

Почвоведение, 2019, № 11, стр. 1295-1306
Сравнительная характеристика современных и погребенных почвенных комплексов в пустынно-степной зоне Волго-Донского междуречья
Т. С. Демкина a, *, А. В. Борисов a, Т. Э. Хомутова a
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, 2, Россия
* E-mail: demkina-t@rambler.ru
Поступила в редакцию 09.04.2018
После доработки 15.11.2018
Принята к публикации 26.12.2018
Аннотация
Проведены исследования древних (XXVII–XXVI вв. до н. э.) и современных почвенных комплексов на низкой и высокой надпойменных террасах р. Джурак-Сал и на водораздельном участке в пределах Сальско-Манычской гряды пустынно-степной зоны (юго-восток Восточно-Европейской равнины). Выполнены морфолого-генетические, физико-химические и микробиологические анализы почв. Установлено, что наиболее контрастные почвенные комплексы были развиты в эпоху бронзы на низких надпойменных террасах, наименее контрастные – на водораздельных участках. Микробные сообщества почв в погребенных и современных комплексах в меньшей степени зависели от особенностей проявления солонцового процесса. Суммарная численность микроорганизмов, растущих на почвенном агаре и использующих элементы питания из рассеянного состояния, растущих на нитритном агаре и потребляющих гумус, растущих на богатой среде и разлагающих растительные остатки, в погребенных и современных почвах довольно близка в пределах каждого ключевого участка, но существенно различается на разных элементах рельефа. При этом и в светло-каштановых почвах (Eutric Cambisols (Loamic, Protocalcic)), и в солонцах (Haplic Solonetz (Loamic, Cutanic, Ochric)) сохраняются закономерности в соотношении микроорганизмов разных трофических групп, отражающие климатические условия, имевшие место в древности и в настоящее время.
ВВЕДЕНИЕ
Изучение солонцов в структуре почвенного покрова пустынно-степной зоны, их физико-химических свойств и генезиса началось с фундаментальных работ К.К. Гедройца в 1910-х гг., активизировалось в середине и второй половине XX в. и продолжается до наших дней [3–5, 11, 16–18, 20–23, 25, 28–31, 34]. Столь пристальное внимание к солонцовому процессу обусловило очень глубокую теоретическую и практическую изученность этого феномена. Менее проработана временная составляющая солонцового процесса. В частности, показано, что в восточно-европейских степях солонцовый процесс получил развитие лишь в суббореальном периоде голоцена, причем в различных географических зонах этот процесс возникал асинхронно в зависимости от геоморфологических и литологических условий [6, 11, 12, 15]. Итоги изучения вопросов, связанных с возникновением солонцового процесса в южнорусских степях, были сформулированы Демкиным в 1997 г. [9]. По мнению автора, на большей части степной зоны солонцеватость появилась в эпоху средней и поздней бронзы (II тыс. до н. э.). Несколько позднее, во второй половине II тыс. до н. э. солонцовый процесс начался в почвах недренированных равнин, ранее всего признаки солонцового процесса выявлены в почвах восточного склона Ергенинской возвышенности и Приволжской полосы [8, 9]. С того времени солонцовый процесс в той или иной мере проявлялся в почвах пустынно-степной зоны на протяжении всех исторических этапов, кроме рубежа 3–2 тыс. до н. э., когда в условиях резкой аридизации широкое распространение получили каштановидные карбонатные засоленные почвы без признаков солонцового процесса [10].
Изучению свойств погребенных и современных почв степной зоны посвящен большой объем литературы. Однако наблюдений за изменением свойств погребенных и современных почвенных комплексов мало [13]. Это связано с тем, что редко встречаются подкурганные погребенные комплексы хорошей сохранности, позволяющие выделить почвы, составляющие комплексы, и проследить изменение их свойств. Что касается исследований почвенных комплексов одного времени погребения на разных элементах рельефа, то подобных работ до настоящего времени не было. В настоящей статье впервые предпринята попытка оценить морфологические, физико-химические и микробиологические свойства погребенных и современных почвенных комплексов на низкой и высокой речных террасах и локальном водоразделе.
ОБЪЕКТЫ И МЕТОДЫ
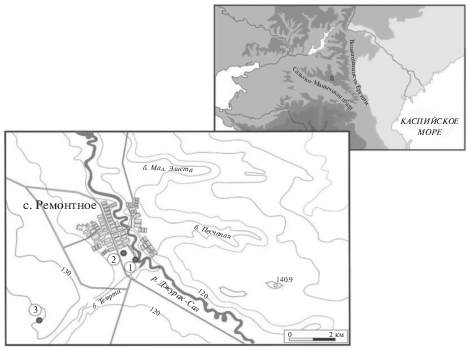
В качестве объектов исследования погребенных и современных почвенных комплексов выбраны подкурганные почвы и их современные аналоги на территории Ремонтненского района Ростовской области. Эта территории лежит в пределах Сальско-Манычской гряды и представляет собой юго-восточную часть Восточно-Европейской равнины (рис. 1).
Рис. 1.
Район, местоположение объектов (курганные могильники) исследований: 1 – Темрта-5, 2 – Темрта-3, 3 – Темрта-1.
Климат района умеренно континентальный. Среднегодовая норма атмосферных осадков 300–350 мм, из них на летний период приходится от 120 до 220 мм. Наибольшее количество осадков выпадет в июне 25–40 мм. Снежный покров формируется в третьей декаде ноября. Высота снежного покрова от 3–8 до 10–12 см. Характерны оттепели, когда температура поднимается до 5–10°С. Рельеф полого-волнистый, поверхность имеет развитую овражно-балочную сеть, что обеспечивает хорошую дренированность территории. Почвообразующие породы представлены лёссовидными карбонатными засоленными суглинками.
В геоморфологическом плане исследованные объекты располагались в следующем порядке: могильник Темрта-5 – на низкой надпойменной террасе р. Джурак-Сал (поверхность почвы в 3–4 м над урезом воды), могильник Темрта-3 – в 300 м выше по склону на слабовыраженной второй террасе при уровне залегания грунтовых вод в пределах 5–8 м, могильник Темрта-1 – на вершине локального водораздела с уровнем залегания грунтовых вод более 10 м. Исследованы погребенные почвы под курганами раннекатакомбной культуры, датируемые XXVII–XXVI вв. до н. э. (калиброванные даты). Высота курганов составляла около 1.2–1.5 м, диаметр – 30–40 м. При раскопках насыпь кургана разрезали серией траншей глубиной до гор. ВС, при этом оставляли 4–5 бровок, на которых можно было с двух сторон исследовать погребенную почву. Общая площадь погребенной почвы, которая вскрывается под курганом, составляла до 500 м2. Это позволило получить полное представление о структуре почвенного покрова и выбрать участки, где наиболее четко происходит смена почв комплексов. В таких местах закладывали почвенный разрез в виде траншеи, длиной, достаточной для охвата двух наиболее контрастных почвенных разностей, для которых проводили описание профилей и отбор образцов для химического и микробиологического исследований (рис. 2).
Рис. 2.
Курганная насыпь (А) и траншея с погребенной почвой (Б). В траншее вскрыт солонец корковый (в левой части) и светло-каштановая солонцеватая почва (справа). Линией показана верхняя граница погребенной почвы.

Изучение современного почвенного покрова проводили в пределах каждого объекта. Выбор места заложения разрезов современных почв определяли по характеру растительности таким образом, чтобы солонцовое пятно с чернополынно-прутняковой растительностью максимально резко сменялось светло-каштановыми почвами с ковыльково-мятликово-ромашниковыми растительными ассоциациями наибольшего проективного покрытия. В этом месте закладывали разрез, длина которого позволяла в одной фронтальной стенке описать солонец, в другой – светло-каштановую почву. По боковым стенкам определяли характер перехода одной почвенной разности в другую. Правильность местоположения опорного разреза современных почв проверяли серией прикопок.
В образцах, отобранных по генетическим горизонтам, выполняли определение содержания органического углерода методом мокрого сжигания по Тюрину, содержание карбонатов – ацидометрически, содержание гипса гравиметрическим методом при извлечении из почвы 0.2 н HCl, анионно-катионный состав водной вытяжки при соотношении почва : вода 1 : 5, величины рН потенциометрически. Гранулометрический состав определяли пирофосфатной подготовкой образца [1] пипеточным методом [26]11.
Следует отметить, что в результате проникновения солей из насыпи в погребенную почву происходит ее диагенетическое засоление, и величина плотного остатка в верхних горизонтах может превышать 1.5–2.0%. Следовательно, использовать физико-химические методы оценки развития солонцового процесса невозможно, так как будут заведомо искажены результаты определения содержания обменного натрия [19] и физико-химические условия развития солонцового процесса [29, 30].
Поэтому в настоящей работе в качестве диагностических критериев наличия солонцового процесса в почвах использовали морфологические характеристики верхней части профиля – осветление и листовато-слоистая структура в гор. А1, призматическая структура в гор. В1 с выраженным превышением вертикального размера призм над горизонтальным, наличие органо-глинистых кутан на поверхности структурных отдельностей [29, 30], а также текстурная дифференциация верхней части профиля: отношение содержания ила в гор. В1/А1 как дополнительный диагностический признак, показывающий глубину трансформации почвенной массы под действием солонцового процесса [7, 24]. Также учитывали характер нижней границы и переход между гор. А1 и В1, наличие или отсутствие подсолонцового гор. В2.
На микробиологические анализы отбирали образцы с соблюдением условий стерильности. Перед проведением анализов почвенные образцы просеивали через сито с диаметром ячеек 3 мм для удаления корней и усредняли. Содержание углерода активной микробной биомассы рассчитывали по скорости субстрат-индуцированного дыхания с использованием коэффициента пересчета 40.04 [35]. Живую микробную биомассу определяли по содержанию почвенных фосфолипидов (Ф-фл) [33], для пересчета данных в единицы углерода использовали коэффициент 190 мкмоль Ф-фл/г С [32]. Численность микроорганизмов разных трофических групп, характеризующих увлажненность климата сухо-степной зоны, определяли на богатой органической среде с пептоном (БС), почвенном агаре (ПА) и нитритном агаре (НА) [27] чашечным методом по общепринятой методике [14]. Для выявления различий по активности и устойчивости микробных сообществ современных и погребенных почв измеряли скорости базального (Vбазал) и субстрат-индуцированного дыхания (VСИД), рассчитывали коэффициент дыхательной активности QR = Vбазал/VСИД, который дает представление об устойчивости микробных сообществ, нарушение последнего сопряжено с увеличением этого коэффициента [2].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфолого-генетические и физико-химические свойства исследованных подкурганных и современных почв солонцового комплекса
Первая надпойменная терраса. Ключевой участок Темрта-5. Погребенная почва исследована под насыпью кургана высотой более 1.5 м. Курган располагался на первой надпойменной террасе р. Джурак-Сал. Сохранность погребенной почвы очень хорошая на всем протяжении оставленных бровок.
Для подкурганных почв характерна резко выраженная комплексность, причем под западной (рис. 3.1 ) и восточной (рис. 3.2 ) бровками кургана картина комплексов заметно отличалась. На фоне общего доминирования солонцов выделяются участки, где гор. А1 почв практически отсутствовал, либо его мощность составляла 1–2 см. В таких местах развивались особенно выраженные солонцы с палисадной нижней границей гор. А1, и округлыми “головками” блоков-тумб иллювиального горизонта. Как правило, такие солонцы приурочены к микропонижениям рельефа, при перепаде высот 2–3 см.
Рис. 3.
Ключевой участок Темрта-5, первая надпойменная терраса. Профили погребенных почв XXVII–XXVI вв. до н. э. в восточной (1) и западной (2) бровках кургана и современных почв (3); а – листовато-слоистая структура в гор. А1; б – призматическая структура в гор. В1; в – номер разреза; г – индекс горизонта; д – высота поверхности от условного нуля (для каждого участка).
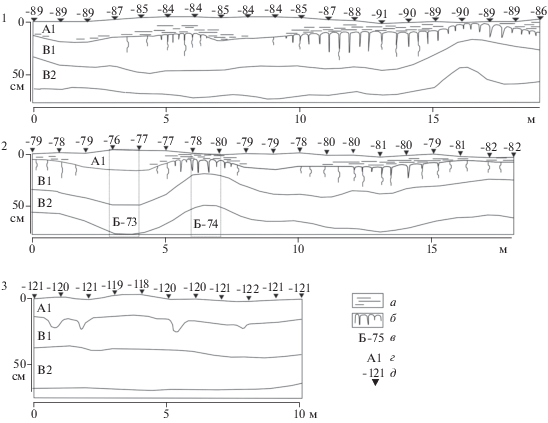
Формирование бронирующего гор. В1 изменило общую организацию почвенного профиля, стало причиной сокращения мощности верхних горизонтов профиля и изменения химических свойств. Под солонцами заметно более близкое к поверхности залегание солевых и гипсовых аккумуляций. Несмотря на сильный солевой диагенез по глубине залегания солевых новообразований, можно говорить, что под солонцами верхняя граница солевого горизонта на глубине 50 см, под светло-каштановыми почвами – на глубине 75 см. Но эта особенность проявляется локально только на тех участках, где солонцовый процесс начался раньше (восточная бровка, участок 16–17 м). Кроме того, выявлены солонцы на начальных этапах солонцового процесса (восточная бровка, участок 5–6 м) на участках без заметных микропонижений.
Солонцовый процесс в той или иной мере проявляется и во всех вскрытых светло-каштановых почвах: на всем протяжении бровок в нижней части гор. А1 заметна листоватая структура, а в гор. В1 хорошо видны кутаны иллювиирования и вертикальная ориентировка структурных отдельностей.
Для исследований были отобраны образцы светло-каштановой солонцеватой почвы (разр. Б-73) и солонца мелкого (разр. Б-74). Расстояние между двумя почвенными профилями не более 1.5 м. Химические свойства почв близки (табл. 1). Но уже хорошо видна текстурная дифференциация профиля солонцов – отношение ила в гор. В1/А1 солонца – 7.0, а в светло-каштановой солонцеватой почве – 2.4. Судя по выраженности солонцового процесса в почвах, можно говорить о тренде на активное развитие солонцового процесса в момент сооружения кургана (XXVII–XXVI вв. до н. э.).
Таблица 1.
Физико-химические свойства подкурганных и современных почвенных комплексов на разных элементах рельефа
| Горизонт, глубина, см | рН H2O | Гумус | Гипс | СаСО3 | Сумма солей | Содержание частиц, % | |
|---|---|---|---|---|---|---|---|
| % | <0.01 мм | <0.001 мм | |||||
| Первая надпойменная терраса, Темрта-5 | |||||||
| Погребенные почвы: | |||||||
| Светло-каштановая солонцеватая (Б-73) | |||||||
| А1, 0–9 | 8.6 | 0.84 | – | 1.5 | 2.07 | 33 | 14 |
| В1, 9–25 | 8.4 | 0.91 | – | 5.1 | 2.09 | 55 | 34 |
| В2, 25–40 | 9.0 | 0.89 | – | 18.8 | 1.97 | 58 | 28 |
| ВС, 40–75 | 9.3 | 0.98 | 0.8 | 17.2 | 2.12 | 50 | 23 |
| Сs, г, 75–200 | 8.8 | 0.98 | 2.2 | 7.8 | 3.11 | – | – |
| Солонец мелкий солончаковатый (Б-74) | |||||||
| А1, 0–5 | 8.7 | 0.90 | – | 1.4 | 1.67 | 23 | 4 |
| В1, 5–22 | 8.7 | 1.07 | – | 2.4 | 1.58 | 53 | 28 |
| В2, 22–30 | 8.8 | 1.00 | – | 13.2 | 2.61 | 44 | – |
| ВС, 30–50 | 8.8 | 0.82 | 0.9 | 15.6 | 2.25 | 41 | – |
| Сs, г, 50–200 | 8.8 | 0.65 | 4.9 | 8.0 | 2.91 | – | – |
| Современная почва: | |||||||
| Светло-каштановая несолонцеватая солончаковая (Б-75) | |||||||
| А1, 0–13 | 7.9 | 3.76 | – | 2.9 | 0.10 | 20 | 5 |
| В1, 13–40 | 9.1 | 1.95 | – | 8.7 | 0.17 | 41 | 19 |
| В2, 40–70 | 8.4 | 0.91 | – | 9.4 | 1.01 | 43 | 28 |
| Сs, г, 70–160 | 8.8 | 0.84 | 7.8 | 12.0 | 2.83 | – | – |
| Сг, 160–200 | 8.9 | 0.81 | 10.2 | 9.2 | 2.39 | – | – |
| Вторая надпойменная терраса, Темрта-3 | |||||||
| Погребенные почвы: | |||||||
| Светло-каштановая (Б-43) | |||||||
| А1, 0–20 | 9.1 | 0.83 | 0.6 | 3.5 | 1.81 | 42 | 21 |
| В1, 20–36 | 8.2 | 0.77 | 1.5 | 6.0 | 1.82 | 60 | 34 |
| В2, 36–44 | 8.8 | 0.97 | 2.1 | 12.3 | 1.87 | 60 | 34 |
| ВССа S, 44–128 | 8.4 | – | 3.4 | 11.2 | 2.76 | 52 | 26 |
| С, 128–200 | 8.9 | – | 0.6 | 13.0 | 1.60 | 41 | 22 |
| Солонец мелкий (Б-44) | |||||||
| А1, 0–6 | 7.5 | 0.67 | 0.6 | 0.9 | 2.65 | 41 | 12 |
| В1, 6–26 | 7.7 | 0.78 | 0.4 | 2.3 | 1.99 | 57 | 34 |
| В2, 26–34 | 7.7 | 0.88 | 0.3 | 2.2 | 2.01 | 63 | 42 |
| ВССа S, 34–80 | 7.7 | – | 2.2 | 11.8 | 2.42 | 59 | 34 |
| Сs, г, 80–130 | 7.8 | – | 0.7 | 13.6 | 1.50 | 58 | 38 |
| С, 130–200 | 7.7 | – | 1.1 | 13.8 | 1.77 | 49 | 17 |
| Современные почвы: | |||||||
| Светло-каштановая незасоленная (Б-107-6) | |||||||
| А1, 0–27 | 7.2 | 1.70 | – | – | – | 35 | 20 |
| В1, 27–47 | 7.9 | 1.50 | – | 1.5 | 0.02 | 43 | 28 |
| В2, 47–62 | 8.3 | 1.00 | – | 8.7 | 0.02 | 46 | 27 |
| ВС, 62–80 | 8.3 | 0.31 | 0.1 | 13.9 | 0.23 | 42 | 22 |
| С, 80–200 | 8.1 | – | 0.4 | 6.7 | 0.22 | 41 | 22 |
| Солонец средний (Б-107-0) | |||||||
| А1, 0–9 | 7.6 | 2.78 | – | 0.3 | – | 30 | 13 |
| В1, 9–24 | 8.4 | 1.50 | – | 1.5 | 0.22 | 52 | 37 |
| В2, 24–37 | 9.1 | 0.89 | – | 9.2 | 0.16 | 47 | 31 |
| ВС, 37–75 | 8.6 | 0.50 | 0.2 | 12.1 | 0.85 | 43 | 24 |
| Сs, г, 75–200 | 8.5 | – | 2.8 | 7.9 | 1.82 | 34 | – |
| Водораздел, Темрта-1 | |||||||
| Погребенные почвы: | |||||||
| Светло-каштановая (Б-104-К) | |||||||
| А1, 0–13 | 8.5 | 0.79 | – | 1.9 | 0.87 | 39 | 16 |
| В1, 13–35 | 8.5 | 0.69 | – | 2.6 | 0.52 | 48 | 26 |
| В2, 35–60 | 8.1 | 0.55 | – | 10.8 | 0.53 | 48 | 27 |
| ВС, 60–90 | 8.0 | 0.31 | – | 10.9 | 0.92 | 41 | 19 |
| ВС2, 90–125 | 8.0 | 0.24 | 0.3 | 7.5 | 0.62 | 45 | 19 |
| Сs, г, 125–160 | 8.0 | – | 2.5 | 10.1 | 1.44 | 50 | 23 |
| С, 160–180 | 8.3 | – | 1.0 | 10.2 | 0.93 | 51 | 26 |
| Солонец мелкий (Б-104-С) | |||||||
| А1, 0–6 | 7.9 | 0.60 | – | 0.5 | 0.58 | 35 | 14 |
| В1, 6–30 | 8.2 | 0.79 | – | 1.3 | 0.35 | 47 | 27 |
| В2, 30–55 | 8.2 | 0.45 | – | 12.7 | 0.57 | 48 | 27 |
| ВС, 55–100 | 8.1 | 0.39 | 0.3 | 11.1 | 0.59 | 43 | 23 |
| Сs, г, 100–135 | 7.9 | – | 6.1 | 7.2 | 1.94 | 37 | 24 |
| С, 135–180 | 8.3 | – | 0.3 | 9.3 | 0.72 | 50 | 24 |
| Современные почвы: | |||||||
| Светло-каштановая солонцеватая (Б-105-К) | |||||||
| А1, 0–17 | 7.1 | 1.77 | – | 0.5 | 0.04 | 30 | 15 |
| В1, 17–38 | 7.6 | 0.99 | – | 1.3 | 0.07 | 49 | 34 |
| В2, 38–50 | 8.2 | 0.79 | – | 15.3 | 0.07 | 50 | 28 |
| ВС, 50–110 | 9.1 | 0.20 | 0.1 | 12.1 | 0.05 | 43 | 24 |
| С, 110–160 | 9.1 | – | 0.1 | 11.4 | 0.07 | 46 | 27 |
| Солонец мелкий (Б-105-С) | |||||||
| А1, 0–8 | 7.3 | 1.82 | 0.8 | 1.0 | 0.03 | 38 | 18 |
| В1, 8–28 | 8.3 | 1.63 | 0.4 | 1.0 | 0.40 | 52 | 37 |
| В2, 28–37 | 8.3 | 0.63 | 3.6 | 13.6 | 0.29 | 52 | 30 |
| ВС, 37–95 | 8.4 | 0.44 | – | 13.6 | 0.86 | 44 | 26 |
| С, 95–118 | 8.6 | – | – | 8.3 | 0.61 | 42 | 23 |
| Сs, г, 118–160 | 8.3 | – | – | 9.8 | 1.81 | 42 | 26 |
Совершенно иная картина наблюдается в настоящее время (рис. 3.3 ). В современных почвах на территории первой надпойменной террасы не выявлено наличие комплексов. Серия прикопок с шагом 10 × 10 м выявила абсолютное доминирование светло-каштановых несолонцеватых солончаковатых почв с однотипными морфологическими характеристиками. Различия были связаны только с современными антропогенными нарушениями. Свойства такой почвы приведены в табл. 1 (разр. Б-75). Гор. А1 мощностью от 10 до 15 см комковато-порошистой структуры имеет волнистую нижнюю границу, часто видны потеки гумуса до глубины 20–25 см. Гор. В1 обеднен илом, имеет крупноглыбистую структуру, грани шероховатые, кутаны иллювиирования визуально не выражены. Педогенные карбонаты не обособляются от почвенной массы; легкорастворимые соли визуально не выражены, но при подсыхании стенок разреза уже с глубины 45–50 см становится заметна солевая корочка. С глубины 70–75 см аккумуляции легкорастворимых солей гипса. Причиной затухания солонцового процесса в почвах первой террасы, по всей видимости, стал подъем минерализованных грунтовых вод в связи с повышением уровня их разгрузки при заиливании реки.
Вторая надпойменная терраса. Ключевой участок Темрта-3. В почвенном покрове второй террасы и в XXVII–XXVI вв. до н. э., и в настоящее время выявляются ареалы светло-каштановых почв и солонцов. Но в отличие от почв первой надпойменной террасы, в погребенных почвенных комплексах на более высокой террасе доля солонцов была заметно меньше. При раскопках могильника Темрта-3 были исследованы погребенные почвы под тремя бровками на общей площади более 400 м2. Почвенный покров представлен комплексами светло-каштановых почв и солонцами, при этом было выявлено только три солонцовых пятна диаметром 5–7 м. Один из таких участков изображен на рис. 4.1. Таким образом, общая доля солонцов не превышала 20–30%. Опорный разрез погребенной почвы, в котором был вскрыт погребенный солонец и светло-каштановая несолонцеватая палеопочва, имел длину 6 м, на остальном участке погребенная почва была вскрыта до нижней границы гор. В2. Свойства почв представлены в табл. 1.
Рис. 4.
Ключевой участок Темрта-3, вторая надпойменная терраса. Профили погребенных XXVII–XXVI вв. до н. э. (1) и современных почв (2). Условные обозначения см. рис. 3. Высота поверхности от условного нуля.
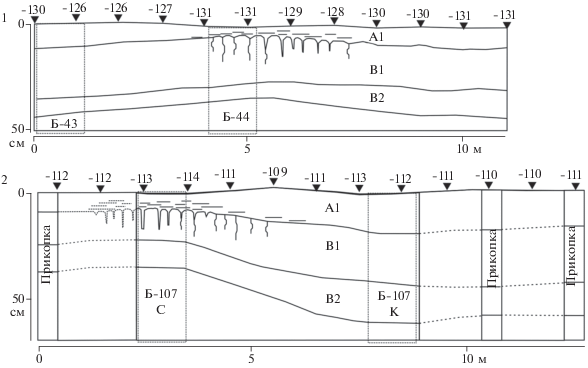
Как и в случае с первой надпойменной террасой, солонцовые пятна были приурочены к микропонижениям, но не во всех микропонижениях были зафиксированы солонцы. С другой стороны, в отличие от почв первой надпойменной террасы, где солонцовый процесс в той или иной мере проявлялся на всем протяжении вскрытой погребенной почвы, в данном случае в большей части погребенной светло-каштановой почвы признаки солонцового процесса морфологически не проявляются, и текстурная дифференциация слабо выражена: отношение содержания ила в гор. В1/А1 1.6. В солонцах этот показатель составляет 2.8, что также заметно меньше, по сравнению с погребенными солонцами на первой террасе.
Характерно, что, несмотря на хорошо выраженные признаки солонцового процесса, его наличие не вызвало заметной перестройки почвенного профиля в целом. Так, верхняя граница солевого горизонта, реконструируемая по появлению прожилок легкорастворимых солей и гипса, в солонце была на глубине 34 см, а в светло-каштановой почве на расстоянии 5–6 м – на глубине 44 см.
Исследование современных почв на второй террасе позволяет сделать вывод о схожести структуры современного и погребенного почвенного покрова. Например, в светло-каштановых почвах признаки солонцового процесса не проявлялись на макроуровне (рис. 4.2 ), а площадь солонцовых пятен, занятых чернополынно-прутняковыми ассоциациями, не превышала одной трети площади, доступной для исследования и не несущей следов современного антропогенного нарушения. Отметим, что, зачастую, именно там, где проходят следы от периодического проезда техники и прогона скота в почвах более выражены признаки солонцового процесса; но такие участки нами не принимались к рассмотрению. Однако в отличие от погребенных солонцовых комплексов общая организация профиля современных почв существенно варьировала при переходе от солонца к светло-каштановой почве. Так, легкорастворимые соли и гипс вымыты за пределы двухметровой толщи светло-каштановых почв, верхняя часть их профиля практически не несет признаков текстурной дифференциации, в то время как под солонцами верхняя граница солевой аккумуляции начинается уже с глубины 75 см, а отношение содержания ила в гор. В1/А1 2.8.
Водораздел. Ключевой участок Темрта-1. Погребенные почвы водораздела были исследованы при раскопках могильника Темрта-1. Было оставлено 5 бровок длиной до 30 м, что позволило получить полное представление о структуре погребенного почвенного покрова, с участием светло-каштановых почв и солонцов (рис. 5.1 ). Свойства обеих почвенных разностей близки, различия заключались лишь в морфологических признаках выраженности солонцового процесса, что проявилось в более выраженной листовато-чешуйчатой структуре гор. А1 и наличии визуально хорошо различимых натечных органо-глинистых кутан в гор. В1 в солонце, чего не было выявлено в светло-каштановой почве. Следует отметить, что в погребенных почвах солонцовый процесс был выражен в меньшей степени по сравнению с описанными ранее объектами. Так, под всеми бровками не выявлено ни одного участка с палисадной нижней границей гор. А1, и резким переходом между элювиальным и иллювиальным горизонтами. Довольно близка и невысока была степень текстурной дифференциации профиля – 1.9 в профиле солонца и 1.6 – в светло-каштановой почве. Видимо, почвы были погребены на начальном периоде развития солонцового процесса, так как профиль почв еще не претерпел существенных преобразований. Однако уже на этом этапе развития солонцовый процесс обусловил заметные различия в морфологических свойствах и стал причиной более близкого расположения солевого горизонта в профиле солонца. Что касается связи солонцов с элементами рельефа, то и в данном случае можно выявить на уровне тенденции приуроченность солонцов к микрозападинам.
Рис. 5.
Ключевой участок Темрта-1, локальный водораздел. Профили погребенных XXVII–XXVI вв. до н. э. (1) и современных почв (2). Условные обозначения см. рис. 3. Высота поверхности от условного нуля.

Отличия современных почвенных комплексов водоразделов от их погребенных аналогов выражаются в более глубоком развитии солонцового процесса, что проявляется в формировании в солонцовом горизонте вертикальных блоков-тумб с округлыми “головками” (рис. 5.2 ). Однако на современном этапе почти нет иллювиирования ила (табл. 1), а грани структурных отдельностей в гор. В1 шероховатые, кутаны морфологически не проявляются. Подобная ситуация была обнаружена нами на всех прикопках солонцового ареала, что позволяет говорить о затухании солонцового процесса на водораздельных участках в настоящее время. Причины этого явления нами пока не установлены.
Характеристика микробных сообществ подкурганных и современных почв солонцового комплекса
В подкурганных полеопочвах во всех исследованных объектах наибольшая суммарная численность микроорганизмов, исследованных трофических групп (средневзвешенные значения в гор. А1 + В1 + В2), выявлена в светло-каштановых палеопочвах (рис. 6, А). В палеосолонцах этот показатель был на 12–15% меньше. В каштаново-солонцовых комплексах на высокой террасе (ключевой участок Темрта-3) суммарная численность микроорганизмов была в 2 раза больше, чем в палеопочвах на низкой надпойменной террасе и водоразделе.
Рис. 6.
Суммарная численность микроорганизмов, выросших на ПА, НА и БС (средневзвешенные значения в гор. А1 + В1 + В2) в профиле подкурганных (А) и современных (Б) почв.

В современных светло-каштановых почвах суммарная численность микроорганизмов была на 14–28% больше, чем в солонцах (рис. 6, Б). При этом, как и в случае с погребенными почвами, в светло-каштановых почвах на высокой террасе величина этого параметра была в 1.4 и 3.5 раза выше, чем в аналогичных почвах на низкой террасе и водоразделе. Аналогичная закономерность выявлена и для солонцов.
Следовательно, как в подкурганных, так и в современных каштаново-солонцовых комплексах наибольшая суммарная численность микробных сообществ изученных трофических групп, выявлена в светло-каштановых почвах. Максимальное количество колониеобразующих единиц зафиксировано в светло-каштановых почвах и солонцах на высокой надпойменной террасе.
Численность микроорганизмов исследованных трофических групп в подкурганных комплексах изменялась по-разному (рис. 7, А). Численность микроорганизмов, выросших на почвенном агаре (ПА) и использующих элементы питания из рассеянного состояния, достоверно не различалась в светло-каштановой палеопочве и палеосолонце низкой (Темрта-5) и высокой надпойменных террас (Темрта-3). При этом на водораздельном участке (Темрта-1) численность микроорганизмов этой группы в палеосолонце была несколько больше, чем в светло-каштановой палеопочве.
Рис. 7.
Численность микроорганизмов (средневзвешенные значения в гор. А1 + В1 + В2) различных трофических групп в профиле подкурганных (А) и современных (Б) почв.

Численность микроорганизмов, использующих гумусовые вещества (НА), в палеосолонцах на низкой надпойменной террасе была в 1.3 раза больше по сравнению со светло-каштановыми палеопочвами, в то время как на водоразделе и высокой террасе была отмечена обратная картина.
Что касается численности микроорганизмов, выросших на богатой органической среде (БС) и использующих растительные остатки, то во всех случаях этот показатель был выше в погребенных светло-каштановых почвах по сравнению с погребенными солонцами (в 1.4, 1.3 и 1.5 раза, соответственно, в объектах Темрта-5, Темрта-3 и Темрта-1).
В современных почвенных комплексах на высокой террасе (Темрта-3) численность микроорганизмов, изученных трофических групп, была больше в светло-каштановой почве: в 1.5 раза на ПА, в 1.7 раза на НА и в 1.2 раза на БС (рис. 7, Б). В почве водораздела (Темрта-1) достоверные различия выявлены только по численности микроорганизмов, выросших на ПА, которые также были обильнее в светло-каштановой почве (в 1.2 раза) по сравнению с солонцом. Численность микроорганизмов, использующих гумусовые вещества и растительные остатки, в каштаново-солонцовом комплексе этого объекта достоверно не различалась.
Таким образом, в современных почвенных комплексах на высокой террасе выявлены более контрастные различия по численности микроорганизмов, выросших на ПА и НА, по сравнению с подкурганной почвой. Исключение составила численность микроорганизмов, использующих растительные остатки. Их различия были более контрастны в подкурганном комплексе. В почвах объекта Темрта-1, напротив, более контрастные различия по численности на НА и БС установлены в подкурганном комплексе. Изменение численности микроорганизмов, выросших на ПА, в комплексах современной и подкурганной почв имело противоположенную направленность.
Активная биомасса микробных сообществ, дающая отклик на внесение глюкозы, в подкурганных комплексах изменялась по-разному. В светло-каштановой палеопочве на высокой террасе она была в 1.8 раза больше, а в почвах на водоразделе – в 2.5 раза меньше чем в палеосолонце (табл. 2). Коэффициент дыхательной активности QR, характеризующий устойчивость микробных сообществ и представляющий собой отношение скоростей базального (Vбазал) и субстрат-индуцированного дыхания (VСИД), также различался. Если в почвах на высокой террасе он был больше в 1.9 раза в палеосолонце, то в почвах водораздела этот показатель был выше в светло-каштановой палеопочве, что свидетельствует о некотором нарушении устойчивости в микробном сообществе этих палеопочв.
Таблица 2.
Активность и устойчивость микробных сообществ подкурганных и современных почвенных комплексов (средневзвешенные значения в гор. А1 + В1 + В2 ± ошибка среднего)
| Объект | Разрез, почва | Биомасса микроорганизмов, мкг С/г почвы |
Доля активной биомассы в живой, % | Vбазал | VСИД | QR | |
|---|---|---|---|---|---|---|---|
| живая | активная | мкг С /г почвы/ч | |||||
| Погребенные почвы (XXVII–XXVI вв. до н. э.): | |||||||
| Темрта-3 | Б-43, К1 | – | 42.49 ± 7.17 | – | 0.05 ± 0.04 | 0.52± 0.09 | 0.09 |
| Б-44, Сн | – | 23.51 ± 4.45 | – | 0.05 ± 0.02 | 0.29 ± 0.06 | 0.17 | |
| Темрта-1 | Б-104, К1 | 432.03 ± 113.44 | 20.56 ± 5.07 | 4.76 | 0.09 ± 0.02 | 0.25 ± 0.06 | 0.36 |
| Б-104, Сн | 396.68 ± 89.40 | 50.91 ± 8.36 | 12.83 | 0.14 ± 0.02 | 0.62 ± 0.10 | 0.23 | |
| Современные почвы | |||||||
| Темрта-3 | Б-107-6, К1 | 527.42 ± 88.20 | 182.29 ± 3.73 | 34.56 | 1.24 ± 0.06 | 2.25 ± 0.05 | 0.55 |
| Б-107-0, Сн | 397.78 ± 30.70 | 138.82 ± 7.00 | 34.90 | 0.98 ± 0.08 | 1.71 ± 0.08 | 0.57 | |
| Темрта-1 | Б-105, К1 | 436.29 ± 97.95 | 155.45 ± 5.89 | 35.63 | 0.94 ± 0.05 | 1.91 ± 0.23 | 0.49 |
| Б-105, Сн | 427.43 ± 81.39 | 160.79 ± 4.04 | 37.62 | 1.13 ± 0.02 | 1.98 ± 0.05 | 0.57 | |
По величине живой биомассы, определяемой по содержанию фосфолипидов, которые являются обязательным компонентом мембран живых клеток независимо от их физиологического состояния, не установлено достоверных различий в подкурганном комплексе Темрта-1. Однако доля активной биомассы в живой была в 2.7 раза больше в палеосолонце по сравнению с подкурганной светло-каштановой почвой.
В современных каштаново-солонцовых комплексах и живая, и активная биомасса микроорганизмов были больше в светло-каштановой почве в 1.3 раза, чем в солонце на высокой террасе и не различались достоверно на водораздельном участке. Доли активной биомассы в живой в исследованных современных комплексах были сопоставимы. Коэффициент дыхательной активности QR незначительно увеличивался (в 1.2 раза) в солонце Темрта-1 и практически не различался в комплексе Темрта-3.
Таким образом, активность и устойчивость микробных сообществ подкурганных комплексов на разных элементах рельефа существенно варьировали, в то время как в современных почвенных комплексах различия более сглажены.
ЗАКЛЮЧЕНИЕ
Результаты исследования позволяют говорить о том, что в почвах Сальско-Манычской гряды в XXVII–XXVI вв. до н. э. солонцовый процесс в той или иной мере проявлялся на всех элементах рельефа. Наиболее ярко его признаки были выражены в почвах первой надпойменной террасы, где солонцы доминировали, а светло-каштановые почвы также несли признаки солонцового процесса. Но уже на более высокой террасе доля солонцов не превышала 25–30%, а в светло-каштановых почвах солонцовый процесс не проявлялся на макроуровне. Еще менее выраженные признаки солонцового процесса отмечены в погребенных почвах водораздельного участка.
В настоящее время на низкой надпойменной террасе солонцы не выявлены, в почвенном покрове доминируют светло-каштановые солончаковатые почвы с глубокими гумусовыми затеками. На более высокой террасе структура современного и погребенного почвенного покрова, в целом, однотипна. Обращает внимание только большая выраженность солонцового процесса в настоящее время и связанные с этим более глубокие изменения всех морфологических и химических свойств почв комплексов. При этом солонцовый процесс в настоящий момент находится в активной стадии, в то время как в почвах водораздельного участка отмечены признаки затухания солонцового процесса.
Для всех описанных выше погребенных и современных комплексов можно говорить об общей тенденции приуроченности солонцов к микропонижениям, но при этом наличие микропонижений не всегда приводит к развитию солонцов.
Несмотря на явно выраженные различия в особенностях проявления солонцового процесса в почвах на разных элементах рельефа в эпоху бронзы и в настоящее время, микробиологические свойства почв в погребенных и современных комплексах изменяются менее контрастно. Суммарная численность микроорганизмов (растущих на почвенном агаре и использующих элементы питания из рассеянного состояния, на нитритном агаре и потребляющие гумус, на богатой органической среде и разлагающие растительные остатки) в погребенных и современных почвах довольно близка в пределах каждого ключевого участка, но существенно различается на разных элементах рельефа. При этом и в светло-каштановых почвах, и в солонцах сохраняются закономерности в соотношении микроорганизмов разных трофических групп. То есть этот показатель не подвержен влиянию солонцеватости, а отражает климатические условия, имевшие место в момент сооружения кургана и в настоящее время.
Местные современные условия почвообразования на различных участках, к которым приурочены исследованные объекты, заметно отличаются: речные террасы и водораздел, различные уровни грунтовых вод, целина и залежь. Это нашло отражение в характеристике микробных сообществ изученных почв. Состояние микробных сообществ в тот или иной исторический период в значительной мере определяется закономерностями эволюции конкретного почвенного ареала (ландшафта) и вековой изменчивостью биоклиматических условий. В связи с этим возникает местная специфика в динамике состояния микробных сообществ, их структурно-функциональной организации и устойчивости.
Список литературы
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во Моск. ун-та, 1970. 488 с.
Благодатская Е.В., Ананьева Н.Д. Оценка устойчивости микробных сообществ в процессе разложения поллютантов в почве // Почвоведение. 1996. № 11. С. 1341–1346.
Болышев Н.Н. Происхождение и свойства почв полупустыни. М.: Изд-во Моск. ун-та, 1972. 196 с.
Большаков А.Ф., Боровский В.М. Почвы и микрорельеф Прикаспийской низменности // Солонцы Заволжья. Материалы изысканий, исследований и проектирования ирригации Заволжья. М.: ВАСХНИЛ, 1937. Вып. VII. С. 134–169.
Гедройц К.К. Избранные сочинения. Т. 1. Почвенные коллоиды и поглотительная способность почв. М.: Сельхозгиз, 1955. 559 с.
Геннадиев А.Н., Пузанова Т.А. Голоценовая эволюция почв и природно-экологических условий в низменной части Калмыкии // Проблемы древней истории Северного Прикаспия. Куйбышев, 1990. С. 71–72.
Гоголев И.Н., Волошин И.Н. Диагностические признаки и принципы классификации солонцеватых почв // Мелиорация солонцов. М.: Сельхозгиз, 1968. Ч. 1. С. 206–216.
Демкин В.А. Палеопочвы юго-востока Русской равнины в эпоху бронзы // Известия РАН. Сер. Геогр. 1995. № 2. С. 82–88.
Демкин В.А. Почвоведение и археология. Пущино, 1997. 213 с.
Демкин В.А., Борисов А.В., Демкина Т.С., Удальцов С.Н. Эволюция почв и динамика климата степей юго-востока Русской равнины в эпохи энеолита и бронзы (IV–II тыс. до н. э.) // Известия РАН. Сер. Геогр. 2012. № 1. С. 46–57.
Демкин В.А., Иванов И.В. Развитие почв Прикаспийской низменности в голоцене. Пущино, 1985. 165 с.
Демкин В.А., Лукашов А.В. О скорости и направленности почвообразовательного процесса в зоне сухих степей в голоцене // Почвоведение. 1987. № 6. С. 5–14.
Демкина Т.С., Попова И.В., Демкин В.А. Характеристика микробных сообществ современных и подкурганных почв солонцовых комплексов сухих степей Нижнего Поволжья // Почвоведение. 2013. № 7. С. 840–849. https://doi.org/10.7868/S0032180X13070034
Звягинцев Д.Г., Асеева И.В., Бабьева И.П., Мирчинк Т.Г. Методы почвенной микробиологии и биохимии. М.: Изд-во Моск. ун-та, 1980. 224 с.
Иванов И.В. Эволюция почв степной зоны в голоцене. М.: Наука, 1992. 140 с.
Иванова Е.Н. Солонцы каштановой зоны междуречья Волга-Урал // Почвы комплексной равнины Северного Прикаспия и их мелиоративная характеристика. М.: Наука, 1964. С. 114–155.
Иванова Е.Н., Фридланд В.М. Почвенные комплексы сухих степей и их эволюция // Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР. М.: Изд-во АН СССР, 1954. С. 162–190.
Кауричев И.С. Почвоведение. М.: Колос, 1975. 496 с.
Князева Н.В. Пептизация почв и активность ионов Na в почвенных суспензиях // Почвоведение. 1981. № 8. С. 52–60.
Ковда В.А. Почвы Прикаспийской низменности. М.: Изд-во АН СССР, 1950. 256 с.
Конюшкова М.В., Абатуров Б.Д. Особенности микрорельефа и свойства почв солонцового комплекса на поздних стадиях развития в Прикаспийской низменности // Бюл. Почв. ин-та им. В.В. Докучаева. 2016. Вып. 83. С. 53–79.
Любимова И.Н., Хан В.В., Салпагарова И.А. Диагностика солонцового процесса в целинных и агрогенно-измененных почвах разных регионов // Почвоведение. 2014. № 9. С. 1046–1055.
Можарова Н.В., Федоров К.Н. Эволюция почвенных мезоструктур аккумулятивно-морской равнины Терско-Кумской низменности // Биологические науки. 1990. № 2. С. 132–144.
Новикова А.В., Коваливнич П.Г. О корреляционной зависимости между степенью солонцеватости и степенью иллювиированности в солонцовых почвах Крымского Присивашья // Материалы Всес. научн.-техн. совещ. по пробл. мелиорации солонцов. М.: 1968. С. 216–227.
Роде А.А., Польский М.Н. Почвы Джаныбекского стационара // Тр. Почв. ин-та им. В.В. Докучаева. 1961. Т. 56. С. 3–214.
Теории и методы физики почв / Под ред. Шеина Е.В., Карпачевского Л.О. М.: Гриф и К, 2007. 616 с.
Теппер Е.З. Микроорганизмы рода Nocardia и разложение гумуса. М.: Наука, 1976. 199 с.
Фридланд В.М. Структура почвенного покрова. М.: Мысль, 1972. 424 с.
Хитров Н.Б. Генезис, диагностика, свойства и функционирование глинистых набухающих почв Центрального Предкавказья. М.: Типография Россельхозакадемии, 2003. 505 с.
Хитров Н.Б. Выбор диагностических критериев существования и степени выраженности солонцового процесса в почвах // Почвоведение. 2004. № 1. С. 18–31.
Хитров Н.Б. Связь почв солонцового комплекса Северного Прикаспия с микрорельефом // Почвоведение. 2005. № 3. С. 271–284.
Хомутова Т.Э., Демкина Т.С., Борисов А.В., Шишлина Н.И. Состояние микробных сообществ подкурганных палеопочв пустынно-степной зоны эпохи средней бронзы (XXVII–XXVI вв. до н. э.) в связи с динамикой увлажненности климата // Почвоведение. 2017. № 2. С. 239–248. https://doi.org/10.7868/S0032180X1702006X
Хомутова Т.Э., Каширская Н.Н., Демкин В.А. Оценка живой и суммарной биомассы микробных сообществ современной каштановой почвы и подкурганных палеопочв // Почвоведение. 2011. № 12. С. 1496–1503.
Чижикова Н.П., Ковда И.В., Борисов А.В., Шишлина Н.И. Проявление солонцового процесса в современных и подкурганных почвах и его отражение в минералогической памяти почв // Почвоведение. 2009. № 10. С. 1266–1277.
Anderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. Biochem. 1978. V. 10. № 3. P. 215–221.
Дополнительные материалы отсутствуют.