

Палеонтологический журнал, 2023, № 1, стр. 42-48
Обзор триасовых ручейников (Insecta: Trichoptera) с описанием новых видов из триаса Киргизии (местонахождение Джайляучо)
И. Д. Сукачева a, *, Н. Д. Синиченкова a, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: lab@palaeoentomolog.ru
** E-mail: nina_sin@mail.ru
Поступила в редакцию 26.04.2022
После доработки 12.09.2022
Принята к публикации 12.09.2022
- EDN: FKXPXP
- DOI: 10.31857/S0031031X23010130
Аннотация
Составлен обзор всех триасовых ручейников и обсуждается их стратиграфическое и географическое распространение. Описаны новые виды ручейников семейств Cladochoristidae: Cladochorista curta sp. nov. и Cladochoristella sola sp. nov., и Prorhyacophilidae: Prorhyacophila rara sp. nov. и P. batkenica sp. nov. из местонахождения Джайляучо в Киргизии (средний–верхний триас, мадыгенская свита).
Триасовые местонахождения насекомых обнаружены на всех континентах, но, за редким исключением, остатки насекомых там немногочисленные и часто не лучшей сохранности. В результате триасовый период пока остается самым плохо изученным стратиграфическим интервалом мезозоя, в отличие от юрского и мелового с их многочисленными и богатыми местонахождениями, особенно, в Евразии.
В настоящее время триасовые ручейники известны из Европы, Австралии и Северной Америки. Они представлены семействами Cladochoristidae Riek, 1953, Prorhyacophilidae Riek, 1955, Necrotauliidae Handlirsch, 1906 и Philopotamidae Stephens, 1836 (Wieland, 1926; Riek, 1953, 1955; Сукачева, 1973, 1982; Eskov, Sukatsheva, 1997; Ivanov, Sukatsheva, 2002). Все триасовые виды, за редкими исключениями, описаны по единичным остаткам изолированных крыльев. В нижнетриасовых отложениях ручейники до сих пор не найдены, они встречаются, начиная со среднего триаса, хотя некоторые из них известны из перми. В настоящей статье описываются четыре новых вида из знаменитого местонахождения Джайляучо (Киргизия).
Возможно, что именно в триасе одно из семейств ручейников могло дать начало современным формам. Например, Р. Вильман (Willmann, 1989), учитывая общую редукцию жилкования крыльев, считал, что именно Cladochoristidae могли быть предками современных Trichoptera. Характерная для Cladochoristidae необычная многоветвистая Sc, скорее всего, вторичный признак, встречающийся и у современных ручейников.
Изучение морфологии пермских представителей ручейников и анализ их геологической истории дало возможность В.Г. Новокшонову (1997) предположить, что именно в триасе произошли переход личинок ручейников к обитанию в воде и разделение отряда на современные подотряды. Однако удостовериться в этом окончательно очень трудно, так как триас достаточно беден остатками ручейников. Их относительное количество даже в богатых местонахождениях не достигает среднего пермского уровня. Лишь со второй половины юры ручейники начинают встречаться достаточно часто и представлены в сборах и коллекциях несколькими семействами из обоих современных подотрядов (Сукачева, 1980, 1982).
Семейство Cladochoristidae впервые в палеонтологической летописи появляется в поздней перми. Cladochorista belmontensis Tillyard, 1926 описана из верхнепермских отложений Нового Южного Уэльса в Австралии (Tillyard, 1926). Два экз. C. issadica Sukatsheva et Aristov, 2013 найдены в верхней перми местонахождения Исады в Вологодской области России. Еще один экземпляр этого рода, не определенный до вида, найден в верхней перми на р. Муи провинции Наталь в Южной Африке (Van Dijk, Geerstsema, 1999). Один вид другого рода этого семейства, Cladochoristella ryzkovae Sukatsheva et Aristov, 2013, описан по единственному отпечатку из верхнепермских отложений местонахождения Балымотиха во Владимирской области России (Sukatsheva, Aristov, 2013).
Оба указанных рода преодолевают пермо-триасовую границу (Sukatsheva, Aristov, 2013). В знаменитом местонахождении Джайляучо в Киргизии в отложениях мадыгенской свиты, датируемых средним–поздним триасом (ладин–карний), найдено по одному экземпляру двух видов: Cladochorista multivenosa Sukatsheva, 1973 (Сукачева, 1973) и C. curta sp. nov. Большим числом экземпляров (37) представлена в верхнем триасе Австралии (Маунт Кросби) только Cladochoristella bryani Riek, 1955 (Riek, 1955). Ниже по одному экземпляру описывается новый вид C. sola sp. nov. из Джайляучо.
Недавно представители Cladochoristidae были описаны из нижнего мела Бурятии (Melnitsky et al., 2020), где впервые приведены морфологические особенности строения груди и брюшка. Из юры это семейство неизвестно. Важно, что у самок был развит телескопический яйцеклад, представленный втяжными сегментами брюшка. Схожие морфологические структуры встречаются у представителей современных семейств Rhyacophilidae Stephens, 1836, Hydrobiosidae Ulmer, 1907, Glossosomatidae Wallengren, 1891 и Philopotamidae Stephens, 1836.
Семейство Prorhyacophilidae, включающее в себя единственный род Prorhyacophila, также переходит из перми в триас, но встречается редко: все виды описаны по единичным экземплярам. Из верхнепермских отложений Балымотихи во Владимирской области России описан P. rasnitsyni Sukatsheva et Aristov, 2013 (Sukatsheva, Aristov, 2013). В триасе семейство становится более разнообразным, всего описано пять видов. Из верхнего триаса угленосного бассейна Ипсвич в Квинсленде в формации Маунт Кросби, Австралия, описаны P. collivery Riek, 1955 (Riek, 1955) и, возможно, Eocorona iani (Tindale, 1980) (Сукачева, 1982, с. 9). Остальные виды найдены в Джайляучо: один вид описан ранее: Prorhyacophila furcata Sukatsheva, 1973, два новых – P. rara sp. nov. и P. batkenica sp. nov. – описываются ниже.
Вблизи Джайляучо в юрском местонахождении Мадыген найден один экз. самого массового вымершего мезозойского семейства Necrotauliidae Handlirsch, 1906 – Necrotaulius proximus Sukatsheva, 1973 (Сукачева, 1973). Это семейство в юрских ориктоценозах начинает доминировать среди ручейников, хотя первые находки известны из триаса (карний) Северной Америки (Cow Branch Formation) (Pierwola, Grimaldi, 2022). Кроме того, Necrotauliidae описаны из верхов триаса (рэт) Англии (Kelly et al., 2018).
Из Джайляучо было описано переднее крыло Prophilopotamus asiaticus Sukatsheva, 1973, отнесенное к современному семейству Philopotamidae (Сукачева, 1973). Эта реликтовая группа с небольшим числом родов распространена всесветно, но главным образом, в Северном полушарии. Древность и примитивность филопотамид отмечали Т. Коккерель (Cockerell, 1916) и А.В. Мартынов (1934), а находка триасового представителя семейства подтверждает это мнение.
Данная работа посвящена результатам изучения новых сборов ископаемых ручейников из лагерштетта Джайляучо в 2006, 2007 и 2009 гг. (Киргизия, Ошская обл., Баткенский р-н). Местонахождение было открыто в 1933 г. геологом Е.А. Кочневым (Кочнев, 1934). Насекомые – наиболее многочисленная группа ископаемых в мадыгенской свите, а в этом местонахождении в особенности. Подробная характеристика местонахождения дана в отдельных работах (Шаров, 1968; Щербаков, 2008; Shcherbakov, 2008; Fischer et al., 2011; Voigt et al., 2017).
В настоящее время в Джайляучо собрано более 25 тыс. экз. насекомых из 20 отрядов и 170 семейств, описано более 500 видов. Насекомые, большинство которых представлено изолированными крыльями, встречены во всех обнажениях мадыгенской свиты, но более 80% – в урочище Джайляучо. В фауне мадыгенской свиты доминируют жуки (Coleoptera), равнокрылые (Homoptera) и таракановые (Blattodea) (Voigt et al., 2017). Настоящие амфибиотические насекомые, такие как веснянки, стрекозы и ручейники, очень редки (в целом описано около 50 видов, большая их часть принадлежит стрекозам) (Сукачева, 1973; Притыкина, 1981; Синиченкова, 1987; Щербаков, 2008). Поденки в этом местонахождении не найдены. Единственный фрагмент нимфы (экз. ПИН, № 2069/40), определенный ранее как близкий к Mesobaetis Brauer, Redtenbacher et Ganglbauer, 1889 семейства Siplohlonuridae (Sinitshenkova, 2000), и упомянутый в некоторых работах (Shcherbakov, 2008; Voigt еt al., 2017), в действительности поденкой не является. Мадыгенская свита сформировалась в озерном бассейне с хорошо аэрированным дном в нескольких сотнях километров от ближайшей морской береговой линии. Теплый семиаридный климат и богатая растительность создавали весьма благоприятные условия для развития разнообразной фауны фитофагов (Coleoptera, Hemiptera, Miomoptera, Orthoptera, Phasmatodea) и сапрофагов (Blattodea, Mecoptera, Coleoptera) (Shcherbakov, 2008; Fischer et al., 2011; Voigt et al., 2017).
Разнообразие насекомых в мадыгенской свите необыкновенно высоко по сравнению с другими триасовыми и даже пермскими местонахождениями, и характеризуется большим числом первых и последних находок (в 40% всех семейств), что является показателем типичного лагерштетта (Shcherbakov, 2008). При этом за все время сборов (1933–2009 гг.) найдено всего 55 экз. ручейников, среди них большинство представлено фрагментами передних крыльев и тел.
Работа поддержана грантом РНФ, проект № 21-14-00284, и выполнена на базе Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН).
Авторы сердечно благодарят рецензентов В.Д. Иванова и А.С. Башкуева за глубокий, всесторонний анализ статьи, способствовавший ее несомненному улучшению, а также М.Я. Попову за техническую помощь.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД TRICHOPTERA
СЕМЕЙСТВО CLADOCHORISTIDAE RIEK, 1953
Род Cladochorista Tillyard, 1926
Cladochorista curta Sukatsheva et Sinitshenkova, sp. nov.
Табл. VI , фиг. 1 (см. вклейку)
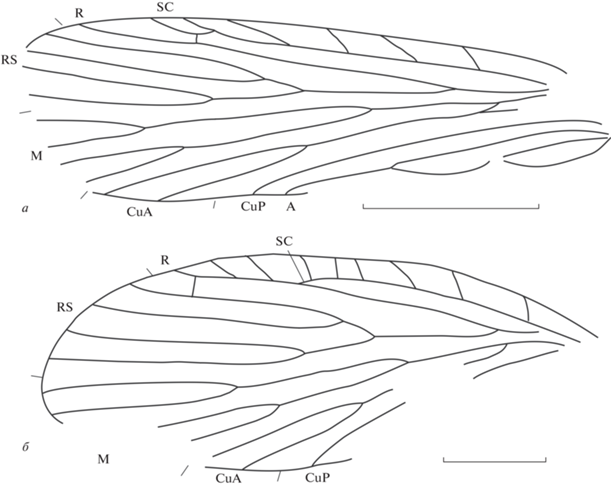
Рис. 1.
Представители семейства Cladochoristidae из местонахождения Джайляучо, средний–верхний триас Киргизии: а – Cladochorista curta sp. nov., голотип ПИН, № 5330/92, длина масштабной линейки 2 мм; б – Cladochoristella sola sp. nov., голотип ПИН, № 5330/84, длина масштабной линейки 1 мм.
Название вида от curtus лат. – неполный.
Голотип – ПИН, № 5330/92, прямой отпечаток переднего крыла; Киргизия, Ошская обл., Баткенский р-н, местонахождение Джайляучо; средний–верхний триас, ладин–карний, мадыгенская свита.
Описание (рис. 1, а). Крыло довольно узкое, длина больше ширины в 3.2 раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Передний край слегка выпуклый, вершинная часть крыла и торнус сохранились не полностью. SC длинная, с пятью косыми ветвями и небольшим вершинным развилком; оканчивается в самом начале четвертой четверти длины крыла. R прямой, без вершинного развилка, оканчивается в середине четвертой четверти длины крыла. Костальное и субкостальное поля широкие. Костальное поле на уровне середины длины крыла в 1.2 раза шире субкостального. Имеется небольшая прямая поперечная жилка sc–r в вершинной части крыла. RS и M четырехветвистые. Ствол RS в 3.7 раза длиннее ствола RS1 + 2 и в 1.6 раза – ствола развилка F3 + 4. Ствол развилка F1 в 2.3 раза короче ствола развилка F2. Ствол развилка F4 чуть короче ствола F3. Соответственно, вершины развилков F1 и F3 расположены проксимальнее вершин развилков F2 и F4. Ячеи DC, MC и TC открытые. Развилки F3 и F4 очень короткие. Разветвление M–CuA расположено заметно проксимальнее разветвления R–RS. Развилок CuA длинный, мощный, начинается немного дистальнее середины длины крыла. CuP и A1 впадают в задний край крыла в разных точках несколько дистальнее середины длины крыла в начале его третьей четверти, довольно далеко друг от друга. Имеется Y-образная жилка с почти равными плечами. Жилка A1 в 1.5 раза длиннее слабоизогнутой A2. A3 короткая. Окраска крыла отсутствует.
Размеры в мм: длина крыла 6.8, ширина 2.2.
Сравнение. Описываемый вид отличается от остальных четырех известных видов рода Cladochorista очень большой разницей в длинах стволов развилков F1 и F2 (в 2.3 раза).
Материал. Голотип.
Род Cladochoristella Riek, 1955
Cladochoristella sola Sukatsheva et Sinitshenkova, sp. nov.
Табл. VI , фиг. 2
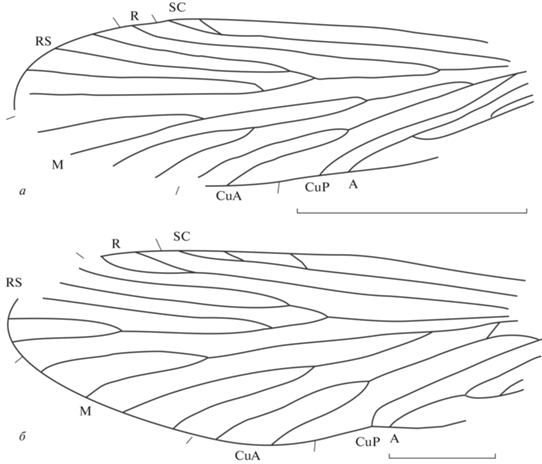
Рис. 2.
Представители семейства Prorhyacophilidae из местонахождения Джайляучо, средний–верхний триас Киргизии: а – Prorhyacophila rara sp. nov., голотип ПИН, № 5330/83, длина масштабной линейки 1 мм; б – P. batkenica sp. nov., голотип ПИН, № 5330/200, длина масштабной линейки 2 мм.
Название вида от solus лат. – одиночный.
Голотип – ПИН, № 5330/84, прямой и обратный отпечатки переднего крыла; Киргизия, Джайляучо; средний–верхний триас, мадыгенская свита.
Описание (рис. 1, б). Крыло довольно широкое, длина больше ширины в 2.6 раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Передний край крыла слегка выпуклый, вершинная часть и торнус сохранились не полностью. SC средней длины, впадает в радиус, с шестью косыми ветвями, оканчивается в середине третьей четверти длины крыла. R прямой, с двумя ветвями на вершине, оканчивается у конца третьей четверти длины крыла. RS и M четырехветвистые. Костальное и субкостальное поля в своем основании почти одинаково широкие, но SC у своей вершины, изгибаясь, приближается к R, и субкостальное поле становится вдвое уже. Между вершинами R и RS1 имеется прямая поперечная жилка r–rs1. Развилки F1 и F2 длинные, но ствол развилка F1 в 1.7 раза короче ствола развилка F2. Развилки F3 и F4 намного короче, ствол F4 в 1.1 раза короче ствола F3. Ячеи DC, MC и TC открытые. M–CuA разветвляется почти на одном уровне с разветвлением R–RS. Развилок F5 мощный, длинный, начинается на середине длины крыла. Y-образная жилка сохранилась плохо. Анальное поле тоже не сохранилось. Окраска крыла темная.
Размеры в мм: длина крыла 6.0, ширина крыла 3.0.
Сравнение. Описываемый вид отличается от двух других видов рода Cladochoristella иной разницей в длинах стволов развилков F1 и F2 (ствол F1 в 1.7 раза длиннее ствола F2). У C. bryani Riek, 1955 разница составляет в 1.5 раза, а у C. ryzkovae Sukatsheva et Aristov, 2013 – 1.4.
Материал. Голотип.
СЕМЕЙСТВО PRORHYACOPHILIDAE RIEK, 1955
Род Prorhyacophila Riek, 1955
Prorhyacophila rara Sukatsheva et Sinitshenkova, sp. nov.
Табл. VI , фиг. 3
Название вида от rarus лат. – редкий.
Голотип – ПИН, № 5330/83, прямой и обратный отпечатки переднего крыла; Киргизия, Джайляучо; средний–верхний триас, мадыгенская свита.
Описание (рис. 2, а). Крыло довольно широкое, длина больше ширины в 2.2 раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Передний край прямой, вершинная часть и торнус сохранились не полностью. SC и R длинные, прямые. SC с небольшим развилком на вершине, оканчивается на границе третьей и четвертой четвертей длины крыла. R простой, оканчивается в начале четвертой четверти длины крыла. Костальное и субкостальное поля одинаковой ширины почти по всей длине. RS и M четырехветвистые. Ствол RS в 4.3 раза больше ствола RS1 + 2 и в 2.4 раза больше ствола RS3 + 4, а ствол развилка F2 в 2.5 раза больше ствола F1. Ячеи DC, MC и TC открытые. Ствол M в 1.2 раза больше ствола развилка F4 и в 1.4 раза меньше ствола F3. Y-образная жилка сохранилась плохо. Жилки CuP и A1 оканчиваются во второй четверти длины крыла в разных точках далеко друг от друга. Жилка A1 в 1.5 раза длиннее A2, A3 короткая.
Размеры в мм: длина крыла 5.5, ширина крыла 2.5.
Сравнение. Описываемый вид по наличию развилка только на SC близок к P. collivery Riek, 1955, отличаясь гораздо более длинным развилком F4 по сравнению с F3. У P. collivery оба развилка одинаковой длины (короткие). Кроме того, у него ствол F1 в 1.5 раза длиннее ствола F2, в то время как у описываемого вида он в 2.5 раза короче ствола F2.
Материал. Голотип.
Prorhyacophila batkenica Sukatsheva et Sinitshenkova, sp. nov.
Табл. VI , фиг. 4
Название вида от Баткенского района Киргизии.
Голотип – ПИН, № 5330/200, прямой и обратный отпечатки переднего крыла; Киргизия, Джайляучо; средний–верхний триас, мадыгенская свита.
Описание (рис. 2, б). Переднее крыло довольно узкое, длина больше ширины в три раза. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Передний край крыла прямой, вершина немного вытянута, торнус округлый. SC и R длинные, прямые; SC с коротким развилком на вершине, впадает в C на середине третьей четверти крыла. R с длинным узким вершинным развилком, оканчивается в начале четвертой четверти длины крыла. Костальное и субкостальное поля одинаково широкие в основании и в средней части, но к вершине сужаются. В костальном поле на уровне вершины развилка RS имеется косая длинная поперечная жилка c‒sc. RS пятиветвистый, ствол RS в 4.7 раза длиннее ствола RS1 + 2 и в 2.3 раза длиннее ствола RS3 + 4. Ствол развилка F2 в 2 раза длиннее ствола развилка F1. На RS3 + 4 имеется средней величины развилок, ствол которого почти равен по длине самому развилку. Ячеи DC, MC и TC открытые. Ствол M в 1.8 раза короче ствола развилка F3 и в 1.1 раза длиннее ствола развилка F4. Y-образная жилка не равносторонняя, нижнее плечо вдвое длиннее верхнего. Жилки CuP и A1 впадают в задний край крыла в разных точках далеко друг от друга. Жилка A1 в 1.6 раза длиннее A2, A3 короткая. CuP и A1 оканчиваются в начале второй четверти длины крыла.
Размеры в мм: длина крыла 11.0, ширина крыла 4.0.
Сравнение. Новый вид отличается от остальных четырех видов рода Prorhyacophila наличием дополнительного развилка на RS3 + 4 переднего крыла и имеет другие соотношения длин стволов всех апикальных развилков. P. batkenica sp. nov. сходен с P. furcata Sukatsheva, 1973, описанным также из местонахождения Джайляучо, по наличию вершинных развилков на R и SC.
Материал. Голотип.
Список литературы
Кочнев Е.А. К вопросу изучения юрских угленосных отложений Ферганы // Недра Средней Азии. 1934. № 5–6. С. 136–158.
Мартынов А.В. Ручейники. Trichoptera Annulipalpia, 1. Л.: Изд-во Зоол. ин-та АН СССР, 1934. 343 с. (Определители по фауне СССР. Т. 13).
Новокшонов В.Г. Ранняя эволюция скорпионниц. М.: Наука, 1997. 138 с.
Притыкина Л.Н. Новые триасовые стрекозы Средней Азии // Вишнякова В.Н., Длусский Г.М., Притыкина Л.Н. Новые ископаемые насекомые с территории СССР. М.: Наука, 1981. С. 5–42 (Тр. Палеонтол. ин-та АН СССР. Т. 183).
Синиченкова Н.Д. Историческое развитие веснянок. М.: Наука, 1987. 143 с. (Тр. Палеонтол. ин-та АН СССР. Т. 221).
Сукачева И.Д. Новые ручейники (Trichoptera) из мезозоя Средней Азии // Палеонтол. журн. 1973. № 5. С. 100–107.
Сукачева И.Д. Отряд Phryganeida Latreille, 1810 // Историческое развитие класса насекомых. М.: Наука, 1980. С. 104–109 (Тр. Палеонтол. ин-та АН СССР. Т. 175).
Сукачева И.Д. Историческое развитие отряда ручейников. М.: Наука, 1982. 112 с. (Тр. Палеонтол. ин-та АН СССР. Т. 197).
Шаров А.Г. Филогения ортоптероидных насекомых. М.: Наука, 1968. 217 с. (Тр. Палеонтол. ин-та АН СССР. Т. 118).
Щербаков Д.Е. О пермских и триасовых энтомофаунах в связи с биогеографией и пермо-триасовым кризисом // Палеонтол. журн. 2008. № 1. С. 15–32.
Cockerell T.D.A. Some American fossil insects // Proc. U.S. Nat. Mus. 1916. V. 51. № 2146. P. 89–106.
Eskov K.Yu., Sukatsheva I.D. Geographical distribution of the Paleozoic and Mesozoic caddisflies (Insecta: Trichoptera) // Proc. 8th Intern. Symp. Trichoptera. Columbus: Ohio Biol. Serv., 1997. P. 95–98.
Ivanov V.D., Sukatsheva I.D. Order Trichoptera Kirby, 1815. The caddis flies // History of Insects / Eds. Rasnitsyn A.P., Quicke D.I.J. Dordrecht, Boston, L.: Kluwer Acad. Publ., 2002. P. 199–220.
Fischer J., Voigt S., Schneider J.W. et al. A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries // J. Vertebr. Paleontol. 2011. V. 31. № 5. P. 937–953.
Kelly R.S., Ross A.J., Coram R.A. A review of necrotauliids from the Triassic/Jurassic of England (Trichoptera: Necrotauliidae) // Psyche. 2018. V. 2018. P. 1–12.
Melnitsky S.I., Ivanov V.D., Sukatsheva I.D. Trichoptera. Caddisflies // The Khasurty Fossil Insect Lagerstätte. N.Y., 2020. P. 1322–1350 (Paleontol. J. V. 54. № 11).
Pierwola A.A., Grimaldi D.A. First New World Necrotaulius reflects the Laurasian land masses (Insecta: Amphiesmenoptera: Necrotauliidae) // N. Jb. Geol. Paläontol. Abh. 2022. V. 304. № 1. P. 37–50.
Riek E.F. Fossil mecopteroid insects from the Upper Permian of New South Wales // Rec. Austral. Mus. 1953. V. 23. № 2. P. 55–87.
Riek E.F. Fossil insects from the Triassic beds at Mt. Crosby, Queensland // Austral. J. Zool. 1955. V. 3. № 4. P. 654–691.
Shcherbakov D.E. Magygen, Triassic Lagerstätte number one, before and after Sharov // Alavesia. 2008. V. 2. P. 113–124.
Sinitshenkova N.D. A review of Triassic mayflies, with a description of new species from Western Siberia and Ukraine (Ephemerida = Ephemeroptera) // Paleontol. J. 2000. V. 34. Suppl. 3. P. 275–283.
Sukatsheva I.D., Aristov D.S. New caddisflies of the suborder Protomeropina (Insecta: Trichoptera) from the Permian of Russia // Paleontol. J. 2013. V. 47. № 7. P. 741–751.
Tillyard R.J. Upper Permian insects of New South Wales, Part 2. The orders Mecoptera, Paramecoptera and Neuroptera // Proc. Linn. Soc. N.S.W. 1926. V. 51. № 3. P. 265–282.
Van Dijk D.E., Geertsema H. Permian insects from the Beaufort Group of Natal, South Africa // Ann. Natal Mus. 1999. V. 40. № 1. P. 137–171.
Voigt S., Buchwitz M., Fischer J. et al. Triassic life in an inland lake basin of the warm-temperate biom – the Madygen Lagerstätte (Southwest Kyrgyztan, Central Asia) // Terrestrial conservation Lagerstätten: windows into the evolution of life on land / Eds. Fraser N.C., Sues H.-D. Edinburgh: Dunedin Acad. Press, 2017. P. 65–104.
Wieland G.R. South American fossil insect discovery // Amer. J. Sci. Ser. 5. 1926. V. 12. № 68. P. 130–135.
Willmann R. Evolution und phylogenetisches System der Mecoptera (Insecta: Holometabola) // Abh. Senckenb. Naturforsch. Ges. 1989. Bd 544. 153 s.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал