

Почвоведение, 2020, № 1, стр. 119-130
Динамика микробной биомассы, ее структура и функциональная активность в почвах при лесовозобновлении на вырубках пихтарников Енисейского кряжа
А. В. Богородская a, *, А. С. Шишикин a
a Институт леса им. В.Н. Сукачева Сибирского отделения РАН – обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: anbog@ksc.krasn.ru
Поступила в редакцию 19.04.2019
После доработки 28.05.2019
Принята к публикации 20.06.2019
Аннотация
Изучена динамика микробной биомассы, ее структура и функциональная активность в почвах вырубок пихтарников южной тайги Енисейского кряжа (Красноярский край), находящихся на разных стадиях естественных восстановительных сукцессий растительности. В верхних органо-минеральных горизонтах дерново-подзолистой почвы (Albic Glossic Retisol (Loamic, Cutanic, Ochric)) сукцессионной серии вырубок, восстанавливающихся через смену пихтарников на производные березняки, выявлен рост содержания углерода микробной биомассы (Смик) и интенсивности базального дыхания от свежей вырубки к стадии лиственного жердняка, в профиле которого наиболее высокие общие запасы Смик (170 г С/м3) и микробная продукция СО2 (528 мг СО2–С/(м3 ч)), превышающие контроль на 80–85%. В гумусово-аккумулятивном горизонте названного ряда вырубок отмечена тенденция снижения как доли грибного субстрат-индуцированного дыхания, так и отношения грибы/бактерии по сравнению с контролем. В средневозрастных лиственных насаждениях с темнохвойным подростом все рассматриваемые параметры микробных комплексов снижаются, но продолжают превышать контроль. В профиле дерново-подзолистой почвы лесовосстановительной серии без смены пород за 8-летний период отмечен тренд уменьшения Смик и базального дыхания и уже на стадии пихтового жердняка приближение всех эколого-функциональных параметров микробного комплекса к контрольному мелкотравно-зеленомошному пихтарнику.
ВВЕДЕНИЕ
Лесозаготовительные мероприятия являются одним из главных антропогенных факторов, нарушающих экологическую обстановку и запускающих процесс вторичных сукцессий лесов, направленность которых определяются лесорастительными условиями и технологией лесозаготовки [12, 17, 34].
Влияние сплошной рубки леса на почвы определяется трансформацией режимов света, тепла, влаги, увеличением запасов и изменением химического состава растительного опада, а также влиянием лесозаготовительной техники [5–7, 12–15, 18, 21–23, 33]. Отмечено, что эволюция почвенных свойств зависит от степени повреждения почвы, направленности и скорости восстановительных сукцессий растительности [13, 23, 32].
Вторичные послерубочные сукцессии почв по глубине воздействия предложено разделить на поверхностные и турбогенные [33]. К первому подтипу относят динамику почв в ходе естественного возобновления растительности на пасеке, где нет прямого физического воздействия лесозаготовительной техники; к турбогенным – изменения при возобновлении растительности на трелевочных волоках, погрузочных площадках и других элементах лесозаготовительной инфраструктуры, трансформирующих верхние горизонты почв и приводящие к переуплотнению и перемешиванию подстилок, порубочных остатков и минеральных горизонтов почв [12]. На примере сильноминерализованных участков свежих вырубок пихтарников южной тайги Енисейского кряжа (Красноярский край) показаны последствия механизированных лесозаготовок: в первые годы ухудшаются водно-физические свойства почв, снижаются запасы гумуса и доступных форм питательных элементов, происходит резкое изменение активности функционирования микробных комплексов почв и направленности процессов трансформации органического вещества: численность гетеротрофных микроорганизмов почвы снижена в 10–15 раз, а активность почвенных ферментов – в 4–10 раз [5, 26]. Морфологические и физико-химические изменения свойств почв на механически нарушенных участках лесосек могут сохраняться до 100 и более лет [13, 18, 33].
Вырубки и вывоз древесины в зимний период приводят к меньшим нарушениям не только наземного растительного покрова, но и верхних органогенных и органо-минеральных горизонтов почвы [35].
Лесовосстановление пихтовых вырубок южной тайги Енисейского кряжа чаще идет через смену пород (береза), под пологом которой примерно к 20-ти годам формируется подрост темнохвойных пород [6]. В первые годы после рубки роль основного эдификатора ценоза переходит к растениям напочвенного покрова [12]. Увеличивается поступление азота, кальция, магния и других элементов в составе быстро минерализуемых компонентов травянистого и лиственного опада и скорости его вовлечения в биогеохимический цикл. До смыкания крон древостоев лидирующая роль в поступлении растительного опада остается за растениями напочвенного покрова, состав и проективное покрытие которых изменяются в зависимости от микроклиматических условий и типа вырубок [14, 32].
Для ранней диагностики состояния почв при антропогенном или техногенном нарушении и определения критических пределов ее сбалансированного функционирования в настоящее время применяют биологические параметры [26]. Содержание микробной биомассы в почве, ее структуры (соотношение грибов и бактерий) и функциональной активности (базальное дыхание) являются основными параметрами в экологических исследованиях, в том числе и при различных сценариях лесопользования [1, 10, 25, 29, 40].
Цель исследований заключалась в изучении динамики микробной биомассы, соотношения в ней эукариотного и прокариотного компонентов и ее минерализационной активности в почвах вырубок пихтарников южной тайги Енисейского кряжа, находящихся на разных стадиях естественных восстановительных сукцессий растительности.
ОБЪЕКТЫ И МЕТОДЫ
Исследования разновозрастных сплошнолесосечных вырубок проводили в Приенисейской провинции Приангарского понижения Енисейского кряжа [20]. По лесорастительному районированию территория находится в северной части Южно-Приенисейского округа южно-таежных сосновых и темнохвойных лесов [16]. Округ расположен в самой южной части Приенисейской провинции на правобережье р. Енисей, где преобладают полидоминантные спелые и перестойные темнохвойные леса, в основном представленные пихтовыми насаждениями с фрагментами перестойных кедровников. В последние 50 лет леса округа интенсивно эксплуатируются.
Среднегодовая температура воздуха на исследуемой территории изменяется в пределах от минус 2.6 до плюс 0.9°С, средняя температура января – от минус 23.0 до минус 17.0°С, июля – от 16.1 до 19.9°С [28]. Среднегодовое количество осадков достигает 640 мм, около 70% из них выпадает в жидком виде [27]. Мощный снежный покров (90–180 см) предохраняет почвы от глубокого промерзания [9].
Микробиологические исследования почв на вырубках проводили в 2008–2016 гг. на постоянных пробных площадях (ПП) (58°27′ N, 93°16′ E) в процессе выполнения комплексных (гидрологических, почвенных, геоботанических, лесоводственных, зоологических) исследований.
Вырубки подбирали для двух сукцессионных серий: с сохранением темнохвойного подроста и без сохранения – с лиственным возобновлением и последующей подпологовой сменой на темнохвойные породы. Возрастной ряд вырубок включал свежие лесосеки, лиственные несомкнувшиеся и сомкнувшиеся молодняки, лиственный и темнохвойный жердняки (35 лет), а также средневозрастные темнохвойные и лиственные насаждения. Для сравнения сукцессионных процессов на вырубках с фоновым состоянием насаждений подобран пихтарник мелкотравно-зеленомошный (табл. 1). Рубку проводили по одной технологии: агрегатными машинами, сплошная, пасеками, зимняя с площадью лесосек 150–200 га.
Таблица 1.
Основные характеристики пробных площадей
| № ПП | Год проведения рубки | Тип сообщества | Состав древостоя* | Мощность подстилки**, см |
|---|---|---|---|---|
| 1 | 2008 | Разнотравно-вейниковый лесной луг с лиственным возобновлением | 10Б + ПЕ | 6/2.5 |
| 2 | 2003 | Березняк кипрейно-вейниково-малинниковый (молодняк) | 10Б + ПЕ | 5/4 |
| 3 | 1998 | Березняк кипрейно-вейниковый (молодняк) | 10Б + ОсЕПс** | 7/6 |
| 4 | 1978 | Березняк вейниково-разнотравный с микрогруппировками зеленых мхов (жердняк) | 6Б3П1Пс | 3/3 |
| 5 | 1950 | Осинник с пихтой во втором ярусе разнотравный | 10Ос + Б, II8ПЕК | –/2 |
| 6 | 1978 | Пихтарник с березой разнотравно-зеленомошный (жердняк) | 7П3Б + Пс | 2.5/2 |
| 7 | 1950 | Пихтово-еловый разнотравно-вейниковый | 8П2Е + К | –/1.6 |
| Контроль | Более 150 | Пихтарник мелкотравно-зеленомошный | 10П + КОсБ | 1.5/2 |
Изучаемые насаждения формируются на дерново-подзолистых тяжелосуглинистых почвах (Albic Glossic Retisol (Loamic, Cutanic, Ochric)) [19, 34]. Профиль почв четко дифференцирован по генетическим горизонтам: О–AY–EL–BEL–BT–C.
Для микробиологических анализов в конце августа–начале сентября из почвенных прикопок отбирали образцы послойно до 50 см из органогенного горизонта (О), гумусово-аккумулятивного (AY) и минеральных горизонтов (EL(AEL), BEL и ВТ). Прикопки располагали на пасеках, где влияние лесозаготовительной техники было минимальным.
Отбор почвенных образцов сопровождали определением влажности почвы на момент взятия образцов (термовесовым методом), температуры почвенных слоев (портативным термометром Checktemp) и плотности (буром Качинского). Для анализов использовали свежие образцы, хранившиеся при +5°С не более 2–3 недель.
Определяли содержание микробной биомассы (Смик) методом субстрат-индуцированного дыхания [1, 36]. В стеклянные флаконы (250 мл) помещали 2 г почвы без корней или 1 г подстилки без крупных растительных остатков, добавляли 0.2 мл глюкозо-минеральной смеси (ГМС, мг/мл: глюкоза – 200; К2НРО4 – 20; (NH4)2SO4 – 20), герметично закрывали резиновыми пробками и инкубировали при 22°С в течение 3–4 ч, что соответствует лаг-периоду роста микробной популяции [1]. Смик определяли путем пересчета скорости субстрат-индуцированного дыхания (СИД) по формуле [36]:
Базальное дыхание (БД) почвы измеряли по скорости выделения СО2 почвой за 24 ч инкубации при 25°С [1]. Определение СО2 проводили хроматографически, как описано для определения СИД, только вместо раствора ГМС вносили воду. Интенсивность базального дыхания выражали в мкг СО2–С/(г ч).
БД и СИД определяли по разности концентраций СО2 в начале и конце инкубации при помощи газового хроматографа Agilent Technologies 6890N, снабженного пламенно-ионизационным детектором и метанатором (Hewlett-Packard, США). Во время анализа использовали колонку Supelco 10182004 из нержавеющей стали с внутренним диаметром 3.175 мм, длиной 1828.8 мм. Адсорбент – 80/100 Porapak Q. Рабочие параметры хроматографа Agilent 6890N: температура термостата колонки – 80°С, поток газа-носителя (гелия) – 20 мл/мин. Режим работы пламенно-ионизационного детектора: температура детектора – 300°С, температура заднего порта – 375°С, поток водорода – 30 мл/мин, поток воздуха – 400 мл/мин. Объем вводимой газовой пробы 5 мл.
Для разделения вклада грибов и бактерий в микробную биомассу применяли метод селективного ингибирования [3, 30], основанный на угнетении СИД грибов или бактерий под действием селективных антибиотиков [39]. Оценку грибного и бактериального СИД проводили в образцах почв, отобранных на вырубках в 2016 г. и прошедших предварительную инкубацию в течение 5 суток [3, 29]. Стрептомицина сульфат (водный раствор) и циклогексимид (порошок с тальком в соотношении 1 : 2) вносили в почву по отдельности и вместе, добавляли ГМС и измеряли СИД. Циклогексимид (фунгицид) в различных концентрациях добавляли в почву за 4 ч до внесения глюкозы, а стрептомицин (бактерицид) – за 0.5 ч. При совместном внесении антибиотиков циклогексимид инкубировали с почвой 4 ч, затем добавляли стрептомицин и через 0.5 ч вносили глюкозу. Образец почвы, в который была внесена только глюкоза с тальком, служил контролем [30].
Коэффициент перекрывания активности антибиотиков (ПАА) или дополнительного ингибирования рассчитывали по уравнению:
Соотношение грибного и бактериального вкладов в СИД определяли по формулам:
при условии, что A – [(A – B) + (A – C)] = D ± 5–8% [24, 30].
Рассчитывали запас углерода микробной биомассы Смик (г С/м3) и микробное продуцирование СО2 (мг СО2–С/(м3 ч)) в исследуемых горизонтах почвы как произведение содержания Смик (г/г почвы) или интенсивности БД (мг/г почвы), плотности почвы (ρ, г/см3) и объема почвы в данном слое (V, м3). Суммарные запасы Смик и общее микробное продуцирование СО2 в профиле почв получали суммированием полученных данных для каждого исследуемого горизонта (до 50 см) [29, 31].
Все определения выполняли в трех повторностях и рассчитывали стандартное отклонение от среднего.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Содержание углерода микробной биомассы и интенсивность базального дыхания в почвах разновозрастных вырубок
Морфологическое строение и основные свойства дерново-подзолистой почвы изучаемого мелкотравно-зеленомошного пихтарника типичны для дерново-глубокоподзолистых почв, господствующих в Приангарском районе Енисейского кряжа [11]. Почвы кислые (рН 4.7–4.4), содержание гумуса в горизонте AY достигает 6.7%, постепенно снижаясь до 5% в гор. AYel и 2% в горизонте EL на глубине 30 см. Аналогично гумусу по профилю распределяется валовой азот, наибольшее его содержание (545 мг/100 г) обнаружено в подстилке, при переходе к горизонту AY – сокращается в 2 раза. Гумус слабо обогащен азотом, отношение C/N для горизонта AY не меньше 15, в горизонте EL – 16–20 [34]. В почвенном профиле контрольного пихтарника отмечена низкая годовая вариабельность динамики содержания углерода микробной биомассы: в подстилке Смик достигало 2960–3180 мкг С/г, в горизонте АY – 1100–1260 мкг С/г и снижалось в горизонте АYel до 280–320 мкг С/г (рис. 1).
Рис. 1.
Динамика содержания углерода микробной биомассы и интенсивности базального дыхания в дерново-подзолистой почве вырубок со сменой хвойной породы на лиственные и без смены – темнохвойные. Здесь и далее по оси X указан ПП (год проведения рубки). Стадии сукцессии: Тр – травянистая; Мл – молодняки; Жд – жердняки; Ср – средневозрастные.
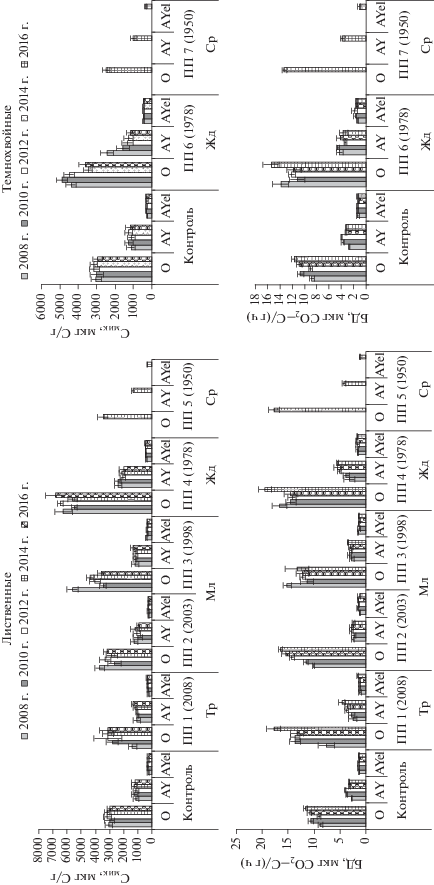
Наибольшая динамичность содержания Смик за 8-летний период обнаружена в подстилке и минеральных горизонтах почвы свежей вырубки и хвойного жердняка. В первый год после вырубки пихтарников снижается содержание Смик в подстилке на 55% и гумусово-аккумулятивном горизонте дерново-подзолистой почвы на 10%, при этом интенсивность базального дыхания не изменяется. На травянистой стадии за 8-летний период лесовосстановления содержание Смик увеличилось в 2.3 раза и достигло контроля к стадии молодняка, тогда как в горизонте АY за 8 лет увеличилось на 30% и составило 1340 мкг С/г, что на 13% выше контроля.
В несомкнувшемся молодняке (ПП 2) за 8-летний период развития фитоценозов и подроста березы к стадии сомкнутых молодняков отмечено незначительное снижение Смик в разных почвенных горизонтах. В процессе смыкания крон березового молодняка (ПП 3) за 8-летний период отмечено снижение Смик с 5600 до 3550 мкг С/г в подстилке и возрастание Смик до 1320 мкг С/г в гумусово-аккумулятивном горизонте. При смыкании древесных крон снижается количество поступающего растительного опада, изменяются гидротермические условия почв. Биотрансформация подстилки и рыхлой дернины обусловливает аккумуляцию гумуса в верхней части минерального профиля, мощность горизонта AY достигает 6 см, содержание гумуса повышается до 9% [34]. В пихтовом жердняке (ПП 6) за 8 лет сукцессии растительности уровень микробной биомассы сократился на 20% в подстилке и в 2 раза в гумусово-аккумулятивном горизонте AY и сравним с контролем. В этом же горизонте почвы лиственного жердняка (ПП 4) Смик уменьшается на 15% за 8 лет сукцессии, тогда как в других горизонтах почвы возрастает на 10%. В целом в почве лиственного жердняка отмечено наибольшее содержание Смик – 6800 и 2000 мкг С/г в подстилке и горизонте AY соответственно, что в 2 и более раз выше контроля.
Таким образом, уровень микробной биомассы в подстилке и в гумусово-аккумулятивном горизонте дерново-подзолистой почвы лиственной сукцессионной серии превышает контроль на травянистой вырубке (ПП 1), сомкнувшемся молодняке (ПП 3), жердняке (ПП 4), а также в средневозрастном лиственном насаждении с темнохвойным подростом (ПП 5).
Интенсивность базального дыхания в почве разновозрастных вырубок за 8-летний период варьировала больше, чем Смик. Максимальный рост интенсивности БД за 8-летний период обнаружен на травянистой стадии вырубки (ПП 1), где величина БД в подстилке и в горизонте AY возрастала более, чем в 2 раза и достигла 17.8 и 4.7 мкг СО2–С/(г ч) соответственно. Это связано с интенсивной минерализацией травянистого и лиственного опада, что также отмечено для почв молодых вырубок [34, 42, 44, 45]. В подстилке несомкнувшегося молодняка (ПП 2) интенсивность БД за 8‑летний сукцессии к стадии сомкнутых молодняков возросла на 60% и осталась без изменения в нижележащих горизонтах почвы. В подстилке сомкнувшегося молодняка (ПП 3) величина БД несколько снижается, тогда как в горизонте AY незначительно увеличивается. В лиственном жердняке (ПП 4) интенсивность БД также повышается к 2016 г., особенно заметно в горизонте AY – на 70%, достигая максимальной величины 5.5 мкг СО2–С/(г ч), что в 1.7 раз выше контроля.
Таким образом, наибольшая скорость БД в 2016 г. отмечена в подстилке 8-летней вырубки (ПП 1), лиственного жердняка (ПП 4) и средневозрастного осинника с темнохвойным подростом (ПП 5). При прямом возобновлении темнохвойных пород (ПП 6, 7), БД ниже и сравнимо с контролем.
Величины Смик и БД тесно коррелировали с влажностью почв вырубок (R = 0.79–0.84), имели слабую корреляцию с температурой почв (R = = 0.34–0.49).
Рост уровня микробной биомассы и интенсивности БД в подстилках сукцессионной серии вырубок, восстанавливающихся через смену породы на производные березняки, имел логарифмический тренд от свежей вырубки к стадии жердняка и экспоненциальный – в гумусово-аккумулятивном горизонте дерново-подзолистой почвы для Смик, где интенсивность БД возрастала линейно (рис. 2). В нижележащем почвенном горизонте тенденция увеличения рассматриваемых параметров выражена не отчетливо. В средневозрастных лиственных насаждениях с темнохвойным подростом названные параметры имели тенденцию к снижению.
Рис. 2.
Рост содержания Смик (А) и интенсивности БД (Б) в почве разновозрастных вырубок, восстанавливающихся через смену пихты на производные березняки. На оси Y указаны контрольные значения.
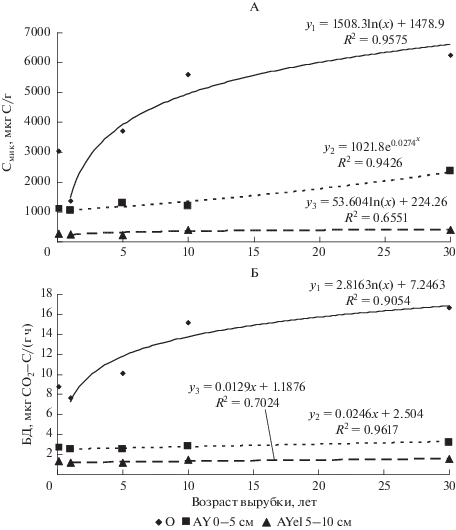
В целом по уровню содержания Смик и интенсивности БД почвы пихтового жердняка и средневозрастного елово-пихтового леса наиболее приближены к контрольному мелкотравно-зеленомошному пихтарнику. На вырубках, восстановительная сукцессия которых идет со сменой на производные березняки, преобладают дерновые процессы, отмечается значительное повышение содержания Смик и скорости БД, поскольку увеличивается напряженность минерализационных процессов при изменении гидротермических условий почвы вырубок и смене хвойного на лиственный опады [4, 7, 12, 22, 26, 38]. Известно, что опад лиственных пород и травянистых растений гораздо богаче зольными элементами и азотом, чем хвоя, что способствует быстрой минерализации и вовлечению в биотический круговорот [7, 45]. Изменение качества и количества растительного опада на вырубках, восстанавливающихся лиственными породами, приводит к снижению кислотности (рН 5.2–6), увеличению содержания гумуса на 50% в горизонте AY почвы на стадии жердняка, где оно достигало 10.2%. Также на стадии жердняка в 3 раза повышается содержание валового азота, при этом доля легкоминерализуемой фракции азота снижается, что является результатом интенсивной минерализации его компонентов [34].
Соотношение эукариотных и прокариотных микроорганизмов в общем Смик в почвах вырубок
Для определения доли грибов и бактерий в составе активной микробной биомассы почвы применяли метод селективного ингибирования антибиотиками [39]. Ранее отмечено, что в почвах экосистем с высоким содержанием микробной биомассы необходимо применение высоких доз антибиотиков [3, 24, 30], поэтому в предварительных экспериментах по подбору оптимальных доз антибиотиков использовали концентрации циклогексимида от 30 до 60 мг/г, а стрептомицина – от 10 до 20 мг/г. Оптимальные концентрации антибиотиков, при которых достигалось максимальное снижение СИД и ПАА = 1 ± 8%, составляли для циклогексимида 40 мг/г (49–68% подавления общего СИД) и стрептомицина – 20 мг/г (13–29% подавления общего СИД). При совместном внесении использовали концентрацию циклогексимида 30 мг/г и стрептомицина 20 мг/г.
В гумусово-аккумулятивном горизонте почв исследованного сукцессионного ряда вырубок пихтарников обнаружен доминантный вклад грибов (73–83%) в общее СИД, тогда как на долю бактериального СИД приходилось 19–35% (табл. 2). Уменьшение величины СИД почв при последовательном внесении двух антибиотиков достигало 66–85% от контроля, что укладывается в диапазон ранее установленных величин [2, 24, 29, 30, 39].
Таблица 2.
Вклад грибов и бактерий в субстрат-индуцированное дыхание гумусово-аккумулятивного горизонта дерново-подзолистой почвы вырубок разных сукцессионных стадий
| № ПП | Стадия сукцессии* | Грибы, % | Бактерии, % | Ц + С**, % | ПАА*** | $\frac{{{{{\text{C}}}_{{{\text{грибов}}}}}}}{{{{{\text{C}}}_{{{\text{бактерий}}}}}}}$ |
|---|---|---|---|---|---|---|
| Контроль | 81 ± 1.5 | 21 ± 1 | 73 ± 2 | 1.02 | 3.9 | |
| 1 | Тр | 76 ± 4 | 30 ± 2 | 66 ± 3 | 1.07 | 2.6 |
| 3 | Мл | 77 ± 4 | 35 ± 3 | 75 ± 3 | 1.12 | 2.3 |
| 4 | Жд | 73 ± 5 | 36 ± 3 | 80 ± 4 | 1.09 | 2.2 |
| 6 | 80 ± 3 | 31 ± 1 | 85 ± 3 | 1.11 | 2.6 | |
| 5 | Ср | 78 ± 3 | 27 ± 2 | 68 ± 2 | 1.05 | 2.9 |
| 7 | 83 ± 2 | 19 ± 1 | 67 ± 1 | 1.02 | 4.4 | |
* Обозначения см. рис. 1. ** Циклогексимид + стрептомицин. *** Коэффициент перекрывания активности антибиотиков.
Вклад грибов в общее СИД лесных почв европейской территории России разных растительных подзон для верхнего минерального горизонта составил 52–99% [29]. Было показано, что доля грибов в гумусовом горизонте (0–10 см) естественных экосистем составила 60, 81, 70 и 95%, для дерново-подзолистой (лес), серой лесной (лес), чернозема (целина) и каштановой (залежь) почв соответственно [2, 25, 30]. Другими авторами показано, что на долю грибного дыхания в пахотных, луговых и лесных почвах приходится в среднем 61–65% [43].
Обнаружена тенденция снижения доли грибного СИД в ряду лесовосстановления вырубок через смену на производные березняки: в почве 8‑летней вырубки на травянистой стадии с подростом березы доля грибов снижалась до 76% против 81% на контроле и составляла 73% на стадии жердняка, тогда как в средневозрастном лиственном насаждении с темнохвойным подростом достигала почти 78%. В почве темнохвойных вырубок на стадии жердняка и средневозрастного насаждения составляла 80–83%.
На основании полученного долевого вклада грибов и бактерий в общее СИД рассчитано соответственно грибное и бактериальное Смик (рис. 3А). Содержание грибного Смик в гумусово-аккумулятивном горизонте дерново-подзолистой почвы контрольного пихтарника составляло 960 мкг С/г и увеличивалось до 1020–1450 мкг С/г на вырубках всех сукцессионных стадий лиственного возобновления, достигая максимума в лиственном жердняке. В темнохвойном жердняке и средневозрастном насаждении этот показатель сравним с контролем. Содержание бактериального Смик варьировало больше (195–650 мкг С/г) и составляло 250 мкг С/г в горизонте АY контрольного пихтарника, увеличиваясь в 1.5–3 раза на разных стадиях лиственного возобновления и приближена к контролю – в темнохвойной серии.
Рис. 3.
Вклад грибов (1) и бактерий (2) в Смик (А) и БД (Б) гумусово-аккумулятивного горизонта дерново-подзолистой почвы разновозрастных вырубок.
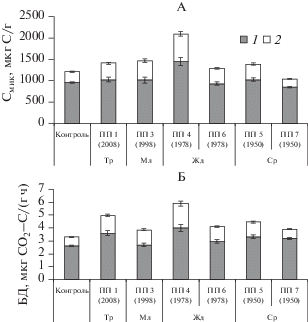
В горизонте АY дерново-подзолистой почвы вырубок на травянистой стадии, молодняках и лиственных жердняках происходит снижение отношения биомассы Сгрибы/Сбактерии до 2.2–2.6 против 3.9 в почве контрольного пихтарника. В средневозрастном темнохвойном насаждении (ПП 7) отношение Сгрибы/Сбактерии превышает контроль и достигает 4.4. Доминирование грибной микрофлоры и увеличение соотношения грибы : бактерии в почве хвойных лесов отмечено ранее и связано с преобладанием в опаде бактерицидных веществ и кислых субстратов [29, 37]. На отношение грибы : бактерии влияет кислотность почв [24, 46], а также сочетание кислотности почв и растительного опада [41].
На основе вклада грибного и бактериального в общее СИД было дифференцировано грибное и бактериальное продуцирование СО2 гумусовым горизонтом дерново-подзолистых почв вырубок (рис. 3Б). Вклад грибов в общее микробное продуцирование СО2 был наибольшим в почве на травянистой стадии вырубки и лиственного жердняка, тогда как в темнохвойных жердняке и средневозрастных насаждениях – приближены к контролю.
Запасы углерода микробной биомассы и микробное продуцирование СО2 в профиле почв вырубок
Запас микробной биомассы примерно одинаков в разных минеральных горизонтах почвенного профиля дерново-подзолистой почвы контрольного пихтарника мелкотравно-зеленомошного (29.6–30.8 г С/м3), тогда как максимум микробного продуцирования СО2 приходится на наиболее мощный элювиальный горизонт EL–BEL (рис. 4).
Рис. 4.
Запасы углерода микробной биомассы (А) и микробное продуцирование СО2 (Б) разных горизонтов (мощность, см) дерново-подзолистой почвой вырубок лиственной (ПП 1, 3, 4, 5) и темнохвойной (ПП 6, 7) сукцессионных серий.
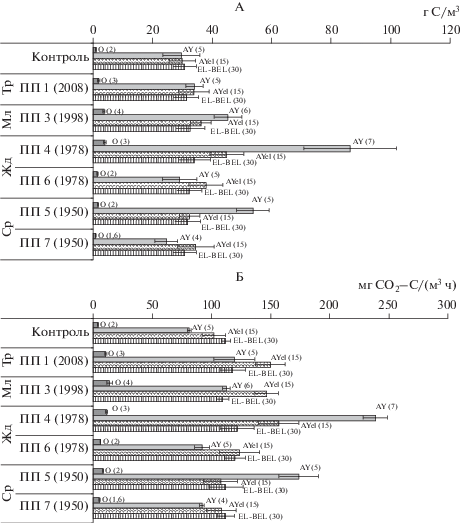
Наименьший вклад (не более 2–3%) в профильные запасы Смик и микробной продукции СО2 вносила подстилка контрольного пихтарника, несмотря на максимальное содержание в ней Смик и скорости БД, что связано как с низкой плотностью растительных подстилок, так и их маломощностью. Суммарные запасы углерода микробной биомассы и общее продуцирование СО2 в профиле до 50 см дерново-подзолистой почвы пихтарника составляли 91 г С/м3 и 300 мг СО2–С/(м3 ч), что несколько ниже, чем в дерново-подзолистых почвах светлохвойных лесов Средней Сибири [8] и в 2–10 раз ниже, чем в дерново-подзолистых и серых лесных почвах в разных типах южно- и среднетаежных лесов европейской части России [10, 29, 31], что является следствием как меньшей мощности гумусового горизонта, так и влиянием климатических особенностей региона.
Для разных сукцессионных серий восстановления растительности на вырубках характерна определенная трансформация профильного распределения запасов Смик и микробной продукции углекислого газа (рис. 4). На 8-летней вырубке на травянистой стадии лиственного возобновления (ПП 1) изменений в профильных запасах Смик не выявлено, но микробная продукция СО2 подстилки возрастала более, чем в 2 раза, а в горизонтах АY, АYel увеличивалась почти на 50% от контроля и достигала 120–150 мг СО2–С/(м3 ч). В подстилке сомкнутого березового молодняка (ПП 3) запас Смик больше контроля почти в 4 раза, а его вклад в суммарные запасы почвенного профиля достигает 12%. Микробное продуцирование углекислого газа подстилкой молодняка также выше контроля в 3.5 раза. Запас Смик в горизонтах АY, АYel увеличивается до 36–45 г С/м3, что на 20–50% больше контроля, а продуцирование СО2 выше контроля на 40%. Максимальное увеличение запаса Смик и продукции СО2 (в 2–4 раза от контроля) наблюдается в профиле дерново-подзолистой почвы лиственного жердняка (ПП 4), при этом наибольшее увеличение приходится на гумусово-аккумулятивный горизонт, где названные величины достигали 86 г С/м3 и 240 мг СО2–С/(м3 ч), что составляло 45–50% от общего запаса Смик и продукции СО2 на весь рассматриваемый профиль почвы. В средневозрастном лиственном насаждении (ПП 5) запас Смик выше контроля на 80% в горизонте АY, тогда как в других почвенных горизонтах увеличен незначительно. В свою очередь микробная продукция СО2 подстилкой и горизонтом АY осиново-березового насаждения выше контроля более, чем в 2 раза и достигала 8.5 и 180 мг СО2–С/(м3 ч).
В темнохвойных жердняке и средневозрастном насаждении изменений в профильном распределении запаса Смик и микробной продукции СО2 не наблюдалось, отмечено незначительное повышение продукции СО2 разными горизонтами пихтового жердняка относительно контроля.
Суммарные запасы углерода микробной биомассы и общая микробная продукция СО2 в профиле до 50 см дерново-подзолистой почвы увеличивались при лиственном возобновлении, достигая максимальных величин в березовом жердняке (170 г С/м3 и 530 мг СО2–С/(м3 ч)), что выше контроля на 80–85% (рис. 5). В профиле почвы средневозрастного лиственного насаждения с темнохвойным подростом общие запасы Смик и продукции СО2 снижаются, но остаются выше контроля на 30–35%. При темнохвойном возобновлении суммарные запасы Смик и микробной продукции СО2 находятся на уровне контроля.

ЗАКЛЮЧЕНИЕ
В дерново-подзолистой почве контрольного пихтарника мелкотравно-зеленомошного содержание Смик в подстилке достигало 2960–3180, в горизонте АY – 1100–1260 и АYel – 280–320 мкг С/г; интенсивность БД соответственно варьировала больше 8.8–11.7, 2.8–4.0, 1.2–1.4 мкг СО2–С/(г ч). В первый год после вырубки пихтарников снижается содержание Смик в подстилке на 55% и гумусово-аккумулятивном горизонте дерново-подзолистой почвы на 10%, при этом интенсивность базального дыхания не изменяется. В верхних органо-минеральных горизонтах дерново-подзолистой почвы сукцессионной серии вырубок, восстанавливающихся через смену пихтарников на производные березняки, отмечен рост содержания Смик и интенсивности БД от свежей вырубки к стадии жердняка. На травянистой стадии лесовосстановительных послерубочных сукцессий растительности в горизонте АY увеличивается Смик до 1336 мкг С/г и БД до 4.7 мкг СО2–С/(г ч), на 30% возрастает общая микробная продукция СО2 в профиле дерново-подзолистой почвы. На стадии лиственных жердняков значительно увеличиваются мощность гумусово-аккумулятивного горизонта, содержание в нем азота и гумуса, что коррелирует с содержанием Смик и БД, достигающими максимальных величин (2000–2380 мкг С/г и 3.2–5.5 мкг СО2–С/(г ч)). В почвенном профиле лиственного жердняка наиболее высокие общие запасы Смик (170 г С/м3) и микробная продукция СО2 (528 мг СО2–С/(м3 ч)), превышающие контроль на 80–85%. В гумусово-аккумулятивном горизонте почв вырубок пихтарников доминировал вклад грибов (73–83%) в общее СИД. Для производных березняков отмечена тенденция снижения как доли грибного СИД, так и отношения Сгрибы/Сбактерии по сравнению с контролем, однако абсолютное содержание грибного Смик увеличивалось на 10–50%, достигая максимума в почве лиственного жердняка (1450 мкг С/г). В горизонте АY темнохвойных жердняка и средневозрастного насаждения доля грибов и содержание грибного Смик сравнимы с контролем. В средневозрастных лиственных насаждениях с темнохвойным подростом все рассматриваемые параметры микробных комплексов снижаются, но продолжают превышать контроль. В профиле дерново-подзолистой почвы лесовосстановительной серии без смены пород за 8‑летний период отмечен тренд снижения Смик и БД и уже на стадии пихтового жердняка приближение всех эколого-функциональных параметров микробного комплекса к контрольному мелкотравно-зеленомошному пихтарнику. Следовательно, лиственное лесовозобновление на вырубках пихтовой формации на 30–40 лет создает благоприятные условия для увеличения почвенного плодородия и создает ландшафтное биоразнообразие.
Выявленные закономерности сукцессий микробного комплекса в процессе лесовозобновления на вырубках темнохвойных насаждений южной тайги Средней Сибири могут использоваться при экспертной оценке лесорастительных условий почв вырубок, при решении вопросов контроля и прогноза состояния лесных экосистем при антропогенном и техногенном воздействии, а также при проведении разного рода лесохозяйственных мероприятий.
Список литературы
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 222 с.
Ананьева Н.Д., Полянская Л.М., Стольникова Е.В., Звягинцев Д.Г. Соотношение биомассы грибов и бактерий в профиле лесных почв // Изв. РАН. Сер. биол. 2010. № 3. С. 308–317.
Ананьева Н.Д., Сусьян Е.А., Чернова О.В., Чернов И.Ю., Макарова О.Л. Соотношение грибов и бактерий в биомассе разных типов почв, определяемое селективным ингибированием // Микробиология. 2006. Т. 75. № 6. С. 807–813.
Аристовская Т.В. Микробиология процессов почвообразования. Л.: Наука, 1980. 187 с.
Бабинцева Р.М., Горбачев В.Н., Сорокин Н.Д. Экологические аспекты лесовосстановления при современных лесозаготовках // Лесоведение. 1984. № 5. С. 19–25.
Батура И.В., Батура А.В. Лесовосстановление пихтовых вырубок южной тайги в лесорастительных условиях Енисейского кряжа // Лесная таксация и лесоустройство. 2005. № 2(35). С. 78–81.
Бахмет О.Н., Медведева М.В. Изменение свойств почв Карелии в процессе искусственного лесовосстановления // Лесоведение. 2013. № 3. С. 38–45.
Богородская А.В., Кукавская Е.А. Состояние микробных сообществ в почвах лиственных и светлохвойных лесов Средней Сибири после рубок и пожаров // Лесоведение. 2016. № 5. С. 383–396.
Буренина Т.А., Шишикин А.С., Онучин А.А. Снежный покров на вырубках разных лет в пихтово-кедровых лесах Енисейского кряжа // Лесоведение. 2013. № 6. С. 26–36.
Гавриленко Е.Г., Ананьева Н.Д., Макаров О.А. Оценка качества почв разных экосистем // Почвоведение. 2013. № 12. С. 1505–1515.
Горбачев В.Н., Попова Э.П. Почвенный покров южной тайги Средней Сибири. Новосибирск: Наука, 1992. 223 с.
Дымов А.А. Влияние сплошных рубок в Бореальных лесах России на почвы // Почвоведение. 2017. № 7. С. 787–798.
Дымов А.А. Изменение почв в процессе естественного лесовостановления (на примере подзолов средней тайги, сформированных на двучленных отложениях). Автореф. дис. … канд. биол. н. Сыктывкар, 2007. 17 с.
Дымов А.А., Милановский Е.Ю. Изменение органического вещества таежных почв в процессе естественного возобновления растительности после рубок (средняя тайга Республики Коми) // Почвоведение. 2014. № 1. С. 39–47.
Дымов А.А. Почвы механически нарушенных участков лесосек средней тайги Республики Коми // Лесоведение. 2018. № 2. С. 130–142.
Жуков А.Б., Коротков И.А., Кутафьев В.П., Назимова Д.И., Реган С.П., Савин Е.Н., Чередникова Ю.С. Леса красноярского края // Леса СССР. М.: Наука, 1969. Т. 4. С. 248–321.
Иванов В.В. Экологические последствия механизированных лесозаготовок в южной тайге Красноярского края // Лесоведение. 2005. № 2. С. 3–8.
Исаченкова Л.Б. Изменение свойств дерново-подзолистых почв в сукцессионных рядах восстановления широколиственно-хвойных лесов (на примере юго-западного Подмосковья). Автореф. дис. … канд. геогр. н. М., 2007. 24 с.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Козловская С.Ф. Енисейский кряж // Плоскогорья и низменности Восточной Сибири. М.: Наука, 1971. С. 46–53.
Краснощеков Ю.Н., Сорокин Н.Д. Почвенно-экологические изменения на вырубках и гарях Восточного Хэнтэя (МНР) // Почвоведение. 1988. № 1. С. 117–127.
Медведева М.В., Кудинова Ю.С. Изменение биологической активности почв в процессе естественного лесовосстановления соснового древостоя // Вестник Поволжского государственного технологического университета. Сер.: Лес. Экология. Природопользование. 2017. № 1(33). С. 17–25.
Орфанитский Ю.А., Орфанитская В.Г. Почвенные условия таежных вырубок. М.: Наука, 1971. 96 с.
Приходько В.Е., Сиземская М.Л. Базальное дыхание и состав микробной биомассы целинных, агро- и лесомелиорированных полупустынных почв Северного Прикаспия // Почвоведение. 2015. № 8. С. 974–983.
Полянская Л.М., Звягинцев Д.Г. Содержание и структура микробной биомассы как показатели экологического состояния почв // Почвоведение. 2005. № 6. С. 706–714.
Сорокин Н.Д. Микробиологическая диагностика лесорастительного состояния почв Средней Сибири. Новосибирск: Изд-во СО РАН, 2009. 222 с.
Справочник по климату СССР. Влажность воздуха, атмосферные осадки и снежный покров. Л.: Гидрометеорологическое издательство, 1969. Вып. 21. Ч. IV. 404 с.
Справочник по климату СССР. Температура воздуха и почвы. Л.: Гидрометеорологическое издательство, 1967. Вып. 21. Ч. II. 504 с.
Стольникова Е.В., Ананьева Н.Д., Чернова О.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов Европейской территории России // Почвоведение. 2011. № 4. С. 479–494.
Сусьян Е.А., Ананьева Н.Д., Благодатская Е.В. Разделение грибного и бактериального субстрат-индуцированного дыхания с использованием антибиотиков в почвах разных экосистем // Микробиология. 2005. Т. 74. № 3. С. 394–400.
Сусьян Е.А., Ананьева Н.Д., Гавриленко Е.Г., Чернова О.В., Бобровский М.В. Углерод микробной биомассы в профиле лесных почв южной тайги // Почвоведение. 2009. № 10. С. 1233–1240.
Титарев Р.П. Особенности восстановления почвенных свойств и растительности на сплошных вырубках в подзоне южной тайги (на примере Боровинского лесничества Калужской области). Автореф. дис. … канд. биол. н. М., 2009. 26 с.
Тощева Г.П. Изменение почвенного покрова на вырубках ельников южной тайги. Автореф. дис. … канд. биол. н. М., 1988. 24 с.
Трефилова О.В., Ефимов Д.Ю. Изменение растительного покрова почв при естественном зарастании вырубок пихтарников Енисейского кряжа // Почвоведение. 2015. № 8. С. 910–920.
Уланова Н.Г. Механизмы сукцессий растительности сплошных вырубок в ельниках Южной тайги // Актуальные проблемы геоботаники. Мат-лы III Всерос. школы-конф. 24–28 сентября 2007. Петрозаводск: Изд-во КарНЦРАН, 2007. С. 199–211.
Anderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. Biochem. 1978. 10. P. 314–322.
Baath E., Anderson T.-H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA_based techniques // Soil Biol. Biochem. 2003. 7. P. 955–963.
Baath E., Frostegard A., Pennanen T., Fritze H. Microbial community structure and pH response in relation to soil organic matter quality in wood-ash fertilized, clear-cut or burned coniferous forest soils // Soil Biol. Biochem. 1995. 25. P. 229–240.
Bailey V.L., Smith J.L., Bolton H. Fungal-to-bacterial biomass ratios in soils investigated for enhanced carbon sequestration // Soil Biol. Biochem. 2002. V. 34. P. 997–1007.
Bastida F., Monero J.L.A., Hernandez T., Garcia C. Past, present and future of soil quality indices: a biological perspective // Geoderma. 2008. V. 147. P. 159–171.
Blagodatskaya E.V., Anderson T.-H. Interactive effects of pH and substrate quality on the fungal-bacterial ration and qCO2 of microbial communities in forest soil // Soil Biol. Biochem. 1998. V. 30. P. 1269–1274.
Chauvat M., Zaitsev A.S., Wolters V. Successional changes of Collembola and soil microbiota during forest rotation // Oecologia. 2003. V. 137. P. 269–276.
Joergensen R.G., Wichern F. Quantitative assessment of the fungal contribution to microbial tissue in soil // Soil Biol. Biochem. 2008. V. 40. № 3. P. 2977–2991.
Pietikainen J., Fritze H. Clear-cutting and prescribed burning in coniferous forest: comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification // Soil Biol. Biochem. 1995. V. 27. P. 101–109.
Thiffault E., Hannam K.D., Quideau S.A., Pare D., Belanger N., Oh S.-W., Munson A.D. Chemical composition of forest floor and consequences for nutrient availability after wildfire and harvesting in the boreal forest // Plant Soil. 2008. V. 308. P. 37–53.
Velvis H. Evaluation of the selective respiratory inhibition method for measuring the ratio of fungal: bacterial activity in acid agricultural soils // Biol. Fertil. Soils. 1997. V. 25. P. 354–360.
Дополнительные материалы отсутствуют.
Инструменты
Почвоведение