

Палеонтологический журнал, 2023, № 3, стр. 92-96
Первая находка вида рода Acicnemis Fairmaire (Coleoptera: Curculionidae) в миоценовом суматранском янтаре
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Алтайский государственный университет
656049 Барнаул, Россия
* E-mail: fossilweevils@gmail.com
Поступила в редакцию 07.11.2022
После доработки 21.11.2022
Принята к публикации 21.11.2022
- EDN: QBWNWJ
- DOI: 10.31857/S0031031X23030091
Аннотация
Из раннемиоценового суматранского янтаря описан новый вид рода Acicnemis Fairmaire, 1849. Новый вид, A. sumatranus sp. nov., отличается от A. suturalis Roelofs, 1875 более крупными размерами тела, небольшой булавой усиков, более длинными бедрами и крупными зубцами на задних бедрах. Это первая находка трибы Acicnemidini в неогене и первое описанное жесткокрылое насекомое из суматранского янтаря.
ВВЕДЕНИЕ
Наиболее хорошо исследованы неогеновые палеоэнтомофауны американских доминиканского и мексиканского янтарей. Значительно хуже обстоит ситуация с миоценовыми янтарями Старого Света. Начато изучение эфиопского янтаря (Schmidt et al., 2010; Bouju, Perrichot, 2020; Solórzano-Kraemer et al., 2021). Появились работы, посвященные насекомым китайского янтаря из Чжанпу (Wang et al., 2021; Solórzano-Kraemer et al., 2021). Имеются предварительные данные о таксономическом составе беспозвоночных новозеланского миоценового янтаря (Schmidt et al., 2018). Фауна индонезийских янтарей, в т.ч. суматранского, остается одной из самых плохо изученных неогеновых янтарных фаун. На сегодня есть только одна публикация с описанием комара-болотницы из суматранского янтаря (Ngô-Muller et al., 2021). Из этого же янтаря указаны представители жуков – жужелиц (Carabidae), стафилинов (Staphylinidae) и плоскоходов (Platypodidae) (Ngô-Muller et al., 2019, 2021).
В настоящей статье описывается новый вид жука-долгоносика рода Acicnemis Fairmaire, 1849, принадлежащего к трибе Acicnemidini. Это первая находка жуков-долгоносиков в суматранском янтаре.
МАТЕРИАЛ И МЕТОДЫ
Описываемый экземпляр происходит из суматранского янтаря, добытого в Южно-Суматранском нефтегазоносном бассейне, в районе Синамара (округ Дхармасрая, пров. Западная Суматра, Индонезия). Возраст суматранского янтаря датируют ранним миоценом (Lambert et al., 2013). Его вероятными продуцентами были деревья семейства диптерокарповые (Dipterocarpaceae), вероятно, близкие к шорее (Shorea) (Brackman et al., 1984).
Голотип хранится в коллекции Ин-та систематики и экологии животных СО РАН (ИСЭЖ СО РАН), г. Новосибирск.
Фотографии, описание и промеры были сделаны под бинокулярным микроскопом Zeiss Stemi 2000-C с камерой AxioCam MRc5. Для обработки иллюстраций использовался Adobe Photoshop версии 21.1.3.
ТАКСОНОМИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО CURCULIONIDAE LATREILLE, 1802
ПОДСЕМЕЙСТВО MOLYTINAE SCHOENHERR, 1823
Триба Acicnemidini Lacordaire, 1866
Род Acicnemis Fairmaire, 1849
Acicnemis sumatranus Legalov, sp. nov.
Табл. XVII
Название вида – от о-ва Суматра, где обнаружен янтарь.
Голотип – SA2017/1; суматранский янтарь, ранний миоцен.
Объяснение к таблице XVII
Acicnemis sumatranus sp. nov. голотип SA2017/1, общий вид, сбоку, справа; суматранский янтарь, ранний миоцен. Длина масштабного отрезка соответствует 1.0 мм.

Описание (рис. 1). Тело темное, покрытое густыми прилегающими широкими чешуйками и более редкими полуприлегающими узкими чешуйками. Промежутки между бороздками надкрылий, помимо прилегающих чешуек, несут также довольно широкие наклонные чешуйки. Головотрубка длинная и довольно тонкая, слабоизогнутая. Длина головотрубки едва меньше длины переднеспинки, в 7.9 раза больше ширины на вершине, в 5 раз больше ширины на середине, примерно в 2.9 раза больше ширины на основании. Усиковые бороздки явственные, латеральные. Глаза не выступают из контура головы. Виски длиннее глаза. Стволик довольно длинный, не достигает глаза. Жгутик семичлениковый. Второй–восьмой членики усика удлиненно-конические. Длина первого членика примерно в 5.2 раза больше длины второго членика. Длина второго членика примерно в 1.7 раза больше его ширины на вершине. Длина третьего членика примерно в 3.8 раза больше его ширины на вершине и в 1.5 раза больше длины второго членика. Ширина третьего членика составляет примерно 0.7 ширины второго членика. Длина четвертого членика в 2.5 раза больше его ширины на вершине и составляет примерно 0.7 длины третьего членика. Ширина четвертого членика примерно равна ширине третьего членика. Длина пятого членика примерно в 1.8 раза больше его ширины на вершине и составляет 0.8 длины четвертого членика. Ширина пятого членика примерно в 1.1 раза больше ширины четвертого членика. Длина шестого членика примерно в 1.3 раза больше его ширины на вершине и составляет 0.8 длины пятого членика. Ширина шестого членика примерно равна ширине пятого членика. Булава небольшая, компактная, густо покрытая волосками. Переднеспинка удлиненная, без торчащих чешуек. Диск переднеспинки слабовыпуклый. Надкрылья довольно длинные, выпуклые. Плечи явственные. Основания надкрылий не скрывают основание переднеспинки. Длина надкрылий в 5.6 раза больше длины переднеспинки. Бороздки надкрылий явственные, узкие. Промежутки широкие, плоские. Переднегрудь без бороздки для вкладывания головотрубки. Тазиковые впадины переднегруди разделенные. Заднегрудь немного уплощенная, ее длина примерно в 5 раз больше длины заднего тазика. Брюшко выпуклое. Первый и второй вентриты немного вдавленные посередине. Длина первого вентрита в 3.4 раза больше длины заднего тазика. Длина второго вентрита примерно в 1.1 раза больше длины первого вентрита. Длина третьего вентрита составляет примерно 0.2 длины второго вентрита. Длина четвертого вентрита в 1.2 раза больше третьего вентрита. Длина пятого вентрита в 1.4 раза больше длины четвертого вентрита.
Рис. 1.
Acicnemis sumatranus sp. nov., голотип SA2017/1: а – общий вид, сбоку, справа; б – членики усиков; суматранский янтарь, ранний миоцен. Обозначения: an2 – второй членик усиков, an3 – третий членик усиков, an4 – четвертый членик усиков, an5 – пятый членик усиков, an6 – шестой членик усиков, cl – булава усиков. Длина масштабного отрезка соответствует 1.0 мм для а и 0.5 мм для б.
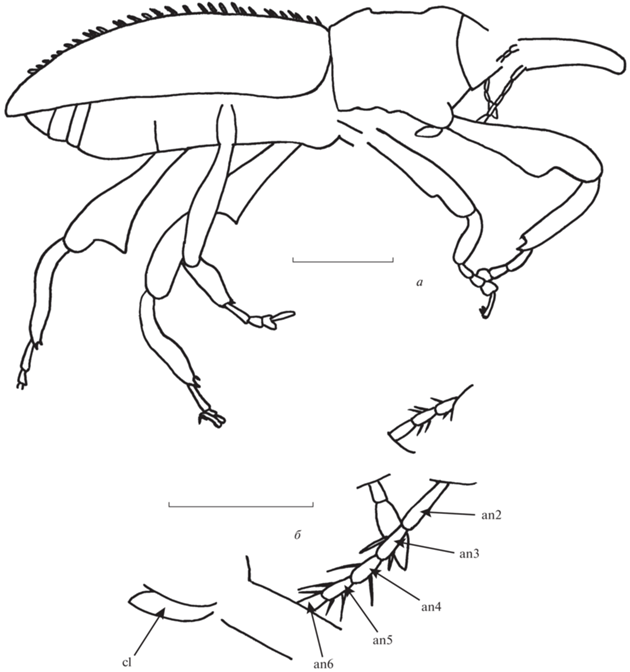
Ноги длинные. Задние тазики поперечные. Бедра удлиненные, расширенные за серединой, с крупным зубцом в вершинной трети. Задние бедра более крупные, чем средние и передние бедра, их вершины достигают середины пятого вентрита, с треугольным большим зубцом перед вершиной. Снаружи край зубца почти прямой. Длина переднего бедра примерно в 4.0 раза больше ширины перед зубцом и примерно в 1.7 раза больше длины передней голени. Длина среднего бедра примерно в 4.2 раза больше ширины перед зубцом и примерно в 1.4 раза больше длины средней голени. Длина заднего бедра примерно в 4.7 раза больше ширины перед зубцом и примерно в 1.7 раза больше длины задней голени. Голени явственно двояковогнутые, на вершине с крупным ункусом и двумя пучками щетинок. Длина передней голени примерно в 3.6 раза больше наибольшей ширины. Длина средней голени примерно в 4.5 раза больше наибольшей ширины. Длина задней голени примерно в 4.0 раза больше наибольшей ширины. Лапки длинные, короче голеней. Первый и второй членики лапок конические. Первый членик лапок более длинный, чем второй. Третий членик лапок двухлопастный. Пятый членик удлиненный. Коготки свободные, без зубцов. Длина задней лапки составляет 0.7 длины голени. Длина первого членика передних лапок примерно в 2.7 раза больше ширины на вершине. Длина второго членика передних лапок примерно в 1.1 раза больше ширины на вершине и составляет примерно 0.5 длины первого членика. Ширина второго членика передних лапок немного больше ширины первого членика. Длина третьего членика передних лапок составляет 0.6 ширины на вершине и 0.8 длины второго членика. Ширина третьего членика передних лапок примерно в 1.5 раза больше ширины второго членика. Длина пятого членика передних лапок примерно в 4.8 раза больше ширины на вершине и примерно в 2.7 раза больше длины третьего членика. Ширина пятого членика передних лапок составляет примерно 0.3 ширины третьего членика. Длина первого членика средних лапок примерно в 2.8 раза больше ширины на вершине. Длина второго членика средних лапок примерно в 1.4 раза больше ширины на вершине и составляет 0.5 длины первого членика. Ширина второго членика средних лапок примерно равна ширине первого членика. Длина третьего членика средних лапок составляет примерно 0.7 ширины на вершине и примерно 0.8 длины второго членика. Ширина третьего членика средних лапок примерно в 1.7 раза больше ширины второго членика. Длина пятого членика средних лапок примерно в 4.3 раза больше ширины на вершине и примерно в 1.9 раза больше длины третьего членика. Ширина пятого членика средних лапок составляет примерно 0.3 ширины третьего членика. Длина первого членика задних лапок примерно в 3.1 раза больше ширины на вершине. Длина второго членика задних лапок примерно в 1.6 раза больше ширины на вершине и составляет 0.5 длины первого членика. Ширина второго членика задних лапок примерно равна ширине первого членика. Длина третьего членика задних лапок составляет примерно 0.7 ширины на вершине и 0.7 длины второго членика. Ширина третьего членика задних лапок примерно в 1.6 раза больше ширины второго членика. Длина пятого членика задних лапок примерно в 3.8 раза больше ширины на вершине и примерно в 1.9 раза больше длины третьего членика. Ширина пятого членика задних лапок составляет примерно 0.4 ширины третьего членика.
Размеры в мм: длина тела без головотрубки – 4.7; длина головотрубки – 1.3.
Сравнение. Новый вид отличается от A. suturalis Roelofs, 1875 более крупными размерами тела, небольшой булавой усиков, более длинными бедрами и крупными зубцами на задних бедрах.
Замечания. Новый вид отнесен к семейству Curculionidae, поскольку у него коленчатые усики, удлиненные первый и второй вентриты и короткие третий и четвертый вентриты, а также голени с крупным ункусом и пучками волосков на вершине. Последний признак также указывает на принадлежность к подсемейству Molytinae. Новый вид помещен в трибу Acicnemidini, поскольку обладает переднегрудью без бороздки для вкладывания головотрубки; коническим вторым члеником лапок; основаниями надкрылий, не скрывающими основание переднеспинки; латеральными усиковыми бороздками; поперечными задними тазиками; бедрами с зубцами; булавой, густо покрытой волосками. Новый вид относится к роду Acicnemis, поскольку характеризуется задними бедрами более крупными, чем средние и передние бедра, с треугольным большим зубцом перед вершиной, причем снаружи край зубца почти прямой, а также лапками с двухлопастным, довольно широким третьим члеником.
Материал. Голотип.
ЗАКЛЮЧЕНИЕ
Триба Acicnemidini состоит из шести современных родов, встречающихся в Ориентальной, Палеарктической и Австралийской областях (Alonso-Zarazaga, Lyal, 1999; Lyal, 2014; Alonso-Zarazaga et al., 2017). Род Acicnemis, включающий в себя около 200 видов, распространен от юга Дальнего Востока России (Приморье, Сахалин и Курилы) на севере до Австралии на юге, от Кавказа на западе до Французской Полинезии на востоке (Alonso-Zarazaga, Lyal, 1999; Alonso-Zarazaga et al., 2017). Центр разнообразия приходится на Большие Зондские о-ва: так, на Суматре и Калимантане зарегистрировано более чем по 30 видов на каждом (Hubenthal, 1919a, b; Klima, 1935). В палеонтологической летописи триба представлена довольно бедно. Единственный палеогеновый род трибы, Electrotribus Hustache, 1942, с тремя видами, известен из балтийского янтаря (Hustache, 1942; Ulke, 1947; Voss, 1953, 1972; Legalov, 2020a, b). Следует отметить, что один из видов этого рода, E. theryi Hustache, 1942, встречается в балтийском янтаре очень часто. Другие представители трибы в палеогене пока не обнаружены (Legalov, 2015). Еще один вид рода Acicnemis, A. fossilis Kleine, 1924, описан из четвертичного копала с о-ва Сулавеси (Kleine, 1924). Таким образом, только два рода трибы Acicnemidini представлены в палеонтологической летописи, причем их ископаемые находки не выходят за пределы современного ареала этой трибы.
Список литературы
Alonso-Zarazaga M.A., Barrios H., Borovec R. et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea // Monografías electrónicas. 2017. V. 8. 729 p.
Alonso-Zarazaga M.A., Lyal C.H.C. A World Catalogue of Families and Genera of Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae). Barcelona: Entomopraxis, 1999. 315 p.
Bouju V., Perrichot V.A. Review of amber and copal occurrences in Africa and their paleontological significance // BSGF – Earth Sci. Bull. 2020. V. 191. № 17. P. 1–11.
Brackman W., Spaargaren K., van Dongen J.P.C.M. et al. Origin and structure of the fossil resin from an Indonesian Miocene coal // Geochim. Cosmochim. Acta. 1984. V. 48. № 12. P. 2483–2487.
Hubenthal W. Beiträge zur Kenntnis der Curculioniden-Gattung Acicnemis Lacordaire (Col.) // Arch. Naturgesch. Abt. A. 1919a. Bd 83. № 8. S. 92–160.
Hubenthal W. Beiträge zur Kenntnis der Curculioniden-Gattung Acicnemis Lacordaire (Col.) // Arch. Naturgesch. Abt. A. 1919b. Bd 83. № 9. S. 53–155.
Hustache A. Un curculionide de l’ambre de la Baltique // Bull. Mens. Soc. Lin. Lyon. 1942. V. 11. № 7. P. 108–109.
Kleine R. Neue Arten der Gattung Acicnemis Lacordaire nebst faunistischen Mitteilungen // Arch. Naturgesch. Abt. A. 1924. Bd 90. № 4. S. 148–164.
Klima A. Curculionidae: Acicnemidinae // Coleopterorum Catalogus Auspiciis et Auxilio. 1935. V. 145. P. 1–10.
Lambert J.B., Levy A.J., Santiago-Blay J.A., Wu Y. Nuclear magnetic resonance characterization of Indonesian amber // Life: The Excitement of Biology. 2013. V. 1. № 3. P. 136–155.
Legalov A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea) // Paleontol. J. 2015. V. 49. № 13. P. 1442–1513.
Legalov A.A. A review of the Curculionoidea (Coleoptera) from European Eocene ambers // Geosciences. 2020a. V. 10. № 1. P. 1–74.
Legalov A.A. Fossil history of Curculionoidea (Coleoptera) from the Paleogene // Geosciences. 2020b. V. 10. Spec. Issue. P. 1–51.
Lyal C.H.C. 3.7.7. Molytinae Schoenherr, 1823 // Handbook of Zoology. Vol. IV. Arthropoda: Insecta. Part 40. Coleoptera (Beetles). Vol. 3: Morphology and Systematics (Phytophaga) / Eds. Leschen R.A.B., Beutel R.G. Berlin; N.Y.: Walter de Gruyter, 2014. P. 529–569.
Ngô-Muller V., Garrouste R., Carbuccia B. et al. First terrestrial arthropod records from the Miocene amber of Sumatra // Abstracts of 8th Intern. conf. fossil insects, arthropods and amber. Santo Domingo, 2019. P. 116–117.
Ngô-Muller V., Garrouste R., Pouillon J.-M. et al. The first representative of the fly genus Trentepohlia subgenus Mongoma in amber from the Miocene of Sumatra (Diptera: Limoniidae) // Hist. Biol. 2021. V. 33. № 2. P. 254–257.
Schmidt A.R., Kaulfuss U., Bannister J.M. et al. Amber inclusions from New Zealand // Gondwana Res. 2018. V. 56. P. 135–146.
Schmidt A.R., Perrichot V., Svojtka M. et al. Cretaceous African life captured in amber // Proc. Nat. Acad. Sci. USA. 2010. V. 107. № 16. P. 7329–7334.
Solórzano-Kraemer M.M., Hammel J.U., Kunz R. et al. Miocene pinhole borer ambrosia beetles: new species of Diapus (Coleoptera: Curculionidae: Platypodinae) // Palaeoworld. 2021. https://doi.org/10.1016/j.palwor.2021.10.001
Ulke T. A new genus and species of Curculionidae (Coleoptera) in Baltic amber // Notulae Natur. 1947. № 19. P. 1–5.
Voss E. Einige Rhynchophoren der Bernsteinfauna (Coleoptera) // Mitt. Geol. Staat. Hamburg. 1953. Bd 22. S. 119–140.
Voss E. Einige Rüsselkäfer der Tertiärzeit aus baltischen Bernstein (Coleoptera, Curculionidea) // Steenstupia. 1972. V. 2. P. 167–181.
Wang B., Shi G., Xu C. et al. The mid-Miocene Zhangpu biota reveals an outstandingly rich rainforest biome in East Asia // Sci. adv. 2021. V. 7. № 18. P. eabg0625.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал