

Почвоведение, 2019, № 5, стр. 633-642
Изменение лесных экосистем мегаполиса под влиянием рекреационного воздействия
В. А. Кузнецов 1, *, И. М. Рыжова 1, Г. В. Стома 1
1 МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: xts089@gmail.com
Поступила в редакцию 30.06.2018
После доработки 21.09.2018
Принята к публикации 28.11.2018
Аннотация
Представлена комплексная количественная оценка показателей состояния растительности, сообщества мезопедобионтов, характеристик подстилки и свойств дерново-подзолистых почв (Albic Retisols) лесопарков Москвы (“Битцевский лес” и “Лосиный остров”) в зависимости от уровня рекреационной нагрузки. Показано, что изменения основных компонентов лесных экосистем в изучаемых лесопарках имеют одинаковую направленность. К пятой стадии дигрессии в 4–5 раз уменьшается доля деревьев в хорошем состоянии, сомкнутость крон снижается до 20–30%, что приводит к увеличению освещенности более чем в 30 раз, обилие подлеска снижается в 3–4 раза. В травяном ярусе видовое разнообразие с увеличением нагрузки сначала возрастает с 9–11 видов до 23 за счет внедрения лесолуговых, луговых и сорных видов, а затем снижается. Уменьшаются запасы подстилки. В ее фракционном составе доля активной фракции снижается в 1.7–2 раза, а измельченной возрастает в 3.5–7 раз. Плотность и биомасса мезопедобионтов к пятой стадии дигрессии уменьшаются в 2–2.5 раза, а в подстилке в 4–7 раз. Наибольшие изменения почвенных свойств отмечены в верхнем минеральном слое (0–5 см) и уменьшаются с глубиной. По результатам дискриминантного анализа, наибольший вклад в дискриминацию почв по стадиям дигрессии вносит коэффициент структурности. Одиночные вклады других почвенных свойств малы, поэтому при дискриминации важен только их совокупный вклад.
ВВЕДЕНИЕ
В настоящее время урбанизация стала явлением глобального масштаба. С ростом численности городского населения увеличивается значение рекреационной функции лесных экосистем, оказавшихся в городской черте и пригородных зонах, в формировании благоприятной среды обитания. В то же время рекреация относится к числу негативных антропогенных факторов, влияющих не только на отдельные компоненты леса, но и оказывающих действие на экосистемном уровне.
Рекреационная экология активно развивается, начиная с 1960-х гг. К настоящему времени по этой теме опубликованы сотни работ [3, 8, 10, 12, 17–20, 25, 26, 28–34]. Большинство из них посвящены изучению рекреационных лесов [12, 16–20, 25, 26]. Несмотря на многочисленность исследований, многие важные вопросы остаются дискуссионными.
Для характеристики этапов изменения лесных биогеоценозов при возрастании рекреационной нагрузки, начиная с работы Карпиносовой [14], широко и успешно используется подход, основанный на выделении стадий дигрессии (СД). Однако до настоящего времени нет однозначного ответа на вопрос о количестве стадий и наборе показателей состояния лесных экосистем для характеристики рекреационных изменений. Наряду с наиболее распространенной пяти стадийной схемой предлагаются как более, так и менее детальные схемы [3, 12, 23, 25, 26, 28, 32]. Спорным остается вопрос о том, какой вид имеют функции отклика показателей состояния экосистемы от уровня воздействия [33]. Не выработана единая точка зрения по вопросу о критической рекреационной нагрузке. Трудности поиска решений этих вопросов объясняются сложностью проблемы. Устойчивость экосистем к рекреационному воздействию может быть разной в зависимости от характера растительности, типа почв, их гранулометрического состава и др. Кроме того, основные компоненты леса и их характеристики могут различаться по чувствительности к рекреационной нагрузке. Для прогнозирования динамики состояния рекреационных лесов и выбора оптимального управления необходима дополнительная комплексная количественная информации о состоянии экосистем при разных уровнях рекреационного воздействия, позволяющая использовать современные математические методы анализа данных.
Целью настоящей работы является комплексная количественная оценка изменения показателей состояния растительности, мезофауны подстилки, физических, химических и биологических свойств почв от уровня рекреационного воздействия на примере лесопарков Москвы.
ОБЪЕКТЫ И МЕТОДЫ
В качестве объектов исследований были выбраны лесные экосистемы двух крупнейших особо охраняемых природных территорий Москвы: национального парка “Лосиный остров” и природно-исторического парка “Битцевский лес”. Работа выполнялась на территории, непосредственно примыкающей к жилым кварталам и активно используемой для рекреации. В “Лосином острове” это участки 52–54 кварталов, в “Битцевском лесу” исследования проводились в лесном массиве усадьбы Узкое (квартал 7).
Полевые исследования проводились в середине лета в 2011–2013 гг. В каждом лесопарке было заложено по пять пробных площадей размером 25 × 25 м, соответствующих пяти уровням рекреационной нагрузки, определенной по доле вытоптанной площади, характеризующей разные стадий рекреационной дигрессии [23]. Все пробные площади находились в сходных геоморфологических условиях на автономных элементах рельефа.
В “Лосином острове” исследования проводились в елово-липовом лесу на мелких дерново-подзолистых (Albic Retisols (Loamic, Ochric) по WRB, 2014) глубокоосветленных легкосуглинистых почвах на покровных суглинках, подстилаемых флювиогляциальными отложениями. На тропинках представлены их крайне мелкие поверхностно оглеенные варианты. В “Битцевском лесу” под дубово-липовым лесом представлены средне и мелко дерново-подзолистые (Albic Retisols (Loamic, Ochric) по WRB, 2014) неглубокоосветленные легко- и среднесуглинистые почвы на покровных суглинках, подстилаемых мореной. На тропинках преобладали их мелкие и средне мелкие, мелко- и неглубоко осветленные поверхностно оглеенные разности [15]. По WRB (2014) изучаемые почвы относятся к Albic Retisols. На каждой пробной площади сделано геоботаническое описание всех ярусов леса по традиционным методикам. Состояние деревьев оценивали по комплексу показателей, предложенных Мозолевской с соавт. [21]. Дополнительно определялся показатель стабильности развития липы мелколистной (Tilia cordata) по методике Захарова и Чубинишвили [11]. При этом для характеристики условий среды на каждой из стадий дигрессии промерялось по 500 листовых пластинок. Для характеристики травяного яруса использованы эколого-ценотические группы, предложенные Ниценко [22], классифицирующие все разнообразие травяных видов в 4 группы по отношению к совокупности экологических факторов: 1 – лесные, 2 – лесо-луговые, 3 – луговые, 4 – сорно-рудеральные. Эти группы выделяются в большинстве работ, посвященных рекреационным лесам [3, 12, 25, 26].
Для оценки сходства видового состава травяных сообществ на разных стадиях дигрессии вычислялись широко используемые в биогеоценологии индексы общности Жаккара и Съеренсена–Чекановского [5].
Освещенность леса определялась фотоэлектрическим люксметром Ю-117 на уровне поверхности почвы и на высоте 1.5 м в 36-кратной повторности на каждой пробной площади.
В пределах пробных площадей, характеризующих каждую из стадий дигрессии, рекреационная нагрузка на почвы распределяется неравномерно. Максимальная нагрузка приходится на почвы тропинок. Она ослабевает по мере удаления от тропинок по направлению к наименее нарушенному ядру пробной площади. Поэтому образцы подстилки и почв отбирались на тропинках, в зонах вдоль тропинок на удалении 20, 50 и 100 см и вне зоны прямого влияния тропинок. Чтобы учесть высокую пространственную вариабельность свойств почв в лесу, обусловленную влиянием деревьев-эдификаторов, создающих мощные фитогенные поля [13], на участках вне зоны прямого влияния тропинок образцы отбирались методом заложения трансект. На каждой пробной площади было заложено по три трансекты, на которых образцы отбирались по прямой линии от ствола одного дерева до ствола другого: у ствола, в середине проекции кроны и в межкроновом пространстве (окне).
Подстилка отбиралась рамкой 25 × 25 см. Определялись ее тип (по классификации Богатырева [2], мощность, запасы и фракционный состав (доля измельченной (<1 cм), активной (листья) и пассивной (ветки, кора, хвоя) фракций), влажность и кислотность (подстилка : вода 1 : 25).
Глубина отбора почвенных образцов составляла 0–5, 5–10 и 10–20 см, так как влияние рекреации ослабляется с глубиной [16–20, 25, 26].
Определение почвенных свойств проводилось по общепринятым в почвоведении методикам: актуальная кислотность (рНвод) – в водной почвенной суспензии при соотношении почва : вода 1 : 2.5, содержание органического углерода – титриметрическим вариантом метода В.И. Тюрина, твердость – с помощью твердомера Н.А. Качинского, плотность – буровым методом путем отбора определенного объема почвы естественного сложения, агрегатный состав – с помощью ситового анализа: в сухом состоянии (сухое просеивание) и в стоячей воде (мокрое просеивание) [4, 27]. Электропроводность почв определялась в суспензии с соотношением почва : вода 1 : 1 на приборе Cond-315i [35]. Для характеристики биологической активности почв определялось субстрат-индуцированное и базальное дыхание [1].
Учет численности, биомассы и группового состава почвенных беспозвоночных проводился в трехкратной повторности методом ручной разборки монолитов площадью 1/16 м2, вырезанных послойно: подстилка, 0–10, 10–20 см [6]. Классификация почвенных беспозвоночных на отдельные систематические группы и классы проводилась согласно представлениям, изложенным в работе Догеля [9]. В связи с задачами исследования мы ограничились рассмотрением лишь классов мезопедобионтов.
Разнообразие и степень доминирования почвенной мезофауны оценивалось индексами неоднородности Шеннона–Уивера и Симпсона [5].
Статистический анализ результатов проводился в пакете Statistica. Выбранный уровень значимости 0.05. При изучении зависимости изменений почвенных свойств от уровня рекреационной нагрузки анализировались послойные выборки, при составлении которых учитывались доли площади тропинок, зон вдоль тропинок и территории вне прямого влияния тропинок от общей площади пробного участка на каждой из стадий дигрессии.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Полученные данные позволяют проследить за изменениями характеристик растительного покрова, подстилки, почв и показателей состояния сообщества мезопедобионтов в елово-липовых и дубово-липовых экосистемах на дерново-подзолистых почвах (Albic Retisols по WRB, 2014) в зависимости от уровня рекреационного воздействия.
Состояние растительности. Подробная количественная оценка характеристик растительного покрова и подстилки на разных стадиях рекреационной дигрессии рассмотрена нами ранее [17], поэтому здесь приведем только основные результаты. Рекреация оказывает влияние на все ярусы растительного покрова.
Древесный ярус. С ростом рекреационной нагрузки в 4–5 раз уменьшается доля деревьев в хорошем состоянии (табл. 1). Чувствительность к рекреационному воздействию зависит от видового состава древостоя. В хвойно-широколиственном лесу доля деревьев в хорошем состоянии сокращается вдвое при переходе к третьей стадии дигрессии, а в широколиственном лесу только к четвертой. Уже на второй стадии дигрессии в “Лосином острове” доля деревьев в неудовлетворительном состоянии в два раза больше, чем в “Битцевским лесу”, что связано с меньшей устойчивостью к рекреационной нагрузке хвойных пород по сравнению с лиственными [3, 17, 18, 26]. Под влиянием рекреации происходит оголение и повреждение корней деревьев. Хвойные породы, имея поверхностную корневую систему, страдают сильнее.
Таблица 1.
Показатели состояния древостоя на разных стадиях рекреационной дигрессии (над чертой – количество деревьев/100 м2, под чертой – доля, %)
| Категория состояния деревьев (по [21]) | “Лосиный остров” | “Битцевский лес” | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| стадия дигрессии | ||||||||||
| I | II | III | IV | V | I | II | III | IV | V | |
| сомкнутость крон, % | ||||||||||
| 70–75 | 65–70 | 30–35 | 25–30 | 20–25 | 75–80 | 60–65 | 50–55 | 40–45 | 30–35 | |
| 1. Хорошее | 14 | 14/56 | 11/44 | 8/38 | 4/18 | 24/88 | 24/78 | 19/66 | 11/42 | 6/24 |
| 2. Удовлетворительное | 2/7 | 6/24 | 7/28 | 7/29 | 8/36 | 3/12 | 3/11 | 5/17 | 9/35 | 8/32 |
| 3. Неудовлетворительное: | 0/0 | 5/20 | 7/28 | 8/33 | 10/46 | 0/0 | 3/11 | 5/17 | 6/24 | 11/44 |
| 3.1. Количество усыхающих деревьев | 0/0 | 4/16 | 4/16 | 5/15 | 5/24 | 0/0 | 2/7 | 3/10 | 4/15 | 9/36 |
| 3.2. Количество сухих деревьев | 0/0 | 1/4 | 3/12 | 3/15 | 5/24 | 0/0 | 1/4 | 2/7 | 2/8 | 2/8 |
| 4. Количество деревьев с оголенными корнями | 0/0 | 0/0 | 1/4 | 3/13 | 11/50 | 0/0 | 0/0 | 0/0 | 1/4 | 4/16 |
| Показатель стабильности развития липы мелколистной (по [11]), среднее значение n=500 | 0.030 | 0.035 | 0.039 | 0.042 | 0.049 | 0.028 | 0.031 | 0.034 | 0.041 | 0.045 |
В результате ухудшения состояния древостоя уменьшается сомкнутость крон деревьев (с 70–80 до 20–30%). Это приводит к увеличению освещенности, величина которой на пятой стадии дигрессии возрастает в изучаемых лесопарках более чем 30 раз. Ее изменение сказывается на количестве подроста и видовом разнообразии трав.
Об ухудшении условий среды при возрастании рекреационной нагрузки свидетельствует увеличение показателя стабильности развития (ПСР) липы мелколистной, основанного на определении флуктуирующей асимметрии листовой пластинки. На высокую чувствительность этого показателя указывают результаты дисперсионного анализа, свидетельствующие о статистической значимости различия средних значений ПСР на всех стадиях дигрессии.
Состояние подлеска служит важным показателем нарушенности леса под влиянием рекреации [3, 7, 12, 26]. Обилие подлеска мало изменяется на первых четырех стадиях дигрессии, резко уменьшаясь в 3–4 раза на пятой стадии. Изменения в видовом разнообразии проявляются при более слабой рекреационной нагрузке. Уже на второй стадии неустойчивые виды исчезают, в то время как компактные кусты лещины обыкновенной, способные выдерживать большую нагрузку, разрастаются. На третьей и четвертой СД происходит внедрение опушечных видов. На пятой стадии остаются лишь единичные особи лещины обыкновенной.
Количество подроста на первой и второй стадиях дигрессии составляет примерно 10 экз./100 м2. С ростом освещенности на третьей и четвертой стадиях оно увеличивается почти вдвое. При максимальной рекреационной нагрузке на пятой стадии в результате механического повреждения и уплотнения почв количество подроста уменьшается до 2–6 экз./100 м2.
Всходы приурочены к освещенным участкам, лишенным растительного покрова и подстилки [7, 26]. Третья и четвертая стадия дигрессии характеризуются оптимальными условиями для появления и развития всходов, что приводит к увеличению их видового разнообразия и обилия (в 2–4 раза относительно первой стадии). С дальнейшим увеличением рекреационной нагрузки обилие и видовое разнообразие всходов уменьшается: они механически повреждаются, часть из них гибнет, не успевая перейти в категорию подроста. На пятой стадии дигрессии в начале вегетационного сезона (в мае) количество всходов составляет 30–40 экз./100 м2, а к концу июля они полностью исчезают.
В травяном ярусе при слабой рекреационной нагрузке выделено 9–11 видов растений, относящихся в основном к лесной эколого-ценотической группе. При среднем ее уровне вследствие увеличения разнообразия условий местообитания возрастает видовое разнообразие трав за счет внедрения лесолуговых, луговых и сорных видов. Максимальное количество видов трав (23) отмечается при освещенности в 25%: в “Лосином острове” на третьей, а в “Битцевском лесу” на четвертой стадии дигрессии. Дальнейшее увеличение нагрузки приводит к уменьшению видового разнообразия. В травяном ярусе начинают доминировать светолюбивые, устойчивые к вытаптыванию и механическим повреждениям виды. Показатели сходства видового состава травяных сообществ на разных стадиях дигрессии (коэффициент флористического сходства Жаккара и коэффициент общности видового состава Съеренсена–Чекановского) свидетельствуют о резком повышении различий в видовом составе при переходе к СД, характеризующейся максимальным видовым разнообразием. В елово-широколиственном лесу, по сравнению с широколиственным, видовой состав травяного яруса на разных стадиях дигрессии различается сильнее.
Моховой ярус на изучаемой территории встречаются фрагментарно. К третьей СД его площадь увеличивается c 3–5 до 7–10%, а затем в результате усиления рекреационного воздействия на четвертой и пятой стадиях мхи исчезают.
Характеристики подстилки. Подстилка является одним из наиболее уязвимых компонентов рекреационных лесов. По мере усиления рекреационной нагрузки изменяется количество и соотношение подгоризонтов подстилки. В соответствии с классификацией подстилок Богатырева [2] на первой–второй СД они относятся к типу гумифицированных, их мощность составляет в среднем 2–4 см. На третьей стадии отмечается фрагментарность подстилки. Тип подстилок меняется на деструктивный на четвертой стадии. В этом случае они представлены только подгоризонтом листового опада мощностью <1 см. На пятой стадии дигрессии подстилка вдавливается и перемешивается с верхним минеральным слоем почвы.
Запасы подстилки в лесных экосистемах характеризуются высокой пространственной вариабельностью [24]. Под влиянием рекреации на второй и третьей стадиях изменчивость еще больше увеличивается: дисперсия запасов подстилки на этих стадиях максимальна и в 5–7 раз больше, чем на первой. Статистически значимое уменьшение запасов подстилки отмечено лишь на пятой СД (рис. 1).
Рис. 1.
Зависимость изменений свойств подстилки от уровня рекреационного воздействия в лесопарках “Лосиный остров” (А) и “Битцевский лес” (Б). Фракции подстилки: 1 – активная (листья); 2 – пассивная (ветки, кора, хвоя); 3 ‒ измельченная (<1 см). Доверительный интервал 95%.
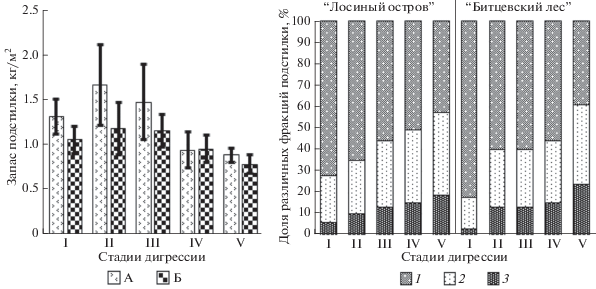
С возрастанием рекреационной нагрузки изменяется фракционный состав подстилки. На первой–четвертой СД в составе подстилки преобладает активная фракция. При вытаптывании в результате измельчения и перетирания подстилки значительно возрастает доля измельченной фракции. В елово-широколиственном лесу “Лосиного острова” она увеличивается в 3 раза, а в широколиственном “Битцевском лесу” – в 8 раз. Доля пассивной фракции (веток, коры и хвои деревьев) также увеличивается в 2–2.5 раза. Это, по всей видимости, связано с ухудшением состояния деревьев и их механическим повреждением. В “Лосином острове” достоверное изменение состава подстилки происходит при переходе к третьей, а в “Битцевском лесу” – уже на второй СД.
В результате увеличения поступления атмотехногенной пыли из-за уменьшения сомкнутости крон деревьев уменьшается кислотность подстилки. В хвойно-широколиственном лесу “Лосиного острова” значения рН увеличивается с 5.5 до 5.9, а в широколиственном “Битцевском лесу” – с 5.8 до 6.0.
Изменение состояния комплекса почвенных беспозвоночных. Мезопедобионты при рекреации подвергается как прямому воздействию, которое выражается в их механическом уничтожении, так и опосредованному – в результате изменения условий среды обитания. На фоновых участках комплекс почвенных беспозвоночных в изучаемых парках характеризуется близкими значениями основных показателей. Плотность и биомасса составляют в среднем соответственно 430–490 экз./м2 и 150–170 г/м2. К пятой СД эти показатели уменьшаются в 2–2.5 раза. Особенно сильное угнетение испытывает мезофауна подстилки, где численность и биомасса уменьшаются в 2 раза, уже на третьей стадии, а на пятой – в 4–7 раз. По мере усиления рекреационной нагрузки уменьшается разнообразие почвенных беспозвоночных. В их составе увеличивается доля малощетинковых червей – с 17–22 до 53–77%, и в 2–3 раза сокращается доля многоножек и насекомых [19].
Изменение свойств почв. Предыдущие исследования свидетельствует о значительных изменениях физических, химических и биологических свойств почв при высоком уровне рекреационной нагрузки, которую испытывают почвы тропинок [16, 20]. Представляется интересным количественно оценить изменения свойств почв на изучаемой территории лесопарков в целом в зависимости от уровня рекреационного воздействия.
С этой целью сравним свойства почв на разных СД путем анализа послойных выборок, что позволяет учесть неравномерность рекреационной нагрузки в пределах участков, характеризующих каждую из стадий. Наиболее заметные изменения свойств почв наблюдаются в верхнем минеральном (0–5 см) слое. С глубиной влияние рекреации на свойства почв ослабляется, поэтому на рисунках, характеризующих зависимость изменения свойств почв от уровня рекреационной нагрузки, представлены данные для слоя 0–5 см. Зависимость физических свойств почв от уровня рекреационной нагрузки характеризует рис. 2.
Рис. 2.
Зависимость изменения физических (А), химических и физико-химических (Б) и биологических (В) свойств почв от уровня рекреационного воздействия. Здесь и далее: 1 – среднее; 2 – ошибка среднего; 3 – доверительный интервал среднего (95%). I–V – стадии дигрессии.
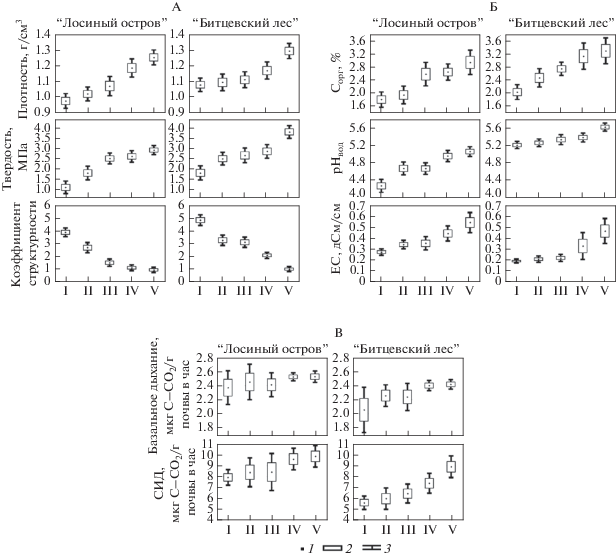
Плотность почв. При рекреации плотность почв возрастает. Статистически значимое увеличение плотности почв елово-липового леса “Лосиного острова” отмечено на четвертой СД, а в дубово-липовом “Битцевском лесу” – на пятой СД и прослеживается до глубины 20 см, однако амплитуда этих изменений с глубиной уменьшается в 2 раза.
В дубово-липовом лесу “Битцевского лесопарка” даже на пятой СД плотность почв до глубины 20 см остается оптимальной для дуба и липы, но становится критической для лесных трав. В елово-липовом лесу “Лосиного острова” плотность почв уже на четвертой стадии превышает верхнюю границу допустимых условий для произрастания ели.
Твердость почв является чувствительным индикатором на рекреационную нагрузку, ее статистически значимое увеличение отмечено уже на второй СД. К пятой СД твердость почв в “Битцевском лесу” и “Лосином острове” возрастает, соответственно, до 3.8 и 2.9 МПа.
Структура почвы. Показатели, характеризующие структуру почвы, как и твердость, высокочувствительны к рекреационной нагрузке. Они статистически значимо изменяются при переходе уже ко второй СД. На первой стадии дигрессии почвы характеризуются как хорошо оструктуренные (коэффициент структурности >1.5, а сумма агрономически ценных агрегатов >60%). К пятой СД коэффициент структурности уменьшается в 4–5 раз, сумма агрономически ценных агрегатов – в 1.8–2 раза. Полученные данные согласуются с литературными [13, 32].
По мере усиления рекреационного воздействия уменьшается водоустойчивость агрегатов (табл. 2).
Таблица 2.
Суммарное количество агрегатов диаметром более 0.25 мм при мокром просеивании (%)
| Лесопарк | Стадия дигрессии | ||||
|---|---|---|---|---|---|
| I | II | III | IV | V | |
| “Лосиный остров” | 76.7 | 74.6 | 74.9 | 68.9 | 55.2 |
| “Битцевский лес” | 79.0 | 67.6 | 61.1 | 51.2 | 33.5 |
Содержание органического углерода в почве по литературным данным под действием рекреации может, как увеличиваться, так и уменьшаться [3, 16–20, 25, 26, 28, 30, 34]. В почвах изучаемых лесопарков отмечено его увеличение (рис. 3 ). Статистически значимые изменения содержания органического углерода отмечены на третьей СД. На пятой стадии содержание органического углерода в почве достигает 2.9–3.3%, а также в 1.5–2 раза увеличивается его коэффициент вариации. В более глубоких слоях почвы увеличения содержания органического углерода под влиянием рекреации не прослеживается. Увеличение содержания органического вещества в почве при рекреации может быть обусловлено общим городским атмотехногенным загрязнением углеродсодержащими соединениями (поступлением сажи), дополнительным локальным загрязнением (остатки кострищ, корм для животных, различный мусор и отходы) и вдавливанием фрагментов опада и подстилки [3, 19, 22–25, 28].
Реакция среды. С увеличением рекреационной нагрузки кислотность почв изучаемых лесопарков уменьшается. Дерново-подзолистые почвы (Albic Retisols по WRB, 2014) хвойно-широколиственного леса “Лосиного острова” более кислые и статистически значимое увеличение рНвод в них происходит уже на второй СД, а к пятой стадии рН увеличивается на 0.8 единиц. В почве широколиственного “Битцевского леса” статистически значимое увеличение рН отмечено только к пятой СД. Зависимость кислотности почв от рекреационного воздействия прослеживается до глубины 20 см. Однако диапазон изменений значений рНвод сужается. Уменьшение кислотности изучаемых почв связано с поступлением атмотехногенной пыли, антигололедных реагентов и загрязнением почв азотсодержащими экскрементами домашних животных и продуктами их распада [28–30].
Электропроводность почв зависит от количества поступающих загрязняющих веществ с атмосферными осадками, пылью и противогололедными реагентами, привносимыми на обуви посетителей, ветром и при снегоуборочных работах. По мере усиления рекреационной нагрузки отмечено статистически достоверное увеличение электропроводности почв “Лосиного острова” при переходе к четвертой, а “Битцевского леса” – лишь к пятой СД. На пятой СД она увеличивается в 2–2.5 раза, а ее коэффициент вариации – в 3–5 раз. Зависимость электропроводности почв от уровня рекреационной нагрузки ослабевает с глубиной. Статистически значимые изменения отмечаются: в “Лосином острове” до глубины 10 см и в “Битцевском лесу” – до 20 см.
Биологическая активность почв оценивалась по данным о базальном и субстрат-индуцированном дыхании. Базальное дыхание почв характеризуются высокой пространственной изменчивостью. По-видимому, этим объясняется отсутствие статистически значимых различий этого показателя на участках, характеризующих разные СД (рис. 4 ).
Увеличение базального дыхания почв с переходом от первой к пятой СД проявляется только как тенденция. Для субстрат-индуцированного дыхания статистически значимые различия выявлены в “Лосином острове” при переходе к пятой, а в “Битцевском лесу” – к четвертой СД.
Полученные результаты согласуются с литературными данными. Увеличение биологической активности почв объясняется тем, что при не очень высокой антропогенной нагрузке микробное сообщество сопротивляется отрицательному влиянию, что выражается в увеличении интенсивности дыхания. Возрастание деструкции органического вещества при рекреации обусловлено изменением его качества, так как в результате вытаптывания происходит измельчение растительных остатков. При умеренном рекреационном воздействии биологическая активность может возрастать [10, 25, 26, 29–32].
Результаты дискриминантного анализа. Для того чтобы выяснить, какие из характеристик подстилок и почвенных свойств вносят наибольший вклад в дискриминацию СД, мы провели дискриминантный анализ полученных данных. Учитывались характеристики подстилок и свойства верхнего минерального слоя почв мощностью 5 см. Объем выборки для каждого изучаемого лесопарка составлял 140.
Результаты пошагового дискриминантного анализа (табл. 3) свидетельствуют о статистической значимости дискриминации. Но она не очень высока, так как критерий Уилкса, характеризующий дискриминационную способность моделей и изменяющийся от 0 (полная дискриминация) до 1 (нет никакой дискриминации), составляет для моделей, характеризующих почвы “Лосиного острова” и “Битцевского леса”, соответственно 0.20 и 0.17.
Таблица 3.
Результаты пошагового дискриминантного анализа данных
| Переменные в модели | Критерий Уилкса | Частный критерий Уилкса | Р – уровень значимости | Толерантность |
|---|---|---|---|---|
| “Лосиный остров” (критерий Уилкса 0.20; переменные вне модели: электропроводность почв (EС)) |
||||
| Коэффициент структурности | 0.356 | 0.560 | 0.000000 | 0.633 |
| рНвод подстилки | 0.233 | 0.857 | 0.000314 | 0.415 |
| Запасы подстилки | 0.217 | 0.918 | 0.019859 | 0.739 |
| pHвод почвы | 0.227 | 0.879 | 0.001455 | 0.389 |
| Твердость почвы | 0.222 | 0.898 | 0.005476 | 0.321 |
| Плотность почвы | 0.218 | 0.916 | 0.018015 | 0.367 |
| Содержание Сорг | 0.211 | 0.9460 | 0.109535 | 0.573 |
| “Битцевский лес” (критерий Уилкса 0.17; переменные вне модели: рНвод подстилки; рНвод почвы) |
||||
| Коэффициент структурности | 0.469 | 0.367 | 0.000000 | 0.521 |
| Твердость почвы | 0.203 | 0.849 | 0.000447 | 0.267 |
| Плотность почвы | 0.192 | 0.894 | 0.007971 | 0.472 |
| Запасы подстилки | 0.185 | 0.929 | 0.060275 | 0.660 |
| Содержание Сорг | 0.205 | 0.837 | 0.000199 | 0.298 |
| EС почвы | 0.198 | 0.870 | 0.001771 | 0.316 |
Одиночный вклад каждой переменной в дискриминацию характеризует частный критерий Уилкса: чем меньше его значение, тем большую роль играет соответствующая переменная в дискриминации. Полученные результаты показали, что коэффициент структурности почв является ведущей переменной. Его вклад в дискриминацию проявляется сильнее в случае среднесуглинистых почв “Битцевского леса” (значение частной лямбды Уилкса составляет 0.367) по сравнению с легкосуглинистыми почвами “Лосиного острова” (частная лямбда Уилкса равна 0.560). Одиночные вклады остальных переменных малы (значения частной лямбды Уилкса для них >0.8). Для дискриминации почв по стадиям рекреационной дигрессии имеет значение их совокупный вклад.
ЗАКЛЮЧЕНИЕ
По результатам исследований, проведенных в двух наиболее крупных лесопарках Москвы (“Битцевский лес” и “Лосиный остров”), получена комплексная количественная оценка состояния основных компонентов лесных экосистем в зависимости от уровня рекреационной нагрузки. Различия хвойно-широколиственных и широколиственных экосистем на легко и среднесуглинистых дерново-подзолистых почвах проявляются только в количественных значениях показателей, характеризующих изменения компонентов лесных экосистем с возрастанием рекреационной нагрузки, но не влияют на их направленность.
Полученные данные позволили уточнить предположение о большей чувствительности растительности к рекреационному влиянию по сравнению с почвами. Оно было сделано на основании того, что некоторые показатели состояния растительности изменяются уже при слабой нагрузке (вторая СД), а статистически значимые изменения плотности почв, которую чаще других свойств используют для характеристики рекреационных изменений, проявляются только к четвертой и пятой стадиям [17]. Анализ зависимости не только плотности, но и других физических, химических и биологических свойств почв от уровня рекреационной нагрузки показал, что они характеризуются разной чувствительностью. Среди них есть свойства с высокой чувствительностью, для которых статистически значимые изменения фиксируются уже при слабой рекреационной нагрузке при переходе от первой ко второй СД (твердость почв, характеристики ее структуры).
Таким образом, предположение о большей чувствительности растительности к рекреационному воздействию по сравнению с почвами не совсем корректно, так как структура почвы, от которой зависит ее водный, воздушный и питательный режимы, является высокочувствительной характеристикой. Рекреация влияет на все компоненты леса, они тесно взаимосвязаны, каждый из компонентов характеризуется показателями с разной чувствительностью, поэтому сложно определить, какой из них наиболее чувствителен.
Полученные зависимости показателей состояния компонентов лесных экосистем от уровня рекреационного воздействия являются нелинейными, что согласуется с результатами других исследователей [33].
Представленные в работе материалы могут быть полезны для прогнозирования динамики состояния лесопарков, определения их рекреационного потенциала и выбора оптимального управления.
Список литературы
Ананьева Н.Д., Сусьян Е.А., Рыжова И.М., Бочарникова Е.О., Стольникова Е.В. Углерод микробной биомассы и микробное продуцирование двуокиси углерода дерново-подзолистыми почвами постагрогенных биогеоценозов и коренных ельников Южной тайги (Костромская область) // Почвоведение. 2009. № 9. С. 1108–1116.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Бурова Н.В., Феклистов П.А. Антропогенная трансформация пригородных лесов. Архангельск: Изд-во Арханг. гос. ун-та, 2007. 264 с.
Вадюнина А.Ф., Корчагина 3.А. Методы исследования физических свойств почв. М.: Агропромиздат, 1986. 416 с.
География и мониторинг биоразнообразия. М.: Изд-во НУМЦ, 2002. 432 с.
Гиляров М.С. Учет крупных почвенных беспозвоночных (мезофауны) // Методы почвенно-зоологических исследований. М., 1975. С. 12–29.
Горбунов А.С., Цветков П.А. Естественное возобновление в рекреационных сосняках зеленой зоны г. Красноярска // Хвойные бореальной зоны. 2009. № 2. С. 244–248.
Грюнталь С.Ю. Почвенные беспозвоночные в условиях рекреационного лесопользования // Влияние рекреации на лесные экосистемы и их компоненты. Институт лесоведения. М., 2004. С. 215–248.
Догель В.А. Зоология беспозвоночных. М.: Высш. школа, 1981. 606 с.
Ефремов А.Л., Новикова Н.В. Биологические особенности почв хвойных насаждений рекреационной зоны г. Могилев // Почвоведение. 2003. № 10. С. 1228–1234.
Захаров В.М., Чубинишвили А.Т. Мониторинг здоровья среды на охраняемых природных территориях. Центр экологической политики России. Центр здоровья среды. М., 2001. 148 с.
Казанская Н.С., Ланина В.В., Марфенин Н.Н. Рекреационные леса. М.: Лесная промышленность, 1977. 96 с.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Моск. ун-та, 1977. 312 с.
Карпиносова Р.А. Дубравы лесопарковой зоны Москвы. М.: Наука, 1967. 104 с.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Кузнецов В.А., Рыжова И.М., Стома Г.В. Изменение свойств почв лесопарков Москвы при высоком уровне рекреационной нагрузки // Почвоведение. 2017. № 10. С. 1270–1281.
Кузнецов В.А., Рыжова И.М., Телеснина В.М., Стома Г.В. Количественная оценка влияния рекреации на растительность, подстилку и плотность почв лесопарков Москвы // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2015. № 1. С. 21–29.
Кузнецов В.А., Стома Г.В. Влияние рекреации на лесные городские ландшафты (на примере национального парка “Лосиный остров” г. Москвы) // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2013. № 3. С. 27–33.
Кузнецов В.А., Стома Г.В., Бодров К.С. Состояние сообщества мезопедобионтов в московских лесопарках как индикатор рекреационной нагрузки и формирования импактных зон вдоль тропинок // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2014. № 1. С. 44–52.
Кузнецов В.А., Стома Г.В., Рыжова И.М. Зависимость изменений свойств почв тропинок и их импактных зон в лесопарках Москвы от уровня рекреационного воздействия // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2018. № 2. С. 19–29.
Мозолевская Е.Г., Соколова Э.С., Жеребцова Г.П., Белов Д.А., Белова Н.К. Оценка жизнеспособности деревьев и правила их отбора и назначения к вырубке и пересадке. Учебно-методическое пособие. 2-е изд. МГУЛ, 2007. 40 с.
Ниценко А.А. Об изучении экологической структуры растительного покрова // Бот. журн. 1969. Т. 54. № 7. С. 1002–1014.
ОСТ 56-100-95. Методы и единицы рекреационных нагрузок на лесные природные комплексы. М., 1995. 14 с.
Рыжова И.М., Подвезенная М.А. Пространственная вариабельность запасов органического углерода в почвах лесных и степных биогеоценозов // Почвоведение. 2008. № 12. С. 1429–1437.
Рысин Л.П., Рысин С.Л. Урболесоведение. М.: КМК, 2012. 240 с.
Серебряноборское опытное лесничество: 65 лет лесного мониторинга / Отв. ред. Б.Р. Стриганова, А.А. Сирин. М.: КМК, 2010. 260 с.
Теория и практика химического анализа почв / Под ред. Л.А. Воробьевой. М., 2006. 400 с.
Cole D.N. Impacts of hiking and camping on soils and vegetation // Environmental Impacts of Ecotourism / Ed. R. Buckley. CABI Publishing, N.Y., 2004. P. 41–60.
Killham K. A physiological determination of the impact of environmental stress on microbial biomass // Environmental Pollution. 1985. V. 38. P. 283–294.
Kissling M., Hegetschweiler K.T., Rusterholz H.-P., Baur B. Short-term and long-term effects of human trampling on above-ground vegetation, soil density, soil organic matter and soil microbial processes in suburban beech forests // Applied Soil Ecology. 2009. V. 42. P. 303–314.
Malmivaara-Lämsä M. Effects of recreational use and fragmentation on the understory vegetation and soil microbial communities of urban forests in southern Finland. Doctoral dissertation. Univ. ofHels. Fac. OfBiosci. 2008. 39 p.
Mariella M., Norman D. Recreational use of forests and disturbance of wildlife – A literature review, Edinburgh, Silvan House, 2012. 52 p.
Monz C.A., Pickering C.M., Hadwen W.L. Recent advances in recreation ecology and the implications of different relationships between recreation use and ecological impacts // Frontiers in Ecology and the Environment. 2013. V. 11. P. 441–446.
Muderrisoglu H., Sarginci M., Toprak B., Uzun S. Effects of recreational usage-type and density on forest floor organic matter in Abant nature park // 1st International Turkey & Japan Environment and Forestry Symposium. Osman Turan Congress and Culture Center Trabzon. 2010. P. 1562–1574.
Soil Survey Division Staff. Soil survey manual. Soil Conservation Service. U.S. Department of Agriculture Handbook 18. U.S. Gov. Printing Office, Washington, DC. 1993. 25 p.
Дополнительные материалы отсутствуют.
Инструменты
Почвоведение