

Палеонтологический журнал, 2023, № 5, стр. 94-106
Ovis gracilis sp. nov. (Artiodactyla, Bovidae) из нижнего плейстоцена пещеры Таврида в Крыму и история рода Ovis
И. А. Вислобокова *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: ivisl@paleo.ru
Поступила в редакцию 15.02.2023
После доработки 16.03.2023
Принята к публикации 16.03.2023
- EDN: JHGEAH
- DOI: 10.31857/S0031031X23050112
Аннотация
На основании изучения ископаемых остатков Bovidae рода Ovis из нижнеплейстоценовых отложений пещеры Таврида в Крыму выделен новый вид – O. gracilis sp. nov. Этот не очень крупный баран имел гомонимно скрученные рога с короткими синусами и стройные метаподии. Остатки Ovis в Тавриде – одни из древнейших в Европе и первое свидетельство присутствия Ovis в раннем плейстоцене Крыма. Вид O. gracilis входил в состав фауны пещеры Таврида, возраст которой, по данным биохронологии, оценивается в пределах 1.8–1.5 млн лет. Появление этого вида на юге Восточной Европы могло быть связано с одной из ранних дисперсионных волн Ovis из Азии, вдоль Альпийско-Гималайского пояса, во время глобального похолодания перед палеомагнитным эпизодом Олдувей.
ВВЕДЕНИЕ
Дикие бараны рода Ovis L., 1758 довольно широко распространены в современной фауне горных районов Евразии и Северной Америки, хотя их ареалы и численность заметно сокращаются (напр., Michel, Ghoddousi, 2020a, b; Reading et al., 2020). В палеонтологической летописи они известны с плиоцена, но находки в плиоцене и нижнем плейстоцене Евразии очень редки, и ранняя история этого компонента плейстоценовой мегафауны Палеарктики остается плохо изученной. Новые материалы из отложений этого возраста вызывают особый интерес.
Самые древние Ovis описаны из верхнего плиоцена местонахождения Удунга в Забайкалье (Vislobokova et al., 1995; Калмыков, 2013). В Европе единичные остатки раннеплейстоценовых Ovis – Ovis sp. – известны из четырех местонахождений: часть черепа с основанием рогового стержня из Сенеза во Франции (Schaub, 1943), вторая фаланга из Сливницы (Spassov, Crégut-Bonnoure, 1999; Spassov, 2005), отдельные зубы, две кости заплюсны, третья фаланга из пещеры Козарника в Болгарии и верхний моляр М2 из Аполлонии-1 в Греции (Fernandez, Crégut, 2007; Kostopoulos et al., 2022). Кроме того, в состав фауны Куэва Виктория в Испании, возможно, входил O. ammon antiqua Pommerol, 1880, характерный для среднего плейстоцена Европы (Crégut-Bonnoure, 2007).
Недавно остатки Ovis (роговой стержень, верхние и нижние челюсти и метаподии) были обнаружены среди материалов из раскопок 2020–2022 гг. в пещере Таврида в 15 км к востоку от Симферополя, Крым. Они найдены в основном костеносном слое, возраст которого по фауне млекопитающих 1.8–1.5 млн лет. Эта фауна сопоставляется с поздним виллафранком Западной Европы, зоной млекопитающих MNQ18 (Лопатин и др., 2019).
Род Ovis хорошо отличается от других родов Bovidae по морфологии рогов и синусов (полостей) в роговом стержне, зубной системы и костей конечностей. Современные виды диких баранов различаются, в частности, общими размерами, размерами рогов, типом их скручивания, особенностями передней (фронтальной) поверхности (по: Groves, Grubb, 2011). Диагностика ископаемых видов Ovis включает в себя размеры, а также особенности строения роговых стержней, щечных зубов и метаподий. Ископаемые остатки из пещеры Таврида относятся к одной из древнейших европейских популяций рода Ovis, которая выделяется в новый вид.
Изучение материалов из Тавриды дает новую информацию о ранней истории Ovis в Европе и расширяет представления о биоразнообразии раннеплейстоценовой фауны Тавриды и ее зоогеографических связях.
Автор выражает благодарность А.В. Лаврову, И.А. Ермольчику (Палеонтологический ин-т им. А.А. Борисяка РАН – ПИН РАН), Д.О. Гимранову (Ин-т экологии растений и животных УрО РАН) и всем участникам раскопок за сбор материалов, Ф. Райволю (Каталонский ин-т палеоэкологии человека и социальной эволюции, Испания) и М. Ватабэ (Ун-т Васэда, Япония) за присланные статьи, а также Р.А. Ракитову (ПИН РАН) за томографию рогового стержня и С.В. Багирову (ПИН РАН) за фотографии образцов.
Исследование выполнено за счет гранта Российского научного фонда № 22-14-00214, https://rscf.ru/project/22-14-00214/.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО BOVIDAE GRAY, 1821
ПОДСЕМЕЙСТВО CAPRINAE GILL, 1872
Триба Caprini Gray, 1821
Род Ovis L., 1758
Ovis gracilis Vislobokova, sp. nov.
Название вида gracilis лат. – стройный.
Голотип – ПИН, № 5644/308, фрагмент левого рогового стержня (рис. 1); Россия, Крым, пещера Таврида; нижний плейстоцен (поздний виллафранк, MNQ 18).
Рис. 1.
Ovis gracilis sp. nov.: а–г – голотип ПИН, № 5644/308, часть левого рогового стержня спереди (а), сзади (б), снаружи (в), изнутри (г); д– реконструкция положения рогового стержня; е – реконструкция облика животного; ж–и – томография голотипа: спереди (ж), сверху (з), снизу (и); поперечные сечения (к, л); продольные сечения (м, н); цифры 1 и 2 показывают положение продольных сечений, а белый квадрат – положение увеличенного фрагмента c окончаниями синусов; Крым, пещера Таврида; нижний плейстоцен.

Описание (рис. 1–3). Размеры сравнительно небольшие. Роговые стержни аркообразно изогнутые дорсально, гомонимно скрученные в дистальной половине, сильно сужающиеся к дистальному концу. Они округленные в сечении, с переднелатеральным и медиальным ребрами, задним килем и уплощенной передней поверхностью. Синусы короткие. Ряд премоляров длинный. Коронки щечных зубов не очень высокие. Метаподии длинные, стройные.
Рис. 2.
Ovis gracilis sp. nov.: а–в – экз. ПИН, № 5644/309, фрагмент правой верхнечелюстной кости с Р2–Р4; г–е – экз. ПИН, № 5644/310, левая верхнечелюстная кость с dP2–M2; ж–и – экз. ПИН, № 5644/311, фрагмент левой ветви нижнечелюстной кости с M1–M3; а, г, ж – с буккальной, б, д, з – с лингвальной, в, е, и – с окклюзиальной сторон; Крым, пещера Таврида; нижний плейстоцен.
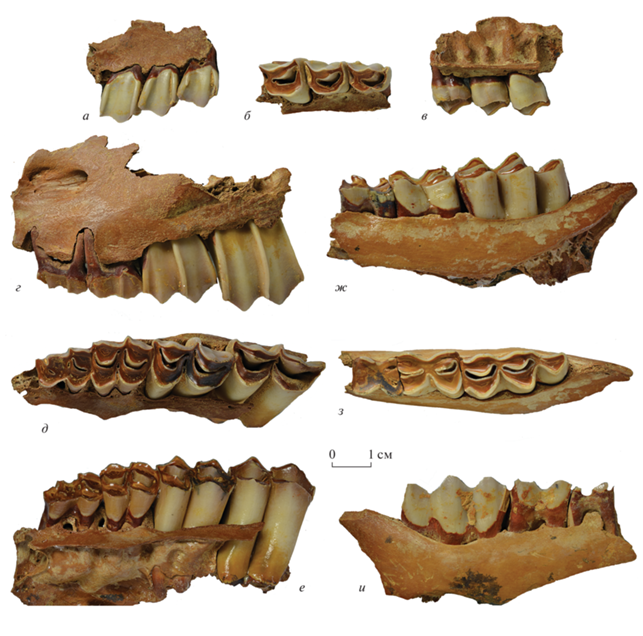
Рис. 3.
Ovis gracilis sp. nov.: а–г – экз. ПИН, № 5644/312, левая пястная кость; д–ж – экз. ПИН, № 5644/313, правая плюсневая кость; а, д – спереди, в, ж – сзади, б, е – сверху, г – снизу; Крым, пещера Таврида; нижний плейстоцен.

Голотип представляет собой среднюю часть левого рогового стержня взрослого самца: внизу он обломан на уровне уплощения передней поверхности (вероятно, на расстоянии около 10 см от основания); его латеральная поверхность местами разрушена, а также повреждена погрызами. Эта часть рогового стержня сильно аркообразно изогнута дорсально, слегка гомонимно скручена (примерно на 15°) и сужается к дистальному концу; дистальный конец заворачивается наружу. Передняя (фронтальная) поверхность сужается кверху, уплощена в нижней трети, с небольшим гребнем у проксимального конца. Переднелатеральное и медиальное ребра сглаженные. Киль на задней поверхности выше трех см от проксимального конца резкий. Поперечное сечение у проксимального конца округленное, немного сужающееся кзади, со слегка уплощенной передней стороной, а у дистального конца круглое, с заостренным задним краем. Костная ткань рогового стержня состоит из тонкого плотного внешнего слоя и пористой внутренней части. На проксимальном конце сохранились следы окончаний синусов (рис. 1, з, и). Компьютерная томография показала, что они имеют вид ячеек (небольших камер), разделенных распорками (рис. 1, и, н). На поперечных и продольных сечениях хорошо видна система крупных и мелких сосудистых каналов (рис. 1, и–н). На продольном сечении видны похожие на септы структуры (тонкие полосы), направленные под углом к поверхности. Они наиболее выражены во фронтальной части рогового стержня.
Верхние премоляры экз. ПИН, № 5644/309 умеренно стерты (рис. 2, а–в). Коронки немного отклонены назад, с хорошо развитыми парастилем, метастилем и столбиком паракона. Метастиль очень сильно выступает буккально. Столбик паракона утолщается к основаниям коронок, на P2 его передненаружный край округлен и на поверхности окклюзии образует уступ по отношению к более узкой передней части коронки. Центральная долинка сужается кзади, на Р3 и Р4 она с очень маленькой задней шпорой. Дополнительные маленькие узкие поперечные долинки присутствуют у заднего края коронок. На P2 протокон и гипокон не полностью слиты на поверхности окклюзии, а на лингвальной стороне они разделены глубокой бороздкой. Лингвальная сторона коронки P3 уплощена и лишь слегка вогнута в средней части у основания. Коронка Р4 с почти прямыми передним и задним краями и выпуклой лингвальной стороной.
Верхнечелюстная кость экз. ПИН, № 5644/310 обломана спереди у альвеолы dP2 и позади М2 (рис. 2, г–е). Подглазничное отверстие, foramen infraorbitale, расположено на уровне dP3/dP4.
Коронки молочных зубов dP3 и dP4 довольно сильно стерты, на поверхности окклюзии dP3 от передней долинки остались лишь ее концы. Коронка моляра М2 стерта частично. Коронки dP3 и dP4 с широким парастилем (на dP4 он отбит с буккальной стороны), выступающим буккально мезостилем и более слабым метастилем. На dP3 передняя доля коронки у́же и длиннее задней доли, столбик паракона очень сильно утолщен, мощный, с округлым передненаружным краем. Стертый цингулумообразный валик на dP3 протягивается почти до передненаружного угла коронки.
Коронка М2 сравнительно невысокая, она удлинена и заметно крупнее коронки М1. Задние половины коронок моляров длиннее передних. На буккальной стороне коронок столбик паракона развит сильнее столбика метакона, парастиль немного толще метастиля, а мезостиль наиболее сильно выступает буккально. Лингвальные стенки протокона и гипокона округленные, передние лингвальные полулуния немного заострены лишь на поверхности окклюзии.
Нижние моляры экз. ПИН, № 5644/311 сильно стерты, с цементом; коронка M1 обломана при жизни и стерта почти до основания (рис. 2, ж–и). Коронки M2 и M3 с уплощенной лингвальной стенкой, без метастилида и эктостилида, с широкой капринной складкой.
Рельеф поверхности стирания моляров (т.e. мезостирания) сравнительно высокий, лабиальные бугорки в основном острые или слабо округленные, углубления между бугорками достаточно глубокие.
Пястная кость экз. ПИН, № 5644/312 удлиненная, стройная (рис. 3, а–г). На проксимальной суставной поверхности фасетка для os trapezoideocapitatum с сильно выступающим вперед передним краем и вырезкой у медиального края. Отверстие между фасетками маленькое. Бугор на передней поверхности кости для лучевого разгибателя запястья, m. extensor carpi radialis, сильно развит. Передняя (дорсальная) поверхность кости круто выпуклая до нижнего питательного отверстия. Задний край проксимальной суставной поверхности вогнутый. На задней (пальмарной) поверхности кости продольный желоб, sulcus longitudinalis palmaris, глубокий и длинный, нижняя треть диафиза уплощенная, отверстие канала canalis metacarpi distalis маленькое, гребни суставных валиков не заходят на диафиз. Расширение проксимальной и дистальной частей диафиза пястной и плюсневых костей к эпифизам не резкое, контур дистального эпифиза (при взгляде спереди) напоминает прямоугольник.
Плюсневая кость экз. ПИН, № 5644/313 с обломанным дистальным эпифизом (рис. 3, д–ж). Переднезадний диаметр проксимального конца заметно превосходит его поперечный диаметр (индекс DAPprox/DTprox 116%), что характерно для Ovis (см. Громова, 1953). Проксимальная суставная поверхность с выступающим вперед, немного угловатым передним краем медиальной фасетки для os cuneiforme intermediolaterale (os tarsale II+III). Медиальная сторона суставной поверхности менее выпуклая, чем латеральная. Угол заднего края задней медиальной фасетки для os cuneiforme mediale (os tarsale I) сильно выступает назад. Задняя латеральная фасетка для os naviculocuboideum короткая, треугольная, сильно наклонена наружу. Задний край проксимальной поверхности между этим фасетками V-образно вогнут. Питательное отверстие на суставной поверхности между передними фасетками маленькое. Продольный желоб на передней поверхности, sulcus longitudinalis dorsalis, углубляется примерно с середины длины кости и расширяется у дистального конца. Отверстие canalis metatarsi proximalis на задней поверхности у проксимального конца маленькое (двойное). Продольный желоб на задней поверхности, sulcus longitudinalis plantaris, глубокий и длинный.
Размеры в мм, индексы в %. Голотип ПИН, № 5644/308, фрагмент рогового стержня: длина (L), по прямой – сa. 120, по большой кривизне – сa. 140, наибольший периметр проксимального конца – са. 170; переднезадний и поперечный диаметры проксимального конца (DAP × DT) – 60 × 47, индекс сжатия (DT/DAP) – 78; переднезадний и поперечный диаметры дистального конца – 41.5 × ca. 35.
Экз. ПИН, № 5644/309, фрагмент верхнечелюстной кости с Р2–Р4, длина Р2–Р4 – 38.3 мм; зубы (L × W): Р2 – 12.42 × 10.35; Р3 – 12.76 × 12.3; P4 – 13.1 × 14.48.
Экз. ПИН, № 5644/310, верхнечелюстная кость с dP2–M2, длина и ширина (L × W) зубов: dP3 – 14.2 × 10.3; dP4 – 15.6 × 13.3; М1 –19.2 × 13.8; М2 – 22.79 × 15.2; высота с буккальной стороны 23.14.
Экз. ПИН, № 5644/311, фрагмент нижнечелюстной кости с M1–M3, длина M1–M3 – 57.15; зубы (L × W): М2 – 15 × 11.2; М3 – 20.1 × 12.1.
Экз. ПИН, № 5644/312, пястная кость: L – 210, DTprox × DAPprox – 31.6 × 22.2, DTdist × DAPdist – 32 × 22.1; DTdiaph × DAPdiaph – 18.2 × 16.3; индекс DAPprox/DTprox – 70; индекс DTdist/L – 15.
Экз. ПИН, № 5644/313, плюсневая кость: DTprox × DAPprox – 28.59 × 32.8, DTdiaph × DAPdiaph – 17.9 × 21.1; индекс DAPprox/DTprox –116.
Сравнение. Форма рогового стержня, форма его поперечного сечения, особенности строения зубов и метаподий, а также морфометрические показатели определенно свидетельствуют о принадлежности описываемых материалов к роду Ovis. Архитектура костной ткани голотипа и синусов (с распорками) в целом сходна с таковой у современных Ovis (напр., Schaffer, Reed, 1972; Vrba, Schaller, 2000; Farke, 2010; Drake et al., 2016). Септовидные пластины похожи по своему положению на септы роговых стержней у современных Ovis, у которых они усиливают прочность фронтальной области. Эта часть роговых стержней у Ovis испытывает наибольшее ударное давление во время боев. Cвоеобразное сочетание примитивных и прогрессивных признаков отличает O. gracilis от всех других видов.
По морфологии O. gracilis sp. nov. проявляет наибольшее сходство с О. shantungensis Matsumoto, 1926 (=О. ammon shantungensis Matsumoto, 1926) из нижнего плейстоцена Китая. Но по размерам он немного меньше и отличается от этого вида менее мощными роговыми стержнями, более крутым изгибом переднего края и более сильным сужением к дистальному концу, а также некоторыми особенностями строения щечных зубов и более стройными метаподиями. Вид О. shantungensis описан из местонахождения Ванчиагна в пров. Шаньдун (Matsumoto, 1926) и местонахождений бассейна Нихэвань в пров. Хэбей (Teilhard de Chardin, Piveteau, 1930; Tong et al., 2021, 2022). На черепе из Санкан-хо (Cяшагоу) бассейна Нихэвань видно, что синусы у этого вида достигали уровня середины аркообразного изгиба (Teilhard de Chardin, Piveteau, 1930, табл. XIV , фиг. 1), а возможно, поднимались и выше. Длина ряда премоляров Р2–Р4 у O. gracilis sp. nov. попадает в предел изменчивости у O. shantungensis (29–39 мм, m = 32.67 мм, n = 3) (Teilhard de Chardin, Piveteau, 1930), и их строение, а также строение молочных зубов, в целом сходно с таковым у китайского вида. Но коронка Р2 у O. gracilis sp. nov. немного длиннее и со слабее слитыми протоконом и гипоконом (примитивные признаки). Передний цингулумообразный валик на dP3 у O. gracilis sp. nov. немного длиннее, чем у O. shantungensis (“bourrelet”: Teilhard de Chardin, Piveteau, 1930, рис. 26 ). Гипсодонтность у O. gracilis sp. nov., очевидно, была меньше; индекс высоты коронки М2 (107%) ниже, чем у O. shantungensis (187%: Teilhard de Chardin, Piveteau, 1930, табл. XIII , фиг. 1). Пястная кость у O. gracilis sp. nov. немного короче и стройнее, чем у O. shantungensis (L – 221–233 мм, индекс DTdist/L – 18–19%: Tong, 2022). По массивности пястной кости O. gracilis sp. nov. ближе к современному муфлону O. orientalis Gmelin, 1774 (15.8–16.9%: Громова, 1953).
От O. zdanskyi Bohlin, 1938 из провинции Хэнань (Loc. 18, Ян-Шао-Цун; верхний плейстоцен: Wang et al., 2016) отличается меньшими размерами, менее утолщенными роговыми стержнями, менее развитыми синусами и более примитивным строением зубной системы. У O. zdanskyi ряд верхних премоляров значительно сокращен (см. Bohlin, 1938, табл. VIII , фиг. 4).
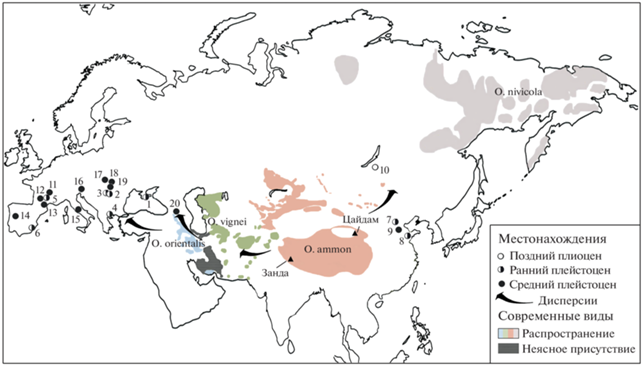
Рис. 4.
Основные местонахождения и дисперсии Ovis в позднем плиоцене – среднем плейстоцене и распространение современных видов в Евразии (c использованием данных: Rivals, 2004; Crégut-Bonnoure, 2007; Harris, Tsytsulina, 2008; Wang et al., 2016; Michel, Ghoddousi, 2020a, b; Reading et al., 2020; Kostopoulos et al., 2022; Tong et al., 2022). Местонахождения: 1 – Таврида, 2 – Сливница, 3 – Козарника, 4 – Аполлония-1, 5 – Сенез, 6 – Куэва Виктория, 7 – Нихэвань, 8 – Ванчиагна, 9 – Чжоукоудянь, 10 – Удунга, 11 – Пон-дю-Шато, 12 – Камп-де-Пейре, 13 – Араго, 14 – Галерия Песада, 15 – Маглиани, 16 – Визоглиано, 17 – Странска Скала, 18 – Уппони 1, 19 – Тарко, 20 – Кударо 1.
От O. ammon antiqua Pommerol, 1880 (=O. antiqua Pommerol, 1880) из среднего плейстоцена Европы отличается сравнительно небольшими размерами, менее утолщенными роговыми стержнями, их более сильным сужением к дистальному концу, более короткой уплощенной фронтальной частью, а также некоторыми примитивными особенностями зубной системы. Голотип O. antiqua, мозговой отдел черепа с неполными роговыми стержнями, и паратипы найдены в местонахождении Пон-дю-Шато во Франции (Pommerol, 1880), неполные черепа с роговыми стержнями описаны и из других местонахождений. У взрослых особей из пещеры Араго около Тотавеля во Франции поперечное сечение роговых стержней в 10 см и в 20 см от основания с уплощенным передним краем, в 20 см от основания оно округленно-треугольное (см. Rivals, 2004, рис. 23, 24 ). Индекс DT/DAP проксимального конца голотипа O. gracilis sp. nov. (78%) немного меньше, чем роговых стержней на расстоянии 10 см от основания O. ammon antiqua из Араго (80–89%). Ряд премоляров у O. ammon antiqua сокращен сильнее, чем у O. gracilis sp. nov. Коронка верхнего премоляра Р2 небольшая, короткая и более симметричная (см. Pommerol, 1880, рис. 69 ).
Замечания. Род Ovis обычно относят к трибе Caprini подсемейства Caprinae. Э. Крегу-Боннур выделила его в трибу Ovini этого подсемейства (Crégut-Bonnoure, 2007). В настоящее время часть исследователей включают Caprini с Ovis в подсемейство Antilopinae (s.l.), учитывая данные кладистики (напр., Gentry, 1992) и молекулярной генетики (напр., Hassanin, Douzery, 1999; Bibi et al., 2009; Wang et al., 2016).
Материал. Кроме голотипа, фрагмент правой верхнечелюстной кости с Р2–Р4, экз. ПИН, № 5644/309; левая верхнечелюстная кость с dP2–M2, экз. ПИН, № 5644/310; фрагмент левой ветви нижнечелюстной кости с M1–M3, экз. ПИН, № 5644/311; левая пястная кость, экз. ПИН, № 5644/312; правая плюсневая кость без дистального конца, экз. ПИН, № 5644/313; все из типового местонахождения.
О РАННЕЙ ИСТОРИИ РОДА ОVIS
Новые данные из Тавриды позволяют обсудить некоторые аспекты эволюции Ovis и их вселения в Европу в раннем плейстоцене и первой половине среднего плейстоцена, в интервале примерно от 2.6 до 0.4 млн лет. История Ovis в этом интервале протекала на фоне общего тренда нарастающего похолодания климата планеты, подъема горных сооружений Альпийско-Гималайского пояса, периодических горных оледенений, становления ледниковых и межледниковых циклов и смен ландшафтно-климатических обстановок. Эти горные животные одни из первых среди представителей плейстоценовой мегафауны Палеарктики начали приспосабливаться к обитанию в умеренном и холодном климате.
Эволюция и филогения. Основные направления эволюции Ovis связаны с адаптациями к обитанию в горных открытых биотопах, развитием грейзинга, а также совершенствованием формы рогов самцов, приспособленных для боя фронтальной поверхностью. Размеры рогов и их пневматизация (синусы) увеличивались. Увеличение рогов сопровождалось усилением их отгиба назад, развала и скручивания. Адаптации к грейзингу проявлялись в усилении гипсодонтности, сокращении цингулумов, уменьшении относительной длины ряда премоляров, увеличении и расширении моляров (особенно последних). Адаптации к передвижению в горах сопровождались сокращением относительной длины и увеличением массивности метаподий.
Палеонтологические данные позволяют предположить, что первые Ovis появились в Центральной Азии. Древнейшие Ovis известны из позднеплиоценовой (ранневиллафранкской) фауны Удунга (~3.5–3.0 млн л. н., MN16a) в Забайкалье (Россия) (Vislobokova et al., 1995; Вислобокова, 2008; русциний: Калмыков, 2013) и раннеплейстоценовых фаун Cяшагоу и Шаншенмиазуй (2.2–1.7 млн л. н.) бассейна Нихэвань (Китай) (Wang et al., 2016; Tong et al., 2022). Уже перед палеомагнитным эпизодом Олдувей (1.95–1.77 млн л. н.) ареал рода протягивался до южных районов Европы (рис. 4).
Новые данные из Тавриды показывают, что в раннем плейстоцене в Евразии существовали, по меньшей мере, две рано разошедшиеся филогенетические ветви рода Ovis, европейская с O. gracilis sp. nov. и азиатская с O. shantungensis (рис. 5). Виды O. shantungensis из Нихэваня и O. gracilis sp. nov. из Тавриды имеют некоторые общие черты (напр., сравнительно небольшие размеры, длинный ряд премоляров, особенности строения dP3 и P2, длинные метаподии), но характеризуются разным сочетанием примитивных и прогрессивных морфологических признаков. Большая прогрессивность O. shantungensis по сравнению с O. gracilis sp. nov. по ряду признаков (длине синусов, степени гипсодонтности, массивности метаподий) свидетельствует о достаточно ранней дивергенции этих ветвей, а также отражает сравнительно высокую скорость эволюции азиатской ветви. Родственные отношения этих видов с другими раннеплейстоценовыми Ovis и линиями современных видов этого рода пока не ясны. По уровню эволюционного развития O. shantungensis и O. gracilis sp. nov. примитивнее ранних O. ammon.
Рис. 5.
Хронологическое положение местонахождений и распространение представителей рода Ovis в раннем–среднем плейстоцене Евразии (c использованием данных: Tong et al., 1995; Rivals, 2004; Barychnikov, 2002; Spassov, 2005, 2016; Fernandez, Crégut, 2007; Kostopoulos et al., 2022; Tong et al., 2022).
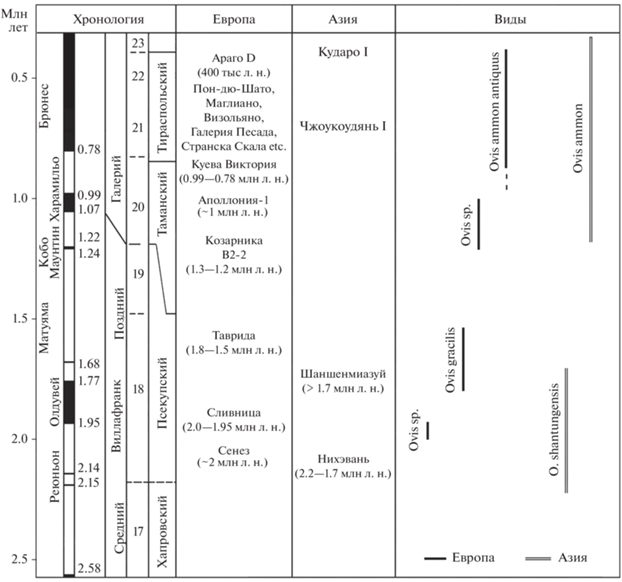
Сведений о морфологии Ovis из других местонахождений раннего плейстоцена Европы очень мало. Имеющиеся ископаемые остатки до вида неопределимы, и предлагаемые для них Э. Крегу видовые названия достаточно условны (см. Crégut-Bonnoure, 2007). Она считала, что в Сенезе (~2 млн л. н.) присутствует O. ammon ssp., а в Сливнице (~2 млн л. н.), Аполлонии-1 (~1 млн л. н.) и Куэва Виктории (0.99–0.78 млн л. н.) – O. ammon antiqua. Форма Ovis sp. из пещеры Козарника, по мнению Ф. Фернандеса и Крегу, имеет некоторое сходство с муфлоном и могла быть промежуточной между O. shantungensis и О. ammon antiqua (Fernandez, Crégut, 2007). По строению зубов бараны биозоны B2-2 пещеры Козарника (~1.3–1.2 млн л. н.) несомненно прогрессивнее, чем O. gracilis sp. nov., так как степень гипсодонтности у них выше, коронки премоляров короче, а третья доля на М3 длиннее и шире (см. Fernandez, Crégut, 2007, табл. III , фиг. 4). Следует подчеркнуть, что среди европейских раннеплейстоценовых форм могли быть потомки разных эволюционных линий, в т.ч. близкородственные видам О. gracilis sp. nov. и О. ammon.
Eвропейский аргали O. ammon antiqua (=O. antiqua) отличается от раннеплейстоценовых O. shantungensis и O. gracilis sp. nov. более крупными общими размерами, более массивными роговыми стержнями и сильнее сокращенным рядом премоляров. Oт O. shantungensis, кроме того, он отличается более сильным загибом роговых стержней назад и их более сильным отклонением в стороны (Pommerol, 1880, 1882; Rivals, 2004). O. ammon antiqua был широко распространен на юге Европы от Пиренейского п-ова до Карпат и Балкан в первой половине среднего плейстоцена, в интервале 0.55–0.4 млн л. н. (MIS15–11) (e.g., Rivals, 2004; Rozzi et al., 2011). Но, возможно, он впервые появился на этой территории раньше (Crégut-Bonnoure, 2007).
В Крыму фрагментарные остатки аргалиобразных и муфлонообразных баранов находили в пещерных местонахождениях позднего плейстоцена (напр., Аджи-Коба) (Громова, 1935; Громов, 1948). В Закавказье крупные аргалиобразные бараны встречались со второй половины среднего плейстоцена (напр., пещера Кударо 1, слой 5с) (напр., Верещагин, 1959; MIS10, ca. 0.360 млн л. н.: Barychnikov, 2002), а муфлоны O. orientalis присутствуют в позднем плейстоцене Северного Кавказа (Мезмайская и Монашеская пещеры) и Закавказья (Ахштырская пещера) (напр., Барышников, 2012). В Китае первое появление O. ammon относится к фазе гонванлин (~1.3 млн л. н.) позднего нихэвания (Tong et al., 1995). Остатки Ovis cf. O. ammon присутствуют в среднем плейстоцене Чжоукоудяня 1 около Пекина (Young, 1932; 0.69 млн л. н.: Qiu, 2006).
Данные филогенетики говорят о раннем расхождении линий, ведущих к современным видам, но оценка эволюционного уровня современных видов неоднозначна и в дальнейшем может быть пересмотрена с учетом палеонтологических и морфологических данных.
В современной фауне насчитывают от шести до 18 видов диких баранов рода Ovis (см. Groves, Grubb, 2011). Три вида или видовые группы – аргали O. ammon L., 1758, муфлона O. orientalis Gmelin, 1774 (=O. gmelini Blyth, 1841) и уриала O. vignei Blyth, 1841 – относятся к центрально-западноазиатской подгруппе (или кладе). Анализы, основанные на цитохроме b, полном митохондриальном геноме (mtDNA) и полном геноме, показывают, что наиболее низкую позицию в этой подгруппе занимает O. ammon (Hassanin et al., 1998; Bunch et al., 2006; Lv et al., 2015; Chen et al., 2021; Upadhyay et al., 2021). Кладистический анализ на основе морфологических признаков и цитохрома b предполагает более низкое положение O. ammon и O. vignei по сравнению с O. orientalis (Bibi et al., 2012). По данным кариосистематики, предковый кариотип у Ovis (n = 60), а кариотипы видов следующие: O. ammon (2n = 56), O. orientalis (n = 54), O. vignei (n = 58) (напр., Bunch et al., 2006). По индексу массивности роговых стержней, а также индексу длины мозгового отдела реконструируется моноклин, вершину которого занимает O. ammon (Groves, Grubb, 2011).
По молекулярным часам аргаливидные и муфлонообразные формы дивергировали от предкового ствола в раннем плейстоцене, но время, вычисленное разными методами на основе разных данных, различается. По одной из версий по цитохрому b время дивергенции аргаливидных форм от предкового ствола – 1.53 млн л. н., а муфлоновидных – 1.23 млн (Bunch et al., 2006). По другой версии O. ammon и O. orientalis дивергировали около 1.72 млн л. н. (Rezaei et al., 2010). Анализ полных геномов показал, что дивергенция линии O. ammon с другими линиями могла произойти 2.312 млн л. н. (Upadhyay et al., 2021), а анализ полных митогеномов понижает ее до 2.68–3 млн л. н. (Lv et al., 2015). Общий предок линий, ведущих к современным видам, мог существовать 3.12 млн л. н. (Bunch et al., 2006) или 3.813 млн л. н. (Upadhyay et al., 2021).
Происхождение. Вопрос о непосредственных предках рода Ovis остается открытым. Одна из последних гипотез предполагает, что предком Ovis мог быть Protovis himalayensis Wang, Li еt Takeuchi, 2016 из плиоцена местонахождения Гуаньцзинтай (5.46–3.10 млн л. н.) в Западных Гималаях Китая (Wang еt al., 2016).
Голотип P. himalayensis (часть крыши черепа с роговыми стержнями, IVPP V18928) и единственный паратип (фрагмент рога) хранятся в Ин-те палеонтологии позвоночных и палеоантропологии Академии наук Китая в Пекине (Wang et al., 2016, рис. 2, 3). Роговые стержни у Ovis и Protovis в целом похожи по форме, но у Protovis их строение примитивнее. Они менее утолщенные, с более коротким синусом и лишь с легким гомонимным скручиванием на концах. У голотипа в проксимальной части роговые стержни слабо отклоняются назад, но сильно расходятся в стороны (угол дивергенции 84°) (Wang et al., 2016). Поперечное сечение рогового стержня округленно-треугольное, с равными поперечным и переднезадним диаметрами (DT = DAP) в основании, тогда как у Ovis поперечный диаметр меньше переднезаднего и роговые стержни в средней части более сжаты медиолатерально. Синусы у Protovis c распорками, как у Ovis, но развиты лишь в вентральной части проксимальной половины рогового стержня (Wang et al., 2016, рис. 3A). У Ovis синусы развиты сильнее и у некоторых современных видов простираются почти до вершины. O. gracilis сходен с P. himalayensis по размерам, не очень утолщенным роговым стержням и слабому развитию синусов, но отличается более сильным дорсальным изгибом рогового стержня, его более резким сужением в дистальном направлении и более сильным гомонимным скручиванием, а также такими типичными для Ovis признаками, как характерная уплощенность фронтальной поверхности и выраженность “бойцовского” ребра. Не исключено, что эволюция Ovis и Protovis могла идти параллельно. Кладистические анализы Protovis допускают оба сценария: 1) сестринское родство в пределах клады Ovis или 2) его положение в основании клады Ovis (Wang et al., 2016, рис. 5). Судя по тому, что у современных видов Ovis представлены разные типы скручивания рогов (гомонимное, гетеронимное или в одной плоскости), предковая форма, вероятно, имела аркообразно загнутые назад рога без гомонимного или гетеронимного скручивания.
Некоторые исследователи предполагают, что близким родственником Ovis мог быть род Sinocapra Chen, 1991 (Mead, Taylor, 2005; Wang et al., 2016; Kostopoulos, 2022). Sinocapra присутствует в позднем плиоцене Юше в Китае (Chen, 1991; 3.6–2.59 млн л. н.: Qiu et al., 2013) и в нижнем плиоцене Невады в Северной Америке (4.95–4.5 млн л. н.: Mead, Taylor, 2005). Роговые стержни у Sinocapra небольшие, сравнительно короткие, почти прямые, очень слабо загнутые назад и без скручивания, овальные в сечении. У Sinocapra из Китая они сильно сжаты медиолатерально и больше похожи на роговые стержни козлов (Capra), а не баранов (Ovis) (Chen, 1991).
И.И. Соколов (1959) считал, что общим предком Ovis и Capra мог быть род Tossunnoria Bohlin, 1937 из позднемиоценовой фауны Таосу бассейна Цайдам на севере Тибетского плато.
Для подтверждения или опровержения этих гипотез нужны новые ископаемые материалы и исследования.
Распространение и экология. “Out of Tibet” модель распространения некоторых видов крупных млекопитающих плейстоценовой мегафауны хорошо подходит и для рода Ovis (Wang et al., 2016). Тибетское нагорье и горные системы Центральной и Средней Азии, по-видимому, были тем основным регионом Палеарктики, в котором происходила адаптация предковых форм и ранних Ovis к обитанию на все большей высоте, понижению температур и увеличению сезонности климата, и откуда началось их расселение на другие территории. Филогеография современных Ovis на основе митохондриальных и ядерных данных отражает последовательное видообразование на путях расселения (Rezaei et al., 2010).
Ранние Ovis распространялись на запад и в Европу по горным цепям Альпийского-Гималайского пояса. На путях их расселения могли возникать вторичные центры видообразования (напр., на юго-западе Азии, Армянском или Иранском нагорьях), откуда они проникали через Кавказ в Крым или через Малую Азию на Балканы и далее на запад. Последующее видообразование могло происходить и в Европе в условиях изоляции, при разрывах ареалов во время значительных изменений окружающей среды.
В плейстоцене хорошо прослеживаются дисперсионные волны крупных растительноядных млекопитающих из Азии в Европу в стадии крупных глобальных похолоданий и во время ледниковых эпох. С каждой новой волной во время похолоданий туда приходили формы, адаптированные к все более прохладному климату, а затем формы, приспособленные к обитанию в холодном климате. Вселение Ovis в Европу из Азии, вероятно, также могло происходить в раннем плейстоцене неоднократно, при наступлении подходящих ландшафтно-климатических условий. Как показали исследования А.Е. Додонова (2002), такие условия могли возникать при последовательных фазах горных оледенений, вызванных подъемом горных сооружений Центральной и Средней Азии (Тибета, Памира и Тянь-Шаня) на рубежах около 2.5, 2–1.8 и 0.8 млн л. н. При похолоданиях климат становился суше и континентальнее, с более выраженной сезонностью, широко распространялись открытые пространства.
Хронология местонахождений и данные по другим Artiodactyla позволяют предположить, что вселение ранних Ovis в Европу по горным системам из Азии могло быть связано с тремя основными дисперсионными волнами: 1) в первой половине раннего плейстоцена (~2 млн л. н.), перед палеомагнитным эпизодом Олдувей, 2) в конце раннего плейстоцена (~1.2 млн л. н.), на уровне эпизода Кобб Маунтин, и 3) на рубеже раннего и среднего плейстоцена (~0.8 млн л. н.), на уровне инверсии Брюнес/Матуяма (напр., Вислобокова, 2022). С этими волнами в Европу приходили формы, вероятно, все более адаптированные к жизни в открытых горных биотопах и во все более прохладном климате. Этот тренд обусловлен изменениями палеообстановок в связи с продолжавшимся подъемом горных сооружений и глобальными похолоданиями. Адаптации к обитанию в таких условиях способствовали тому, что ареалы центральноазиатских форм значительно увеличивались при крупных глобальных похолоданиях, вызывавших перестройки и расширение соответствующих биомов. Из центральных районов Палеарктики по пологим склонам горных цепей и предгорьям, вероятно, через экотоны, такие формы могли распространяться не только к северу, но и в восточные регионы Западной Палеарктики. Приход европейских аргали в Европу мог быть связан со второй или третьей дисперсионными волнами, а адаптированных к менее прохладному климату, возможно, муфлонообразных форм – с первой волной.
Раннеплейстоценовые бараны O. gracilis sp. nov., судя по строению зубной системы, степени гипсодонтности и характеру мезостирания, вероятно, были смешанноядными, с сохранением некоторых черт браузеров. Они были менее приспособлены к грейзингу, чем O. ammon antiqua начала среднего плейстоцена и, по-видимому, были более теплолюбивыми. По своим ландшафтно-климатическим адаптациям O. gracilis sp. nov., вероятно, был ближе к современным O. orientalis (=O. gmelini), чем к O. ammon. Вид O. orientalis обитает как в умеренном климате, так и в субтропиках, в разных биотопах, от лесов до степей (напр., Michel, Ghoddousi, 2020a). Бараны вида O. gracilis sp. nov. могли предпочитать пологие склоны с мозаичными ландшафтами, открытые пространства с кустарниками или участками древесной растительности (в т.ч. саванны). Европейский аргали O. ammon antiqua по своим адаптациям, по-видимому, был близок к современному O. ammon. Экоморфологический анализ и анализ микроизноса зубов барана из пещеры Араго (уровень D, 400 тыс. л. н., MIS 12) на востоке Пиренеев показали, что он предпочитал пологие склоны и открытые пространства, был преимущественно грейзером, значительную долю в его рационе составляли травы (Rivals, Deniaux, 2003; Rivals, 2004). В Европе O. ammon antiqua был более обычен в холодные фазы, но, по-видимому, мог переносить и некоторые глобальные климатические флуктуации (Rivals et al., 2008; Rozzi et al., 2011). Характер мезоизноса зубов у этого вида из Араго в холодные и сухие климатические фазы (MIS 12 и 14) соответствовал грейзерам, а в теплые гумидные (MIS 13) – смешанноядным (Rivals et al., 2008).
ЗАКЛЮЧЕНИЕ
Таким образом, на основании изучения материалов из пещеры Таврида, в т.ч. с применением томографии, установлено присутствие нового вида Ovis gracilis в раннем плейстоцене Крыма (1.8–1.5 млн л. н.). Вселение Ovis в Европу в раннем плейстоцене – начале среднего плейстоцена, очевидно, шло по горным сооружениям Альпийско-Гималайского пояса и связано с дисперсионными волнами из Азии. Первые Ovis могли прийти в Крым из Юго-Западной Азии через Кавказ перед палеомагнитным эпизодом Олдувей.
Список литературы
Барышников Г.Ф. Обзор ископаемых остатков позвоночных из плейстоценовых слоев Ахштырской пещеры (Северо-Западный Кавказ) // Тр. Зоол. ин-та РАН. 2012. Т. 316. № 2. С. 93–138.
Верещагин Н.К. Млекопитающие Кавказа. История формирования фауны. М.–Л.: Изд-во АН СССР, 1959. 704 с.
Вислобокова И.А. Основные этапы эволюции сообществ парнопалых Северной Евразии в плиоцене–начале среднего плейстоцена. Часть 1 // Палеонтол. журн. 2008. № 3. С. 76–91.
Вислобокова И.А. О первой находке Soergelia minor (Artiodactyla, Bovidae) в нижнем плейстоцене пещеры Таврида в Крыму и истории рода Soergelia // Палеонтол. журн. 2022. № 3. С. 61–70.
Громов В.И. Палеонтологическое и археологическое обоснование стратиграфии континентальных отложений четвертичного периода на территории СССР (млекопитающие, палеолит). М.: Изд-во АН СССР, 1948. 522 с.
Громова В.И. Об остатках диких баранов и козлов в четвертичных отложениях Крыма // Докл. АН СССР. 1935. Т. 4. № 1–2. С. 97–100.
Громова В.И. Остеологические отличия родов Capra (козлы) и Ovis (бараны). М.: Изд-во АН СССР, 1953. 124 с.
Додонов А.Е. Четвертичный период Средней Азии. М.: ГЕОС, 2002. 250 с.
Калмыков Н.П. Самая древняя находка горного барана (Mammalia, Artiodactyla: Ovis) в Голарктике (Западное Забайкалье) // Докл. Акад. наук. 2013. Т. 448. № 2. С. 239–241.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Докл. Акад. наук. 2019. Т. 485. № 3. С. 381–385.
Соколов И.И. Копытные звери (Отряды Perissodactyla и Artiodactyla). М.–Л.: Изд-во АН СССР, 1959. 640 с.
Barychnikov G. Local biochronology of Middle and Late Pleistocene mammals from the Caucasus // Russ. J. Theriol. 2002. V. 1. № 1. P. 61–67.
Bibi F., Vrba E., Fack F. A new African fossil caprin and a combined molecular and morphological Bayesian phylogenetic analysis of caprini (Mammalia: Bovidae) // J. Evol. Biol. 2012. V. 25. P. 1843–1854.
Bibi F., Bukhsianidze M., Gentry A.W. et al. The fossil record and evolution of Bovidae: State of the field // Palaeontol. Electron. 2009. V. 12. Iss. 3. 10A. P. 1–10.
Bohlin B. Einige Jungtertiäre und Pleistozäne Cavicornier aus Nord-China // Nova Acta Reg. Soc. Sci. Upsal. Ser. IV. 1938. V. 11. № 2. P. 1–54.
Bunch T.D., Wu C., Zhang Y.-P., Wang S. Phylogenetic analysis of snow sheep (Ovis nivicola) and closely related taxa // J. Heredity. 2006. V. 97. № 1. P. 21–30.
Chen G. A new genus of Caprini (Bovidae, Artiodactyla) from Upper Pliocene of Yushe, Shansi // Vertebr. PalAsiat. 1991. V. 29. P. 230–239.
Chen Z.-H., Xu Y.-H., Xie X.-L. et al. Whole-genome sequence analysis unveils different origins of European and Asiatic mouflon and domestication-related genes in sheep // Communication Biol. 2021. V. 4. № 1307. P. 1–15.
Crégut-Bonnoure E. Apport des Caprinae et Antilopinae (Mammalia, Bovidae) à la biostratigraphie du Pliocène terminal et du Pléistocène d’Europe // Quaternaire. 2007. V. 18. № 1. P. 73–97.
Drake A., Donahue T.L.H., Stansloski M. et al. Horn and horn core trabecular bone of bighorn sheep rams absorbs impact energy and reduces brain cavity accelerations during high impact ramming of the skull // Acta Biomaterialia. 2016. V. 44. P. 41–50.
Gentry A.W. The subfamilies and tribes of the family Bovidae // Mammal Rev. 1992. V. 22 P. 1–32.
Groves C., Grubb P. Ungulate taxonomy. Baltimore: John Hopkins Univ. Press, 2011. 310 p.
Farke A.D. Evolution and functional morphology of the frontal sinuses in Bovidae (Mammalia: Artiodactyla), and implications for the evolution of cranial pneumaticity // Zool. J. Linn. Soc. 2010. V. 159. P. 988–1014.
Fernandez P., Crégut E. Les Caprinae (Rupicaprini, Ovibovini, Ovini et Caprini) de la séquence pléistocène de Kozarnika (Bulgarie du Nord): morphométrie, biochronologie et implications phylogéniques // Rev. Paléobiol. 2007. V. 26. № 2. P. 425–503.
Harris R.B., Tsytsulina K. Ovis nivicola // The IUCN Red List of Threatened Species. 2008. e.T15740A5076357. 9 p.
Hassanin A., Douzery E.J. The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome b gene // Mol. Phylog. Evol. 1999. V. 13. P. 227–243.
Hassanin A., Pasquet E., Vigne J.-D. Molecular systematics of the subfamily Caprinae (Artiodactyla, Bovidae) as determined from cytochrome b sequences // J. Mammal. Evol. 1998. V. 5. P. 217–236.
Kostopoulos D.S. The fossil record of bovids (Mammalia: Artiodactyla: Ruminantia: Pecora: Bovidae) in Greece // Fossil vertebrates of Greece Vol. 2. Laurasiatherians, Artiodactyles, Perissodactyles, Carnivorans, and island endemics / Ed. Vlachos E. Springer, 2022. P. 113–204.
Lv F.-H., Peng W.-F., Yang J. et al. Mitogenomic meta-analysis identifies two phases of migration in the history of eastern Eurasian sheep // Mol. Biol. Evol. 2015. V. 139. P. 1–19.
Mead J.I., Taylor L.H. New species of Sinocapra (Bovidae, Caprinae) from the Lower Pliocene Panaca Formation, Nevada, USA // Palaeontol. Electron. 2005. V. 8. 11A. 20 p.
Matsumoto H. On a new fossil race of bighorn sheep from Shantung, China // Sci. Rep. Tôhoku Imp. Univ. Ser 2. 1926. V. 10. № 2. P. 38–41.
Michel S., Ghoddousi A. Ovis gmelini // The IUCN Red List of Threatened Species. 2020a. e.T54940218A22147055. 16 p.
Michel S., Ghoddousi A. Ovis vignei // The IUCN Red List of Threatened Species. 2020b. e.T54940655A195296049. 28 p.
Teilhard de Chardin P., Piveteau J. Les mammiféres fossils de Nihowan (Chine) // Ann. Paléontol. 1930. V. 19. P. 1–134.
Pommerol F. Le mouflon quaternaire // Congrès de l’Association Française pour l’Avancement des Sciences. C. R. 8e sess., Montpellier, 1879. P., 1880. P. 600–609.
Qiu Z.-X. Quaternary environmental changes and evolution of large mammals in North China // Vertebr. PalAsiat. 2006. V. 44. № 2. P. 109–132.
Qiu Z.-X., Qiu Z.-D., Deng T. Neogene Land Mammal Stages/Ages of China: Toward the goal to establish an Asian Land Mammal Stage/Age scheme // Fossil Mammals of Asia / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 29–90.
Reading R., Michel S., Amgalanbaatar S. Ovis ammon // The IUCN Red List of Threatened Species. 2020. e.T15733A22146397. 32 p.
Rezaei H.R., Naderi S., Chintauan-Marquier I.C. et al. Evolution and taxonomy of the wild species of the genus Ovis (Mammalia, Artiodactyla, Bovidae) // Mol. Phylog. Evol. 2010. V. 54. P. 315–326.
Rivals F. Les petits bovidés (Caprini et Rupicaprini) pléistocènes dans le bassin méditerranéen et le Caucase: étude paléontologique, biostratigraphique, archéozoologique et paléoécologique. Oxford: Archeopress, 2004. 252 p.
Rivals F., Deniaux B. Dental microwear analysis for investigating the diet of an argali population (Ovis ammon antiqua) of mid-Pleistocene age, Caune de l’Arago cave, eastern Pyrenees, France // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2003. V. 193. P. 443–455.
Rivals F., Schulz E., Kaiser T.M. Climate related dietary diversity of the ungulate faunas from the middle Pleistocene succession (OIS 14–12) at the Caune de l’Arago (France) // Paleobiology. 2008. V. 34. P. 117–127.
Rozzi R., Palombo M.R., Barbieri M. The argali (Ovis ammon antiqua) from the Magliana area (Rome) // Il Quaternario. 2011. V. 24. № 1. P. 113–119.
Schaffer W.M., Reed C.A. The co-evolution of social behavior and cranial morphology in sheep and goats (Bovidae, Caprini) // Fieldiana. 1972. V. 61. P. 1–88.
Schaub S. Die oberpliocaene Säugetierfauna von Senèze (Haute-Loire) und ihre verbreitungsgeschichtliche Stellung // Ecl. Geol. Helv. 1943. Bd 36. Hf. 2. S. 270–289.
Spassov N. Brief review of the Pliocene ungulate fauna of Bulgaria // Quaternaire. 2005. V. 2. P. 201–212.
Spassov N. Southeastern Europe as a route for the earliest dispersal of Homo toward Europe: ecological conditions and the timing of the first human occupation of Europe // Paleoanthropology of the Balkans and Anatolia: Human Evolution and its Context / Eds. Harvati K., Roksandic M. Dordrecht: Springer, 2016. P. 281–290.
Spassov N., Crégut-Bonnoure E. Premières données sur les Bovidae Villafranchiens de Bulgarie // C. R. Acad. Sci. Paris. 1999. V. 328. P. 493–498.
Teilhard de Chardin P., Piveteau J. Les mammiféres fossils de Nihowan (Chine) // Ann. Paléontol. 1930. V. 19. P. 1–134.
Tong H.-W., Zhang B., Chen X. et al. New fossils of small and medium-sized bovids from the Early Pleistocene site of Shanshenmiaozui in Nihewan Basin, North China // Vertebr. PalAsiat. 2022. V. 60. № 2. P. 134–168.
Tong Y., Zheng S., Qiu Z. Cenozoic mammal ages of China // Vertebr. PalAsiat. 1995. V. 33. № 4. P. 290–314.
Upadhyay M., Kunz E., Sandoval-Castellano E. et al. Whole genome sequencing reveals a complex introgression history and the basis of adaptation to subarctic climate in wild sheep // Mol. Ecol. 2021. V. 30. № 24. P. 6701–6717.
Vislobokova I., Dmitrieva E., Kalmykov N. Artiodactyls from the Late Pliocene of Udunga, Western Trans-Baikal, Russia // J. Vertebr. Paleontol. 1995. V. 15. № 1. P. 146–159.
Vrba E.S., Schaller G.B. Phylogeny of Bovidae (Mammalia) based on behavior, glands and skull morphology // Antelopes, deer, and relatives: fossil record, behavioral ecology, systematics, and conservation / Eds. Vrba E.S., Schaller G.B. New Haven, CT: Yale Univ. Press, 2000. P. 203–222.
Wang X., Li Q., Takeuchi G.T. Out of Tibet: an early sheep from the Pliocene of Tibet, Protovis himalayensis, gen. et sp. nov. (Bovidae, Caprini), and origin of Ice Age mountain sheep // J. Vertebr. Paleontol. 2016. e1169190. 12 p.
Young C.-C. On the Artiodactyla from the Sinathropus site at Chouk’outien // Palaeontol. Sin. Ser. C. 1932. V. 8. P. 1–158.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал