.С., Алексеев А.С., Кононова Л.И., Мигдисова А.В. Определитель конодонтов верхнего девона и карбона. М.: Изд-во Моск. ун-та, 1987. 144 с.
Бушнев Д.А., Плотицын А.Н., Груздев Д.А., Бурдельная Н.С. Органическое вещество фаменских отложений южной части гряды Чернышева (разрез на р. Изъяель) // Нефтегазовая геология. Теория и практика. 2017. Т. 12. № 3. С. 1–17.
Гагиев М.Х., Кононова Л.И., Пазухин В.Н. Конодонты // Фауна и биостратиграфия пограничных отложений девона и карбона Берчогура (Мугоджары). М.: Наука, 1987. С. 91–97.
Деулин Ю.В. Конодонты верхнедевонских отложений севера Тимано-Печорской провинции и их стратиграфическое значение. Автореф. дисс. … канд. геол.-мин. наук. Казань, 2000. 22 с.
Кононова Л.И. Значение конодонтов для стратиграфии фаменских и турнейских отложений Урала // Конодонты Урала и их стратиграфическое значение. Свердловск: УНЦ АН СССР, 1979. С. 72–93.
Кузьмин А.В. Использование конодонтов для расчленения фаменских отложений Атасуйского рудного района (Центральный Казахстан) // Изв. АH CCCP. Сер. геол. 1992. № 8. С. 30–40.
Кузьмин А.В., Мельникова Л.И. Расчленение по конодонтам франских и нижнефаменских отложений южной части Хорейверской впадины (Тимано-Печорская провинция) // Бюлл. МОИП. Отд. геол. 1991. Т. 66. № 3. С. 62–72.
Международный кодекс зоологической номенклатуры. Изд. 4-е. СПб.: ЗИН РАН, 2000. 221 с.
Наседкина В.А., Плотникова Н.П. О верхнедевонских конодонтах на восточном склоне Северного Урала // Конодонты Урала и их стратиграфическое значение. Свердловск: УНЦ АН СССР, 1979. С. 52–68.
Овнатанова Н.С. Новые верхнедевонские конодонты центральных районов Русской платформы и Тимана // Тр. ВНИГНИ. 1969. Вып. 93. С. 139–141.
Овнатанова Н.С. Новые позднедевонские конодонты Русской платформы // Палеонтол. журн. 1976. № 2. С. 106–115.
Овнатанова Н.С., Гатовский Ю.А. Palmatolepis subperlobata tatarica nom. nov. – новое подвидовое название для фаменского (верхний девон) подвида Palmatolepis subperlobata helmsi Ovnatanova, 1976 (Conodonta) // Палеонтол. журн. 2020. № 3. С. 111.
Плотицын А.Н., Мизенс А.Г., Безносова Т.М. К стратиграфии пограничных фран–фаменских отложений р. Шаръю (гряда Чернышева) // Вестн. Ин-та геол. Коми НЦ УрО РАН. 2015. № 11. С. 13–18.
Стрельченко Т.В., Кручек С.А. Стратиграфия нижнефаменских отложений Припятского прогиба (Белоруссия) по конодонтам // Стратигр. Геол. корреляция. 2013. Т. 21. № 2. С. 22–42.
Тагариева Р.Ч., Артюшкова О.В. Конодонтовая зональность нижнефаменских отложений карбонатных разрезов западного склона Южного Урала // Бюлл. МОИП. Отд. геол. 2020. Т. 95. № 2. С. 26–41.
Фортунатова Н.К., Зайцева Е.Л., Кононова Л.И. и др. Литолого-фациальная и биостратиграфическая характеристика верхнедевонских отложений опорной скважины 1 Мелекесская (Мелекесская впадина, Волго-Уральская область) // Бюлл. МОИП. Отд. геол. 2018. Т. 93. № 5–6. С. 3–49.
Халымбаджа В.Г. Конодонты верхнего девона востока Русской платформы, Южного Тимана, Полярного Урала и их стратиграфическое значение. Казань: Изд‑во Казанск. ун-та, 1981. 216 с.
Халымбаджа В.Г., Чернышева Н.Г. Конодонты верхнедевонских отложений центральной части Волго-Уральской области и их стратиграфическое значение // Стратиграфия и палеонтология палеозоя востока Русской платформы. Казань: Изд-во Казанск. ун-та, 1978. С. 3–42.
Becker R.T., Hartenfels S. Species list of Palmatolepis // Subcommission on Devonian Stratigraphy. Newsletter. 2016. № 31. P. 35–44.
Branson E.B., Mehl M.G. Conodonts from the Grassy Creek Shale of Missouri // Missouri Univ. Studies. 1934. V. 8. № 3. P. 171–259.
Çapkinoğlu Ş. Conodont fauna and biostratigraphy of the Famennian of Büyükada, İstanbul, Northwestern Turkey // Boll. Soc. Paleontol. Ital. 1997. V. 35. № 2. P. 165–185.
Çapkinoğlu Ş. Upper Devonian (Upper Frasnian – Lower Famennian) conodont biostratigraphy of the Ayineburnu Formation (Istanbul Zone, NW Turkey) // Geol. Carpathica. 2005. V. 56. № 3. P. 223–236.
Druce E.C. Conodont Biostratigraphy of the Upper Devonian reef complexes of the Canning Basin, western Australia // Bull. Austral. Bur. Miner. Res. Geol. Geophys. 1976. № 158. 303 p.
Dzik J. The Famennian “Golgen age” of conodonts and ammonoids in the Polish part of the Variscan sea // Palaeontol. Pol. 2006. № 63. 359 p.
Glenister B.F., Klapper G. Upper Devonian conodonts from the Canning Basin, western Australia // J. Paleontol. 1966. V. 40. № 4. P. 777–842.
Helms J. Zur “Phylogenese” und Taxionomie von Palmatolepis (Conodontida, Oberdevon) // Geologie. 1963. Bd 12. № 4. S. 449–485.
Ji Q., Ziegler W. The Lali section: An excellent reference section for Upper Devonian in South China // Cour. Forsch.-Inst. Senckenb. 1993. № 157. 183 p.
Johnston D.I., Chatterton B.D.E. Famennian conodont biostratigraphy of the Palliser Formation, Rocky Mountains, Alberta and British Columbia // Bull. Geol. Surv. Canada. 1991. № 417. P. 163–183.
Johnston D.I., Chatterton B.D.E. Upper Devonian (Famennian) conodonts from the Palliser Formation and Wabamun Group, Alberta and British Columbia, Canada // Paleontogr. Canad. 2001. № 19. 154 p.
Klapper G., Lane H.R. Upper Devonian (Frasnian) conodonts of the Polygnathus biofacies, N.W.T., Canada // J. Paleontol. 1985. V. 59. № 4. P. 904–951.
Klapper G., Uyeno T.T., Armstrong D.K., Telford P.G. Conodonts of the Williams Island and Long Rapids Formations (Upper Devonian, Frasnian-Famennian) of the Onakawana B drillhole, Moose River Basin, Northern Ontario, with a revision of Lower Famennian species // J. Paleontol. 2004. V. 78. № 2. P. 371–387.
Metzger R.A. Multielement reconstuctions of Palmatolepis and Polygnathus (Upper Devonian, Famennian) from the Canning Basin, Australia, and Bactrian Mountain, Nevada // J. Palentol. 1994. V. 68. № 3. P. 617–647.
Müller K.J. Zur Kenntnis der Conodonten-Fauna des europäischen Devons, 1. Die Gattung Palmatolepis // Abh. Senckenb. Naturforsch. Ges. 1956. № 494. 70 s.
Over D.J. Conodont biostratigraphy of the Java Formation (Upper Devonian) and the Frasnian-Famennian boundary in western N.Y. State // Geol. Soc. Amer. Spec. Pap. 1997. № 321. P. 161–177.
Ovnatanova N.S. Frasnian–Famennian conodonts of the Volgo-Ural Region // Serv. Geol. Belg. Prof. Pap. 1979. № 161. P. 62–70.
Ovnatanova N.S., Kononova L.I. Frasnian conodonts from Eastern Russian Platform // Paleontol. J. 2008. V. 42. № 10. P. 997–1166.
Sandberg C.A., Ziegler W. Refinement of standard Upper Devonian conodont zonation, based on sections in Nevada and West Germany // Geol. Palaeontol. 1973. V. 7. P. 97–122.
Sannemann D. Beitrag zur Untergliederung des Oberdevons nach Conodonten // N. Jb. Geol. Paläontol. Abh. 1955. Bd 100. № 8. S. 324–331.
Savage N.M. Late Devonian conodonts from Northwestern Thailand. Eugene: Trinity Press, 2013. 48 p.
Savage N.M., Yudina A.B. Late Devonian Syv`yu River Section, Timan Pechora basin, Northwestern Russia // Boll. Soc. Paleontol. Ital. 1999. V. 37. № 2–3. P. 361–373.
Schülke I. Evolutive Prozesse bei Palmatolepis in der frühen Famenne-Stufe (Conodonta, Ober-Devon) // Göttingen Arb. Geol. Paläontol. 1995. № 67. 108 s.
Silvério G.G., Valenzuelo-Ríos J.I., Liao J. Upper Frasnian and lower Famennian (Upper Devonian) conodonts of the Compte section (Spanish Central Pyrenees) // Spanish J. Palaeontol. 2021. V. 36. № 2. P. 205–220.
Spaletta C., Perri M.C., Over D.J., Corradini C. Famennian (Upper Devonian) conodont zonation: revised global standard // Bull. Geosci. 2017. V. 92. № 1. P. 31–57.
Spasov H., Filipović I. The conodont fauna of the older and younger Palaeozoic in southeastern and northwestern Bosnia // Geol. Glasn. 1966. V. 11. P. 33–54 (In Bulgarian).
Szulczewski M. Upper Devonian conodonts, stratigraphy and facial development in the Holy Cross Mts. // Acta Geol. Pol. 1971. V. 21. № 1. 129 p.
Wang C.Y. Devonian conodonts of Guangxi // Mem. Nanjing Inst. Geol. Paleontol. 1989. V. 11. P. 51–96 (in Chinese).
Wolska Z. Upper Devonian conodonts from the southwest region of the Holy Cross Mountains, Poland // Acta Palaeontol. Pol. 1967. V. 12. № 4. P. 363–456.
Ziegler W. Conodontenfeinstratigraphische Untersuchungen an der Grenze Mittledevon/Oberdevon und in der Adorfstufe // Hess. L.-Amt. Bodenforsch. Notizbl. 1958. № 87. S. 7–77.
Ziegler W. Taxionomie und Phylogenie Oberdevonischer Conodonten und ihre stratigraphishe Bedeutung // Hess. L.-Amt. Bodenforsch. Abh. 1962. № 38. 166 S.
Ziegler W. Eine neue Conodonten-Fauna aus dem höchsten Oberdevon // Forstschr. Geol. Rheinland Westfalen. 1969. № 17. S. 343–360.
Ziegler W. Conodont Stratigraphy of the European Devonian // Mem. Geol. Soc. Amer. 1971. № 127. P. 227–284.
Ziegler W., Sandberg C.A. The Late Devonian standard conodont zonation // Cour. Forsch.-Inst. Senckenb. 1990. № 121. 115 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал