

Палеонтологический журнал, 2023, № 2, стр. 18-27
Новые виды двустворок рода Streblopteria (Pectinida, Streblochondriidae) из перми северо-востока России
А. С. Бяков *
Северо-Восточный комплексный научно-исследовательский институт им. Н.А. Шило ДВО РАН
685000 Магадан, Россия
* E-mail: abiakov@mail.ru
Поступила в редакцию 09.09.2022
После доработки 20.09.2022
Принята к публикации 20.09.2022
- EDN: LDHBPD
- DOI: 10.31857/S0031031X23020071
Аннотация
Кратко рассмотрены проблемы систематики пермских пектиноидных двустворок в контексте изучения представителей известного позднепалеозойского рода Streblopteria M’Coy, и его историческое развитие на Северо-Востоке России. Из нижне- и среднепермских отложений Омолонского массива описано два новых стратиграфически важных вида: S. parenensis sp. nov. и S. gracilicosta sp. nov.
ВВЕДЕНИЕ
Пектиноидные двустворчатые моллюски являются важным элементом пермской бивальвиевой фауны Северо-Востока России. Во многих случаях отдельные их таксоны играют заметную роль для биостратиграфии, биогеографии и биономии перми региона. В то же время бóльшая часть пермских пектиноидных двустворок все еще слабо изучена, что во многом связано с фрагментарностью и часто неважной сохранностью их находок.
Среди пермских пектинид одним из наиболее часто встречающихся таксонов являются представители рода Streblopteria M’Coy, рассмотрению которого посвящена предлагаемая статья.
Несмотря на достаточно скудные морфологические характеристики раковины, которая в большинстве случаев является гладкой или имеет лишь слабую концентрическую скульптуру или струйчатость, отдельные виды этого рода достаточно четко диагностируются за счет особенностей морфологии раковины или ее скульптуры. Нередко немаловажную роль для видовой диагностики имеют и размеры раковин стреблоптерий: от очень мелких (не более 10 мм) до гигантских для рода (до 100 мм).
Стреблоптерии являются космополитами, но все-таки более тяготеют к умеренно-тепловодным бассейнам – Донецкому (Федотов, 1932), Уральскому (Лихарев, 1927), Северо-Американскому (Ciriaсks, 1963; Newell, Boyd, 1995), системе западно-европейских (Logan, 1967; Nakazava, 1999), где встречаются преимущественно в мелководных карбонатных отложениях. Нередки они и в морских бассейнах Гондваны (Dickins, 1963; Waterhouse, 1982, 2001; Gonzalez, 2006; Pagani, 2006; Neves et al., 2014; Simões et al., 2020). Ряд гондванских представителей стреблоптериеподобных форм в последнее время отнесены Б. Уотерхаузом (Waterhouse, 2008, 2014) к новым родам Concentiolineatus и Montorbicula, но этот вопрос требует дополнительного изучения и обсуждения.
ПРОБЛЕМЫ СИСТЕМАТИКИ ПОЗДНЕПАЛЕОЗОЙСКИХ ПЕКТИНИД
Как правило, для достаточно надежной диагностики таксонов пектинид необходимо знать не только строение лигамента и соотношения степени выпуклости обеих створок, но и характер скульптуры раковины, форму и особенности скульптуры ушек (особенно переднего правой створки). Учитывая все перечисленные ограничения, не удивительно, что до сих пор систематика палеозойских форм все еще остается недостаточно разработанной, несмотря на многочисленные исследования, особенно последних двух десятилетий (Waterhouse, 2001, 2008, 2014; Carter et al., 2011; Невесская и др., 2013). В прошлом веке ряд видовых, да и родовых таксонов пектиноидных был установлен на материале плохой сохранности, и часто на основании изучения только одной из створок (как правило, левых), что нередко делает невозможным четкое определение их родовой принадлежности без дополнительного изучения материала из типовых местонахождений.
Одна из последних подробных систем палеозойских пектинид была предложена Уотерхаузом (Waterhouse, 2008, 2014). Однако, на наш взгляд, многие новые таксоны как родового, так и более высокого ранга, фигурирующие в ней, представляются мало обоснованными, и вся система выглядит чересчур громоздкой. Так, например, новое подсемейство Streblopteriinae выделено лишь на основании очень длинных задних ушек (крыльев) и пологих макушечных склонов, а установленное ранее М.М. Астафьевой (Astaf’yeva, 1994) и принимаемое Уотерхаузом подсемейство Saturnellinae характеризуется лишь присутствием резких пережимов-складок на раковине. По нашему мнению, эти признаки могут иметь ранг не выше родового.
Сложность названной проблемы хорошо видна и на примере рода Streblopteria, который рассматривался различными авторами то в семействе Aviculopectinidae (Муромцева, Гуськов, 1984), то в Deltopectinidae (Newell, Boyd, 1995), то в Chaenocardiidae (Astaf’yeva, 1994), то в Streblochondriidae (Waterhouse, 2001, 2008; Carter et al., 2011; Невесская и др., 2013). Более простой, обоснованной и логичной, особенно в отношении рассматриваемого рода Streblopteria, на наш взгляд, представляется классификация, предложенная в последней работе.
РОД STREBLOPTERIA И ЕГО ИСТОРИЧЕСКОЕ РАЗВИТИЕ НА СЕВЕРО-ВОСТОКЕ РОССИИ
Стреблоптерии наиболее характерны для мелководных карбонатных фаций, особенно нижне- и среднепермских отложений Омолонского бассейна, где нередко они являются доминантами сообществ и имеют частую встречаемость. Например, в пограничных каменноугольно-пермских отложениях юго-восточного обрамления Омолонского массива, где нами было описано уникальное тепловодное сообщество двустворок (Davydov, Biakov, 2015), род Streblopteria представлен минимум четырьмя видами в одном местонахождении (биогерме). В мелководных терригенных фациях Верхоянского и Охотского бассейнов стреблоптерии встречаются гораздо реже, однако здесь они имеют важное биостратиграфическое значение для верхнепермских отложений. В глубоководных фациях, в частности, задуговых бассейнах Охотско-Тайгоносской вулканической дуги, представители этого рода практически неизвестны.
С начала изучения северо-восточно-российских пермских стреблоптерий (Люткевич, Лобанова, 1970) было известно лишь четыре вида (относимых первоначально к роду Streblochondria Newell): S. englehardti (Etheridge et Dun), S. pusilla (Schlotheim), S. sericea (Verneuil) и S. radiata (Lutkevich et Lobanova). Причем нельзя исключить, что вследствие весьма скудных морфологических характеристик первые три вида в какой-то степени являются сборными, что косвенно подтверждается весьма длительным их существованием почти на протяжении всей перми.
Всего в настоящее время в пермских отложениях Северо-Востока России нами определено не менее 16–18 видов стреблоптерий, причем почти все из них (15 видов, кроме S. rotunda и Streblopteria sp.) известны из системы Колымо-Омолонских бассейнов (преимущественно из нижней перми Омолонского) и только семь – из Верхояно-Охотского бассейна, причем здесь известно только два эндемика.
Streblopteria появляются в бассейнах Северо-Востока России еще в раннем карбоне, играя второстепенную роль в каменноугольных сообществах бентоса (Муромцева, 1974). Начало пермской истории рода связано с появлением вблизи каменноугольно–пермской границы пяти видов стреблоптерий: S. englehardti (Etheridge et Dun), S. eichwaldi (Stuckenberg), S. krasnoufimskensis Fredericks, S. sericea (Verneuil) и S. parenensis sp. nov. В дальнейшем на протяжении всей перми в то или иное время существования зонального комплекса род был представлен тремя–пятью, как правило, долгоживущими, видами. На протяжении ассельско-раннеартинского этапа развития стреблоптерий их разнообразие было особенно велико, в связи с широким развитием мелководных карбонатных фаций в Омолонском бассейне. Однако имеющийся материал пока не позволяет нам описать другие новые их виды из-за фрагментарности находок.
Во второй половине артинского века появляется долгоживущий вид S. pusilla (Schlotheim). В кунгуре следует отметить проникновение в Омолонский бассейн из Печорского S. bella Guskov. В начале роуда примечательно появление нового эндемичного для Омолонского бассейна вида S. gracilicosta sp. nov., описываемого ниже. Во второй половине кепитена интересно появление в Омолонском бассейне довольно крупных короткоживущих форм, S. aff. radiata (Lutkevich et Lobanova), имеющих слабо выраженную равномерную как концентрическую, так и радиальную скульптуру, что, в общем, не характерно для рода. В это же время или чуть позже отмечается появление мелких S. radiata (Lutkevich et Lobanova), также характеризующихся присутствием тонкой радиальной скульптуры.
Важен позднепермский этап развития стреблоптерий, когда в середине вучапина в Верхоянском и Охотском бассейнах появляются гигантские S. levis (Lutkevich et Lobanova) и S. rotunda (Lutkevich et Lobanova), причем первый вид появился в западной части Верхоянского бассейна еще в позднем кепитене. Они проникают сюда с Таймыра и севера Сибири (Бяков и др., 2021). Также интересен факт присутствия в верхах перми Балыгычанского бассейна своеобразных мелких стреблоптериеподобных форм, возможно, являющихся предковыми по отношению к некоторым триасовым таксонам (в частности, Streblopterinella Kurushin). К середине чансина (конец времени Evenicum) все стреблоптерии вымирают.
Ниже приведено описание двух новых видов стреблоптерий.
МАТЕРИАЛ И МЕТОДЫ
Изученный материал был собран как лично автором на Омолонском массиве и его юго-восточном обрамлении, так и Н.И. Караваевой, также на Омолонском массиве. Местоположение разрезов, из которых происходит описываемый материал, показано на рис. 1, стратиграфическое положение – на рис. 2.
Рис. 1.
Местонахождения новых видов рода Streblopteria на северо-востоке России, Омолонский массив: 1 – р. Левая Бургали-Коркодонская, 2 – руч. Кривой, правый приток р. Левый Кедон, 3 – р. Мунугуджак, 4 – руч. Левый Водопадный, верхнее течение р. Хивач, бассейн р. Гижига; 5 – юго-восточное обрамление Омолонского массива, р. Горная, приток р. Парень.
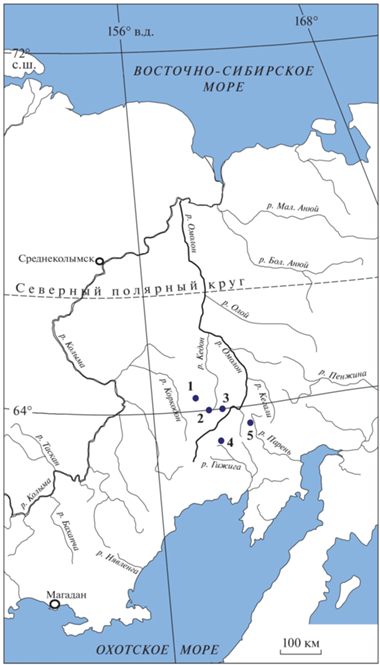
Рис. 2.
Стратиграфическое положение находок новых видов рода Streblopteria. Сокращения: МСШ – Международная стратиграфическая шкала, РСШ – Региональная стратиграфическая шкала.
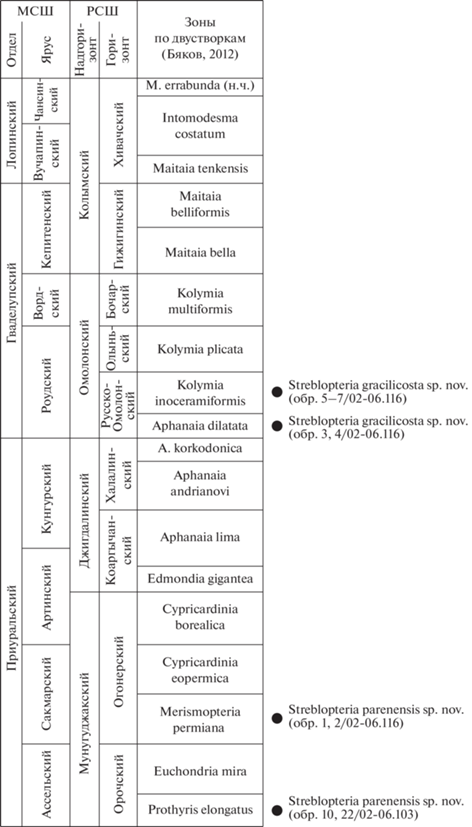
В описаниях приняты сокращения: В – высота раковины, Вп – выпуклость створки, Д – длина раковины, ДЗК – длина замочного края раковины, ДПЧ – длина передней части раковины, ДУ – длина ушек, МУ – макушечный угол, УС – угол скоса раковины.
Изученный материал хранится в музее Северо-Восточного комплексного научно-исследовательского ин-та ДВО РАН им. Н.А. Шило (СВКНИИ), г. Магадан, в колл. №№ 02-06.103, 02-06.116 и 02-06.117.
* * *
Работа выполнена по государственному заданию СВКНИИ ДВО РАН и при поддержке РФФИ, проект 20-05-00604. Автор выражает признательность хранителю ЦНИГР музея им. Ф.Н. Чернышова (С.-Петербург) Т.В. Куражевой за помощь в работе с коллекциями двустворчатых моллюсков и рецензентам С.В. Попову и А.В. Мазаеву, а также А.Ф. Банникову за внимательную работу с текстом рукописи, способствовавшую его улучшению.
ОТРЯД PECTINIDA
СЕМЕЙСТВО STREBLOCHONDRIIDAE NEWELL, 1938
Род Streblopteria M’Coy, 1851
Streblopteria: M’Coy, 1851, с. 170, part.; Waterhouse, 2001, с. 126, part. (non Waterhouse, 2008); Невесская и др., 2013, с. 249.
Типовой вид – Meleagrinella laevigata M’Coy, 1844; нижний карбон Ирландии.
Диагноз. Раковины равностворчатые или слабо неравностворчатые, округлые, слегка прозоклинные или аклинные, обычно слабовыпуклые, с различно выраженными макушками, гладкие или с очень тонкой концентрической скульптурой. У некоторых видов присутствует тонкая радиальная скульптура, обычно заметная лишь на раковине. Задние ушки небольшие, необособленные, переднее ушко правой створки и биссусный вырез хорошо развиты. Замок без зубов. Связка аливинкулярного типа, расположена на удлиненной округленно-треугольной площадке, покрытой параллельными замочному краю бороздками (рис. 3, н). Скульптура ушек подобна таковой на раковине.
Рис. 3.
Двустворчатые моллюски рода Streblopteria M’Coy из пермских отложений Северо-Востока России: а–г – S. parenensis sp. nov., Омолонский массив, нижняя пермь (×2): а – голотип № 10/02-06.103, ядро левой створки с участками сохранившейся раковины; левобережье р. Парень; ассельский ярус, мунугуджакский надгоризонт, орочский горизонт, зона по двустворкам Prothyris elongatus; обн. 4а, нижняя часть магивеемской свиты, сборы автора, 2006 г.; б – экз. № 1/02-06.116, ядро левой створки; р. Мунугуджак; сакмарский ярус, мунугуджакский надгоризонт, огонерский горизонт, зона по двустворкам Merismopteria permiana; обн. 8, средняя часть мунугуджакской свиты, сборы автора, 1999 г.; в – экз. № 2/02-06.116, ядро правой створки с частично сохранившейся раковиной; местонахождение и возраст те же; г – экз. № 22/02-06.103, неполное ядро левой створки; местонахождение и возраст те же, что и у фиг. а; д–м – S. gracilicosta sp. nov., Омолонский массив, средняя пермь, × 1.5 (кроме рис. 3, ж): д – экз. № 4/02-06.116, ядро правой створки с частично сохранившейся раковиной; р. Левая Бургали-Коркодонская; роудский ярус, омолонский надгоризонт, русско-омолонский горизонт, зона по двустворкам Aphanaia dilatatа; т.н. 40-2Б, низы омолонской свиты, сборы Н.И. Караваевой, 1980 г.; е, ж – экз. № 5/02-06.116: е – неполное ядро левой створки с сохранившейся раковиной; руч. Кривой, правый приток р. Левый Кедон; роудский ярус, омолонский надгоризонт, русско-омолонский горизонт, зона по двустворкам Kolymia inoceramiformis; обн. 2, сл. 6, нижняя часть омолонской свиты, сборы Н.И. Караваевой, 1980 г.; ж – участок скульптуры створки того же экземпляра, ×5; з – экз. № 6/02-06.116, ядро правой створки с участками сохранившейся раковины; местонахождение и возраст те же, сл. 5; и–л – голотип № 3/02-06.116, ядро двустворчатого экземпляра с частично сохранившейся раковиной; руч. Лев. Водопадный, верховья р. Хивач (бассейн р. Гижига); возраст тот же, что и у фиг. д, обн. 3а, сл. 4-2, верхи джигдалинской свиты, сборы автора, 1983 г.: и – со стороны правой створки, к – со стороны макушки, л – со стороны левой створки; м – экз. № 7/02-06.116, ядро правой створки с частично сохранившейся раковиной; местонахождение и возраст те же, что и у фиг. е, сл. 4; н – S. rotunda (Lutkevich et Lobanova), экз. № 1/02-06.117, ядро левой створки с участками сохранившейся раковины (×1.5), видно строение связочной площадки, покрытой параллельными замочному краю бороздками, типичное для рода; Центральный Таймыр, листы S–48–I, II. верховья руч. Васин; верхняя пермь, обр. 43105, сборы К.В. Борисенкова, 2017 г.

Видовой состав. В России: S. eichwaldi (Stuckenberg), S. krasnoufimskensis Fredericks, S. bella Guskov, нижняя пермь европейской части и северных регионов России; S. parenensis sp. nov., нижняя пермь Северо-Востока России; S. englehardti (Etheridge et Dun), S. pusilla (Schlotheim), нижняя–верхняя пермь европейской части и северных регионов России; S. gracilicosta sp. nov., средняя пермь Северо-Востока России; S. alenae Biakov, верхняя пермь Забайкалья, S. levis (Lutkevich et Lobanova), S. aff. radiata, S. radiata (Lutkevich et Lobanova), S. rotunda (Lutkevich et Lobanova) и Streblopteria sp., верхняя пермь Северо-Востока России.
Сравнение. От рода Streblochondria Newell, 1938, с которым сходен конфигурацией раковины, отличается отсутствием радиальной скульптуры или очень слабым ее развитием, в т.ч. и на ушках створок.
Замечания. Бóльшую часть форм, отнесенных М. Амлером (Amler, 1994) к роду Streblopteria, по нашему мнению, следует рассматривать в рамках рода Streblochondria, поскольку они имеют хорошо выраженную радиальную скульптуру ушек, а также радиальные ребра на переднем поле раковины.
Streblopteria parenensis Biakov, sp. nov.
Streblopteria sp. 5: Davydov, Biakov, 2015, с. 894, рис. 5–10 .
Название вида – по р. Парень.
Голотип – музей СВКНИИ, № 10/02-06.103; ядро левой створки с участками сохранившейся раковины; юго-восточное обрамление Омолонского массива, левобережье р. Парень; нижняя пермь, ассельский ярус, мунугуджакский надгоризонт, орочский горизонт, зона по двустворкам Prothyris elongatus; обн. 4а, нижняя часть магивеемской свиты; сборы автора, 2006 г.
Описание (рис. 3, а–г). Раковина маленькая, до 22 мм в длину; треугольно-овальной формы, с длиной, почти равной высоте (Д/В = 1.03–1.05); значительно скошенная вперед (УС = 55°–60°). Замочный край относительно длинный, около половины длины раковины (ДЗК/Д = = 0.47–0.53). Передний край в верхней части слабо вогнутый, относительно длинный, в нижней части сильно выпуклый, постепенно переходит в равномерно выпуклый, длинный нижний край, который также постепенно переходит в умеренно выпуклый длинный задний край.
Выпуклость створок раковины примерно одинаковая, умеренная (Вп/В = 0.24–0.30), точка наибольшей выпуклости расположена в верхней трети створки и несколько приближена к переднему краю. Макушки небольшие, относительно широкие, ненависающие, слабо приостренные, немного выступающие над замочным краем. Макушечный угол от 90° до 100°.
Скульптура на обеих створках представлена многочисленными очень тонкими линиями нарастания, на ядрах раковин не сохраняющимися.
Передние ушки обеих створок крупные, значительно больше задних. Переднее ушко левой створки по форме приближается к равнобедренному треугольнику со сглаженной вершиной. Переднее ушко правой створки овально-треугольное, вытянутое в длину, с относительно узким биссусным вырезом. Задние ушки створок в виде неправильного удлиненного треугольника. На ушках скульптура подобна наблюдаемой на теле раковины.
Размеры в мм и отношения
| Экз. № | Д | В | Вп | ДЗК | ДУ | Д/В | Вп/В | ДЗК/Д | УС | МУ | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| п | з | ||||||||||
| 10/02-06.103 Голотип |
20.8 | 20 | 6 | 11 | 7.6 | 3.8 | 1.04 | 0.30 | 0.53 | 60° | 90° |
| 22/02-06.103 | – | – | – | 10.8 | 8.5 | 3.8 | – | – | – | – | 95° |
| 1/02-06.116 | 22 | 21.4 | 6 | 10 | 6.5 | 3.0 | 1.03 | 0.28 | 0.47 | 55° | 95° |
| 2/02-06.116 | 22 | 21 | 5 | 9.8 | 7.8 | 4.0 | 1.05 | 0.24 | 0.47 | 55° | 100° |
Изменчивость проявляется в изменении длины замочного края по отношению к длине раковины (ДЗК/Д = 0.47–0.53).
Сравнение. Описанный вид отличается от всех известных позднепалеозойских представителей рода значительной скошенностью створок при длине, почти равной высоте.
Распространение. Омолонский массив; нижняя пермь, ассельский ярус, мунугуджакский надгоризонт, орочский – нижняя половина огонерского горизонта, зоны по двустворкам Prothyris elongatus – Merismopteria permiana.
Материал. Шесть ядер и два отпечатка левых и правых створок различной степени сохранности из двух местонахождений.
Streblopteria gracilicosta Biakov, sp. nov.
Название вида от gracilis лат. – тонкий, грациозный, и costa лат. – ребро.
Голотип – музей СВКНИИ, № 3/02-06.116; почти полное ядро двустворчатого экземпляра с частично сохранившейся раковиной; Омолонский массив, руч. Лев. Водопадный, верховья р. Хивач (бассейн р. Гижига); средняя пермь, роудский ярус, омолонский надгоризонт, русско-омолонский горизонт, зона по двустворкам Aphanaia dilatata; обн. 3а, сл. 4-2, верхи джигдалинской свиты; сборы автора, 1983 г.
Описание (рис. 3, д–л). Раковина от средних до несколько превышающих средние размеров, до 52 мм в длину; округлой формы, с длиной, почти равной высоте (Д/В = 0.97–1.08), аклинная. Замочный край умеренной длины, менее половины длины раковины (ДЗК/Д = 0.38–0.47). Передний край довольно короткий, в верхней части вогнутый, в нижней – сильно выпуклый, постепенно переходит в равномерно выпуклый, длинный нижний край, который также постепенно переходит в такой же выпуклый длинный задний край.
Выпуклость створок раковины примерно одинаковая, вероятно, незначительная (Вп/В ~ 0.16), но, возможно, это обусловлено частичной деформацией створок. Точка наибольшей выпуклости расположена в верхней четверти створки примерно на равном расстоянии от переднего и заднего краев створки. Макушки небольшие, неширокие, ненависающие, слабо приостренные, немного выступающие над замочным краем. Макушечный угол от 90° до 100°.
Скульптура на обеих створках представлена многочисленными очень тонкими равномерными густо расположенными концентрическими ребрышками (число которых составляет около 60 на 1 см вблизи нижнего края раковины), разделенными примерно такими же по ширине промежутками. На некоторых экземплярах также наблюдается едва заметная тонкая радиальная струйчатость (рис. 3, е, ж), видимая только при напылении или при косом освещении. Также присутствуют своеобразные очень сглаженные концентрические, иногда относительно равномерно расположенные морщины-складки, число которых обычно не превышает пяти–шести. На ядрах раковин тонкая концентрическая скульптура не сохраняется.
Передние ушки обеих створок крупные, значительно больше задних. Переднее ушко левой створки по форме приближается к равнобедренному треугольнику со сглаженной вершиной. Переднее ушко правой створки овально-треугольное, вытянутое в длину, с относительно узким биссусным вырезом. Задние ушки створок маленькие, в виде неправильного удлиненного треугольника. На ушках скульптура подобна наблюдаемой на теле раковины.
Размеры в мм и отношения
| Экз. № | Д | В | Вп | ДЗК | ДУ | Д/В | Вп/В | ДЗК/Д | МУ | |
|---|---|---|---|---|---|---|---|---|---|---|
| п | з | |||||||||
| 3/02-06.116 голотип |
31.8 | 32.8 | 4.5 | ?~13 | 8.7 | >2.0 | 0.97 | 0.14* | 0.41 | 100° |
| 4/02-06.116 | 26.8 | 27 | – | – | 6.0 | – | 0.99 | – | – | 90° |
| 5/02-06.116 | ~34.5 | 31.7 | 5* | 14 | 8.5 | 3.0 | 1.08 | 0.16* | 0.41 | 95° |
| 6/02-06.116 | 47 | 44 | – | 18 | ~10 | 4.0 | 1.07 | – | 0.38 | 95° |
| 7/02-06.116 | 42 | 41 | – | 16 | 9 | 4.5 | 1.02 | – | 0.38 | 95° |
Изменчивость проявляется в изменении соотношения длины раковины к ее высоте – от почти округлых очертаний створок до несколько более вытянутых в длину. Вероятно, также может варьировать степень выраженности радиальной скульптуры.
Сравнение. По очертаниям раковины наиболее близок к широко распространенному виду S. englehardti (Etheridge et Dun, 1906), однако отличается несколько более крупными размерами и своеобразным характером скульптуры.
Распространение. Омолонский массив; средняя пермь, роудский ярус, омолонский надгоризонт, русско-омолонский горизонт, зоны по двустворкам Aphanaia dilatata и Kolymia inoceramiformis.
Материал. Около 10 ядер обеих сворок и двустворчатых экземпляров, иногда с сохранившейся раковиной хорошей и удовлетворительной сохранности из четырех местонахождений.
Список литературы
Бяков А.С. Новая зональная схема пермских отложений Северо-Востока Азии по двустворчатым моллюскам. Статья 1. Зональное расчленение // Тихоокеан. геол. 2012. Т. 31. № 5. С. 13–40.
Бяков А.С., Данукалова М.К., Кузьмичев А.Б., Соболев Е.С. Пермские двустворчатые моллюски кряжа Прончищева (север Сибири): новые данные по биостратиграфии, таксономическому составу и биогеографическим связям // Палеонтол. журн. 2021. № 3. С. 24–34.
Лихарев Б.К. Верхнекаменноугольные пелециподы Урала и Тимана // Тр. Геол. ком-та. Нов. сер. 1927. Вып. 164. 140 с.
Люткевич Е.М., Лобанова О.В. Описание видов. Класс Bivalvia // Полевой атлас пермской фауны и флоры Северо-Востока СССР. Магадан: Магаданск. кн. изд-во, 1970. С. 42–63.
Невесская Л.А., Попов С.В., Гончарова И.А., Гужов А.В., Янин Б.Т., Полуботко И.В., Бяков А.С., Гаврилова В.А. Двустворчатые моллюски России и сопредельных стран в фанерозое. М.: Научный мир, 2013. 524 с.
Муромцева В.А. Двустворчатые моллюски карбона Казахстана и Сибири. Л.: Недра, 1974. 150 с. (Тр. ВНИГРИ. Вып. 336).
Федотов Д.М. Каменноугольные пластинчатожаберные моллюски Донецкого бассейна // Тр. ВГРО НКТП СССР. 1932. Вып. 103. 241 с.
Amler M.R.W. The earliest European streblopteriid bivalves (Pteriomorphia; Late Famennian) // Ann. Soc. Géol. Belg. 1994. V. 117. Fasc. 1. P. 1–17.
Astaf’yeva M.M. A revision of the Chaenocardiiae (=Streblochondriidae) (Bivalvia, Pectinoidea) // Paleontol. J. 1994. V. 28. № 1A. P. 1−22.
Carter J.G., Altaba C.R., Anderson L.C. et al. A synoptical classification of the Bivalvia (Mollusca) // Paleontol. Contrib. 2011. № 4. P. 1–48.
Ciriacks K.W. Permian and Eotriassic bivalves of the Middle Rockies // Bull. Amer. Mus. Natur. Hist. 1963. V. 125. Art. 1. 99 p.
Davydov V.I., Biakov A.S. Discovery of shallow-marine biofacies conodonts in a bioherm within the Carboniferous-Permian transition in the Omolon Massif, NE Russia near the North paleo-pole: Correlation with a warming spike in the southern hemisphere // Gondwana Res. 2015. V. 28. P. 888–897. https://doi.org/10.1016/j.gr.2014.07.008
Dickins J.M. Permian pelecypods and gastropods from Western Australia // Bull. Bur. Miner. Res. Geol. Geophys. Australia. 1963. № 63. 203 p.
Gonzalez C.R. Lower Permian bivalves from central Patagonia, Argentina // Paläontol. Z. 2006. V. 80. P. 130–155.
Logan A. The Permian Bivalvia of Northern England // Palaeontogr. Soc. Monogr. 1967. V. 121(518). 72 p.
M’Coy F. Description of some new Mountain Limestone fossils // Ann. Mag. Natur. Hist., 1851. Ser. 2. P. 167– 175.
Neves J.P., Anelli L.E., Simões M.G. Early Permian post-glacial bivalve faunas of the Itararé Group, Paraná Basin, Brazil: Paleoecology and biocorrelations with South American intraplate basins // J. South Amer. Earth Sci. 2014. V. 52. P. 203–233.
Nakazawa K. Permian bivalves from West Spitsbergen, Svalbard Islands, Norway // Paleontol. Res. 1999. V. 3. № 1. P. 1–17.
Newell N.D., Boyd D.W. Pectinoid bivalves of the Permian–Triassic crisis // Bull. Amer. Mus. Natur. Hist. 1995. V. 227. 95 p.
Pagani M.A. Los bivalvos carboniferos y permicos de Patagonia (Chubut, Argentina). Parte IV: familias Aviculopectinidae, Deltopectinidae y Schizodidae // Ameghiniana (Rev. Asoc. Paleontol. Argent.). 2006. V. 43. № 2. P. 461–476.
Simões M.G., Neves J.P., Taboada A.C. et al. Macroinvertebrates of the Capivari marine bed, late Paleozoic glacial Itarare Group, northeast Parana Basin, Brazil: Paleoenvironmental and paleogeographic implications // J. South Amer. Earth Sci. 2020. V. 98. https://doi.org/10.1016/j.jsames.2019.102433
Waterhouse J.B. Permian Pectinacea and Limacea (Bivalvia) from New Zealand // N. Z. Geol. Surv. Paleontol. Bull. 1982. № 49. 75 p.
Waterhouse J.B. Late Paleozoic Brachiopoda and Mollusca from Wairaki Downs, New Zealand // Earthwise. 2001. V. 3. 195 p.
Waterhouse J.B. Aspects of the evolutionary record for fossils of the Bivalve Subclass Pteriomorphia Beurlen // Earthwise. 2008. V. 8. 220 p.
Waterhouse J.B. Early Permian Conulariida, Brachiopoda and Mollusca from Homevale, Central Queensland // Earthwise. 2014. V. 11. 390 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал