

Почвоведение, 2020, № 8, стр. 921-933
Распределение органического вещества и азота в дерново-подбурах Приокско-Террасного заповедника и его связь со структурой лесных фитоценозов
И. В. Припутина a, *, Г. Г. Фролова a, В. Н. Шанин a, b, Т. Н. Мякшина a, П. Я. Грабарник a
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, 2, корп. 2, Россия
b Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32, Россия
* E-mail: priputina@pbcras.ru
Поступила в редакцию 26.11.2019
После доработки 05.02.2020
Принята к публикации 27.02.2020
Аннотация
Анализ пространственной неоднородности распределения органического вещества в лесных почвах, связанной с влиянием видовой и пространственной структуры фитоценозов, актуален для многих экологических задач, включая оценки стока углерода и моделирование динамики лесных экосистем. На примере дерново-подбуров (Entic Podzols (Arenic)) Приокско-Террасного заповедника (54.89° N, 37.56° E) проанализированы данные о содержании углерода (Сорг) и азота (Nобщ) в лесной подстилке (О) и серогумусовом (AY) горизонте почв под хвойно-широколиственным древостоем, сформировавшемся после зарастания вырубки на месте сосняков. Пробы отбирали вблизи стволов, под кроной и в межкроновом пространстве по трансектам между деревьями разных видов. Содержание Сорг и Nобщ в горизонте О варьировало в диапазонах 17.6–44.9 и 0.84–1.79%; в горизонте AY – 0.71–8.5 (Сорг) и 0.035–0.33% (Nобщ). Более высокое варьирование показателей характерно для горизонта AY. Получена близкая к линейной зависимость между содержанием Сорг и Nобщ в горизонтах О и AY (коэффициенты корреляции Спирмена 0.72 и 0.96 соответственно). Мощность горизонта О и содержание в нем Nобщ, а также содержание Сорг в горизонте AY значимо (P < 0.05) различаются на разных участках трансект. Также значимо различаются мощность и содержание Сорг в горизонте О под кронами разных видов деревьев. В пробах горизонта AY, отобранных в межкроновых пространствах, прослеживается зависимость между содержанием Nобщ и требовательностью видов напочвенного покрова к трофности почв. Полученные данные отражают влияние особенностей локализации поверхностного и внутрипочвенного опада многовидовых древостоев на пространственную неоднородность содержания Сорг и Nобщ в почвенном покрове хвойно-широколиственных лесов.
ВВЕДЕНИЕ
Пространственная неоднородность почвенных условий и связанная с этим вариабельность физико-химических и биологических показателей почв присущи всем иерархическим уровням организации почвенного покрова и ярко проявляются в лесных экосистемах [10, 11, 17, 35, 45]. В частности, различия эдафических условий внутри биогеоценозов (БГЦ), благоприятные для разных видов растительности и почвенной биоты, обеспечивают высокое биоразнообразие лесов [21, 30, 53, 54].
В свою очередь, структура лесных фитоценозов – видовой состав древостоя и напочвенного покрова, взаимное расположение и размеры деревьев – во многом влияет на пространственное распределение почвенных запасов органического вещества и элементов питания, щелочно-кислотные условия и др. Это объясняется определенной локализацией в пространстве БГЦ поступающего в почву видоспецифичного напочвенного и внутрипочвенного (корневого) опада, а также трансформацией химического состава атмосферных осадков при их фильтрации сквозь кроны и по стволам деревьев [2, 14, 22, 24, 41, 52].
Не менее значимо формирование в почвах под пологом лесной растительности разных гидротермических условий [12, 40, 44], от которых зависят активность почвенной биоты и связанные с этим процессы деструкции и минерализации растительных остатков и органического вещества почв. Как следствие, пространственное варьирование массы поступающего растительного опада и внутрипочвенных процессов углеродного и азотного циклов, находят отражение в распределении почвенных запасов органического вещества и азота в пределах лесных БГЦ, для которых характерны определенные закономерности [16, 26, 32, 42, 49]. При этом пространственная локализация фракций корневого опада, связанная с особенностями функционирования и морфологии корневых систем, прослеживается и на микроуровне, что отражают различия в показателях между ризосферной и неризосферной частью почвы [54, 60]. Естественно, что связи между пространственной мозаикой лесного фитоценоза и почвенными характеристиками более явно прослеживаются, когда сформирована относительно устойчивая структура видов-эдификаторов древесного яруса и доминантных видов напочвенного покрова.
Вопросы формирования пространственной неоднородности лесных почв в зависимости от структуры фитоценоза затронуты во многих отечественных публикациях. Например, для хвойных, хвойно-мелколиственных и мелколиственных лесов разных природных зон приводятся данные о значимых различиях в запасах подстилки, элементов питания или микробной активности в почвах прикомлевой зоны деревьев, под кронами разных пород и в межкроновых пространствах [2, 3, 16, 17, 23, 24, 26, 29]. В литературе этим относительно легко выделяемым в пространстве лесного БГЦ участкам соответствуют разные термины: (микро)парцелла, тессера, микросайт, микробиотоп и др. Вопросы терминологии детально обсуждаются в работе [17].
Наше внимание к теме вариабельности почвенных показателей в зависимости от приуроченности к участкам произрастания разных видов древесного яруса и доминант напочвенного покрова объясняется необходимостью получения данных для верификации и тестирования почвенной модели Romul_Hum [48] при моделировании динамики лесных экосистем с использованием системы моделей EFIMOD [4]. Как и другие биогеохимические почвенные модели (например, ForSAFE [59] или Forest-DNDC [50]), Romul_Hum представляет собой модель “почвенного профиля”. В подобных моделях описываются процессы и рассчитываются показатели, соответствующие разным генетическим горизонтам, но без учета их пространственной вариабельности.
В то же время моделирование динамики лесных фитоценозов развивается в направлении создания индивидуально-ориентированных пространственно-распределенных моделей древостоев и напочвенного покрова, в которых каждому растению соответствуют определенные координаты в пространстве условной решетки моделирования. Это позволяет рассчитывать, например, зоны корневого питания растений в зависимости от возраста, видоспецифичных размеров и условий произрастания [20, 36, 38, 55, 57]. Также развиваются подходы к моделированию динамики пространственных микрогруппировок растений лесного напочвенного покрова, в зависимости от эдафических условий местообитаний [37].
В этой связи возникает задача моделирования почвенного покрова как совокупности ячеек (клеток), которые в единой системе координат модельного участка связаны “потоками элементов” с индивидуальными растениями (моделями дерева или растений напочвенного покрова) через соответствующие им зоны корневого питания и поступления разных фракций опада. Такой подход согласуется с представлениями о сотовой структуре почвенного покрова лесных БГЦ как пространственного сочетания ячеек разной размерности [10, 26]. Но остается проблема недостатка экспериментальных данных для параметризации и верификации подобных моделей, а также определения необходимой и достаточной детальности пространственного описания почвенных эдафических условий и связей между потоками элементов в системе почва–растение с учетом пространственной неоднородности лесных БГЦ.
В настоящей статье представлены результаты анализа распределения содержания органического вещества (Сорг) и азота (Nобщ) в дерново-подбурах Приокско-Террасного государственного природного биосферного заповедника (ПТЗ), расположенного на юге Московской области в подзоне хвойно-широколиственных лесов. Лесные фитоценозы заповедника имеют разнообразный видовой состав древостоев, подлеска и напочвенного покрова, что связано с их сукцессионной динамикой в условиях заповедного режима последних 70 лет. Сложная пространственная структура БГЦ определяется разнообразием сочетаний почвенного и растительного покрова [7, 25].
Целью исследований был анализ пространственной неоднородности почвенного покрова в многовидовых хвойно-широколиственных лесах по сравнению с лесами, имеющими более простой состав древостоя и напочвенного покрова, для которых в литературе имеется больше информации. Выбор показателей (содержание Сорг и Nобщ) объясняется тем, что в модели Romul_Hum моделируется динамика пулов органического вещества и азота.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводили на одной из постоянных пробных площадей (ПП) Приокско-Террасного заповедника, где с 2016 г. сотрудниками лаборатории моделирования экосистем ИФХиБПП РАН ведется мониторинг условий функционирования и анализ пространственной структуры лесных экосистем [40].
Краткая характеристика почв. Почвенный покров ПП относится к типу дерново-подбуров (Entic Podzols) [13, 47], которые сформировались на песчаных аллювиально-флювиогляциальных отложениях Приокской зандрово-аллювиальной равнины [34]. Мощность подстилочного горизонта О в исследуемых почвах в среднем составляет 3–7 см. Преобладает ферментативный тип подстилки [1]. Залегающий под лесной подстилкой серогумусовый горизонт AY имеет мощность от 7–8 до 12–15 см. Под ним выделяется альфегумусовый горизонт Bf, который на глубине 70–80 см переходит в горизонт ВC и ниже – в горизонт С. По нашим более ранним неопубликованным исследованиям, гранулометрический состав дерново-подбуров ПТЗ слабо меняется в пределах минеральной части профиля: содержание физического песка в горизонте AY – 85–88%, в нижележащих горизонтах – 90–92%. Кислотность (рНКСl) несколько возрастает с глубиной, изменяясь от 5.4 до 4.4. Содержание Сорг в минеральной части почвенного профиля снижается от 1.5–3% в горизонте AY до 0.3–0.5% в горизонте Bf. Для соединений азота характерен аналогичный гумусово-аккумулятивный характер внутрипрофильного распределения. Максимум содержания обменных катионов также приурочен к горизонту AY и составляет около 5 смоль(экв)/кг почвы.
Видовой состав древостоя и напочвенного покрова. ПП заложена в лесном фитоценозе, сформировавшемся после вырубки сосновых древостоев в 1930–1940 гг. Начальное зарастание участка, на котором, по-видимому, были оставлены семенные деревья и тонкомеры, происходило преимущественно березой (Betula spp.), сохранившей численное доминирование в верхнем древесном ярусе при заметном участии сосны (Pínus sylvéstris L.) и ели (Picea abies (L.) H. Karst.). В этом же ярусе встречаются отдельные деревья липы (Tília cordáta Mill.). Второй ярус представлен елью, липой, дубом (Quércus róbur L.) и березой. В подросте преобладают ель, дуб и липа. По сумме площадей сечений формула древостоя первого яруса 4С4Б2Е + Лп, второго – 6Е2Лп1Д1Б; сумма площадей сечений – 27.02 м2/га. Средний возраст сосны, по данным [40], составляет 115 лет (диапазон возрастов от 70 до 170 лет), березы – 100 лет (диапазон – от 45 до 165 лет), ели – 75 (диапазон – от 30 до 100, а отдельных деревьев – до 150 лет). Более детальная характеристика структуры и онтогенетических состояний древостоя дана в работе [40], где также представлены данные анализа пространственного распределения стволов и центроидов проекций крон деревьев верхнего яруса и динамика неоднородности гидротермических условий под пологом леса.
Мозаика напочвенного покрова в пределах ПП образована сочетанием участков разного размера (от 2–3 до 10 м2 и более) с доминированием 1–2 видов или с сильноразреженным мохово-травяно-кустарничковым ярусом. Доминантными видами на момент проведения полевых исследований (середина вегетационного сезона 2018 г.) были черника (Vaccínium myrtíllus L.), брусника (Vaccínium vítis-idaéa L.), ландыш (Convallária majális L.), орляк (Pterídium aquilínum (L.) Kuhn), вейник (Calamagrostis arundinácea Roth), а также зеленые мхи (преимущественно, Pleurozium schreberi (Willd. ex Brid.) Mitt.). Согласно [28], практически все они относятся к видам нескольких экоценотических ниш; по условиям почвенного питания ландыш предпочитает эвтрофные местообитания, черника – преимущественно мезотроф, брусника, орляк и вейник – олигомезотрофы.
Полевые и химико-аналитические исследования. Оценки пространственного распределения органического вещества и азота в исследуемых почвах основаны на сопряженном анализе содержания Сорг и Nобщ в горизонтах О и AY, поскольку в них сосредоточена бóльшая часть суммарного почвенного пула этих элементов. Пробы из горизонтов отбирали по трансектам, заложенным между парами соседних деревьев первого яруса в виде серии из пяти точек опробования. На данном этапе исследований мы ограничились видами первого яруса (береза, сосна, ель), так как литературные данные имеются преимущественно для этих трех лесообразующих видов.
Было опробовано 7 трансект с разным сочетанием видов деревьев (табл. 1). При закладке трансект выбирали пары деревьев, проекции крон которых не пересекались и было явно выраженное межкроновое пространство, а напочвенный покров в пределах всей трансекты представлен одним доминантным видом (максимум – двумя), информация о которых также приведена в табл. 1. Каждая трансекта включала две точки опробования вблизи комля деревьев (КО), две – в средней части проекций крон с небольшим смещением к периферии (ПК), одну – в межкроновом пространстве (МК). Схожие схемы опробования встречаются во многих исследованиях по изучению пространственных аспектов влияния древостоя на условия под пологом леса [3, 9, 16, 26]. В частности, это позволяет косвенно учесть различия в количестве и соотношении фракций растительного опада [2, 15] или гидротермических условий в почвенных горизонтах [40]. Опробование проводили на трансектах без явных признаков ветровальных комплексов или крупного валежа. В остальном, их расположение в пространстве ПП носит случайный характер.
Таблица 1.
Геоботаническая характеристика трансект и их протяженность
| Номер трансекты* | Расстояние между стволами деревьев, м | Виды | |
|---|---|---|---|
| деревьев | доминанты напочвенного покрова | ||
| 35 | 8.7 | P. ábies–B. spp. | C. arudinácea Roth |
| 57 | 5.7 | P. ábies–B. spp. | V. myrtíllus L., P. aquilínum (L.) Kuhn |
| 79 | 4.6 | P. ábies–B. spp. | V. vítis-idaéa L., P. schreberi |
| 75 | 9.5 | P. ábies–P. ábies | V. myrtíllus L. |
| 91 | 6.0 | P. ábies–P. sylvéstris | C. majális L. |
| 99 | 5.9 | P. sylvéstris–P. sylvéstris | V. myrtíllus L. |
| 95 | 9.2 | P. sylvéstris–B. spp. | P. aquilínum (L.) Kuhn |
* Номера трансект соответствуют номерам квадратов 20 × 20 м, на которые разбита территория ПП. Цифры в номерах квадратов означают расстояние (в десятках метров) от северо-западного угла ПП, который принят за нулевую отметку (первая цифра – направление с запада на восток, вторая цифра – с севера на юг).
Все пробы отобрали в течение одного дня, чтобы минимизировать влияние изменений предшествующих погодных условий на изучаемые показатели на разных участках опробования. Места отбора проб маркировали специальными метками с указанием номеров. При отборе проб из подстилочного горизонта фиксировали его мощность.
Подготовку проб для определения в них Сорг и Nобщ проводили по стандартной методике (ГОСТ 17.4.4.02-84). Количественные определения выполняли методом газовой хроматографии на анализаторе EA-1100 (Carlo Erba) в соответствии с методикой исследований МИ № 88-17641-004-2016 (ФР.1.31.2016.23502) в ЦКП “Хроматография” на базе аккредитованной экоаналитической лаборатории Института биологии ФИЦ Коми НЦ УрО РАН.
Статистическую обработку данных проводили в среде статистического программирования R [55]. Значения переменных Сорг или Nобщ в каждом из горизонтов (О или AY) рассматривали как сопряженные выборки. Еще одна выборка включала данные о мощности горизонта О, которую также считали связанной с другими, что не противоречит представлениям о процессах биохимической трансформации лесной подстилки и формирования гумусового профиля почв [19]. Для всех выборок рассчитывали показатели описательной статистики, выполняли проверку распределений на нормальность (использовали критерий Шапиро–Уилка), рассчитывали ранговые коэффициенты корреляции Спирмена (rs). Анализ различий в распределении значений внутри выборок проводили относительно факторов: 1 – местоположение точки опробования в пределах трансекты (КО, ПК, МК); 2 – вид дерева (сосна, ель, береза). В случае фактора 2 анализировали подвыборки, включающие значения для точек опробования в КО и ПК зонах. Размеры выборок и подвыборок указаны в таблицах или в подписях к рисункам. Для оценки статистической значимости различий по каждому из факторов использовали критерий Уэлча (в случае нормального распределения параметра) или критерий Вилкоксона (в случае распределения, отличного от нормального). Уровень значимости принят равным 5% (т.е. различия считались значимыми при P < 0.05).
РЕЗУЛЬТАТЫ
Содержание Сорг в подстилочном горизонте изменяется от 17.6 до 44.9%, составляя в среднем 34–35% (рис. 1); коэффициент вариации (СV) – 20% (табл. 2). Минимальное значение определено для МК трансекты 57, заложенной между елью и березой с преобладанием брусники и зеленых мхов в напочвенном покрове (рис. S1 ). Максимальное содержание Сорг – на той же трансекте 57, но под кроной ели, где напочвенный покров сильно разрежен, а подстилочный горизонт сформирован преимущественно еловой хвоей. Статистическое распределение исследуемой выборки значений соответствует нормальному, но характеризуется правосторонней асимметрией с преобладанием точек опробования, для которых Cорг > 30%.
Рис. 1.
Содержание Сорг и Nобщ в пробах из подстилочного (А) и серогумусового (Б) горизонтов дерново-подбуров ПТЗ; точки на графиках – измеренные значения, усы – ошибка метода.
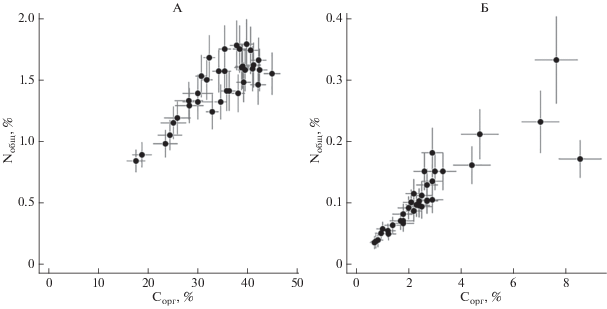
Таблица 2.
Описательные выборочные статистики распределения содержания органического вещества (Сорг) и азота (Nобщ) в подстилочном (О) и серогумусовом (AY) горизонтах дерново-подбуров ПТЗ (размер всех выборок n = 35)
| Горизонт | Описательная статистика | |||||||
|---|---|---|---|---|---|---|---|---|
| минимум | 1-й квартиль | медиана | среднее | 3-й квартиль | максимум | σ, % | СV, % | |
| Сорг, % | ||||||||
| О | 17.60 | 30.00 | 35.40 | 34.18 | 39.35 | 44.90 | 6.9 | 20 |
| AY | 0.71 | 1.75 | 2.40 | 2.68 | 2.90 | 8.50 | 1.8 | 68 |
| Nобщ, % | ||||||||
| О | 0.84 | 1.32 | 1.50 | 1.45 | 1.61 | 1.79 | 0.2 | 17 |
| AY | 0.035 | 0.068 | 0.10 | 0.11 | 0.14 | 0.33 | 0.1 | 56 |
Содержание Сорг в серогумусовом горизонте закономерно ниже, чем в подстилочном, но с бóльшим варьированием: CV = 68%. Максимальное содержание (8.5%) пространственно совпадает с максимумом Сорг в горизонте О под кроной ели трансекты 57 (рис. S2 ). Минимальное значение Сорг (0.71%) определено в МК трансекты 99, заложенной между двух сосен и с редкими кустиками черники в напочвенном покрове. Статистическое распределение величин содержания Сорг в горизонте AY отличается от нормального; суммарно в выборке преобладают значения Cорг < 3%.
Содержание Nобщ в подстилочном горизонте варьирует в диапазоне 0.84–1.79% при средних значениях 1.45–1.5% и CV = 17%. Как и в случае Сорг, минимальное значение получено для МК трансекты 57 (рис. S3 ). Максимальное – в МК трансекты 91, заложенной между елью и сосной с доминированием ландыша в напочвенном покрове. Статистическое распределение значений Nобщ в горизонте О суммарной выборки отличается от нормального.
Содержание Nобщ в серогумусовом горизонте заметно меньше, чем в подстилочном горизонте, и в среднем составляет 0.1% (CV = 56%). Минимальное значение (0.035%) определено в МК трасекты 99, где одновременно отмечен минимум Сорг (рис. S4 ). Максимальное (0.33%) – в КО под елью трансекты 91. Статистическое распределение величин содержания Nобщ в горизонте AY для суммарной выборки отличается от нормального.
Сопряженный анализ данных о содержании Сорг и Nобщ показывает наличие близкой к линейной зависимости между двумя показателями в каждом из горизонтов. Ранговый коэффициент корреляции Спирмена для горизонта О равен 0.72, для AY – 0.96. Следует отметить отсутствие корреляции в содержании Сорг между горизонтами (rs = = −0.037), то же самое для Nобщ (rs = −0.026). Одновременно, результаты анализа свидетельствуют о корреляции между показателями мощности подстилки и содержанием в ней Сорг (rs = 0.56), но для Nобщ эта связь слабее (rs = 0.40). Для горизонта AY связи между содержанием в нем Сорг или Nобщ и мощностью подстилочного горизонта выражены слабо и имеют обратную зависимость (коэффициенты корреляции −0.37 и −0.33).
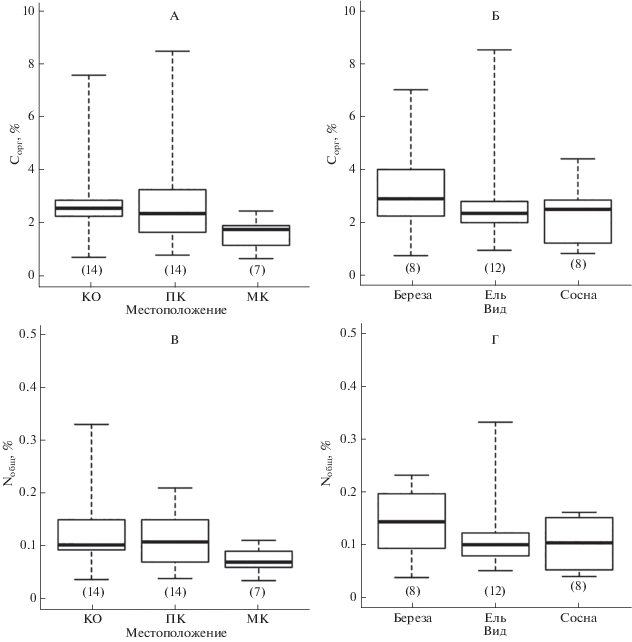
Влияние древесного яруса на мощность лесной подстилки и содержание Сорг и Nобщ в горизонтах О и AY анализировали, исходя из положения мест отбора и вида деревьев. Пространственный аспект наиболее явно выражен в изменении мощности горизонта О (рис. 2), для которого характерно уменьшение максимальных и средних показателей в ряду КО > ПК > МК. Статистически значимые различия по фактору местоположения (P = 0.030) определены для мощности горизонта О между КО и МК, но по содержанию Сорг различия не значимы. Одновременно в подстилочном горизонте значимо различается содержание Nобщ между КО и ПК (P = 0.032), с более высокими средними значениями в КО. В горизонте AY (рис. 3), напротив, значимо различается содержание Сорг между КО и МК (P = 0.018), а для содержания Nобщ статистически значимых различий между разными местоположениями нет.
Рис. 2.
Распределение значений мощности подстилочного горизонта О (А, Б), содержания в нем Сорг (В, Г) и Nобщ (Д, Е) в зависимости от мест отбора проб (КО – вблизи комля, ПК – под кроной, МК – в межкроновом пространстве) и вида деревьев; на графиках отражены медиана, первый и третий квартили, минимальное и максимальное значения соответствующих выборок.
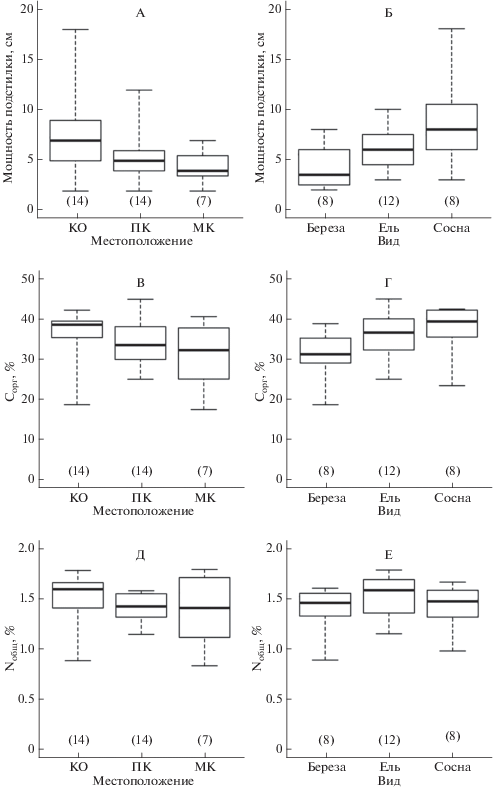
Рис. 3.
Распределение значений содержания Сорг (А, Б) и Nобщ (В, Г) в серогумусовом горизонте AY в зависимости от мест отбора проб (КО – вблизи комля, ПК – под кроной, МК – в межкроновом пространстве) и вида деревьев; на графиках отражены медиана, 1- и 3-й квартили, минимальное и максимальное значения соответствующих выборок.
Влияние видов деревьев на содержание Сорг и Nобщ наиболее заметно прослеживается в подстилочном горизонте (рис. 2). Значимые различия получены между березой и сосной для мощности подстилки (P = 0.032) и содержания в ней Сорг (P = = 0.027). В этой же подвыборке значений показано более высокое содержание Nобщ в горизонте О для ели, хотя значимые различия между видами отсутствуют.
При рассмотрении совместного влияния двух факторов (табл. 3) отмечается повышенное содержание Сорг в подстилке под кронами хвойных видов с более высокими средними показателями у сосны (P. sylvéstris > P. ábies > B. spp.). Содержание Nобщ в горизонте О больше под кронами березы (B. spp. > P. sylvéstris > P. ábies). Для прикомлевых участков получена другая зависимость распределения средних значений Nобщ в подстилочном горизонте (P. ábies > P. sylvéstris > B. spp.). В пробах горизонта AY, отобранных вблизи комля, содержание Сорг и Nобщ в среднем выше для ели и сосны (P. ábies ≥ P. sylvéstris > B. spp.), но под кронами в этом горизонте повышенные средние значения содержания Nобщ получены для березы, а минимальные – для сосны (B. spp. ≥ P. ábies > P. sylvéstris).
Таблица 3.
Содержание Сорг и Nобщ в подстилочном (О) и серогумусовом (AY) горизонтах под разными видами деревьев в зависимости от положения точки отбора (над чертой – среднее арифметическое, в скобках – минимальное и максимальное; КО – у комля, ПК – под кроной; n – размер выборки для соответствующего почвенного горизонта)
| Вид | Горизонт | Cорг, % | Nобщ, % | n | ||
|---|---|---|---|---|---|---|
| КО | ПК | КО | ПК | |||
| Betula spp. | O | $\frac{{30.98}}{{(18.80{\kern 1pt} - {\kern 1pt} 38.80)}}$ | $\frac{{31.23}}{{\left( {28.20{\kern 1pt} --{\kern 1pt} 34.20} \right)}}$ | $\frac{{1.31}}{{\left( {0.89{\kern 1pt} --{\kern 1pt} 1.60} \right)}}$ | $\frac{{1.48}}{{\left( {1.33--1.57} \right)}}$ | 4 |
| AY | $\frac{{3.29}}{{\left( {0.76{\kern 1pt} --{\kern 1pt} 7.00} \right)}}$ | $\frac{{3.23}}{{\left( {2.00{\kern 1pt} --{\kern 1pt} 4.70} \right)}}$ | $\frac{{0.14}}{{\left( {0.04{\kern 1pt} --{\kern 1pt} 0.23} \right)}}$ | $\frac{{0.15}}{{\left( {0.09{\kern 1pt} --{\kern 1pt} 0.21} \right)}}$ | 4 | |
| Picea abies (L.) H. Karst. | O | $\frac{{38.80}}{{\left( {35.40{\kern 1pt} --{\kern 1pt} 41.20} \right)}}$ | $\frac{{33.05}}{{\left( {25.10{\kern 1pt} --{\kern 1pt} 44.90} \right)}}$ | $\frac{{1.68}}{{\left( {1.59{\kern 1pt} --{\kern 1pt} 1.70} \right)}}$ | $\frac{{1.38}}{{\left( {1.15{\kern 1pt} --{\kern 1pt} 1.57} \right)}}$ | 6 |
| AY | $\frac{{3.28}}{{\left( {1.80{\kern 1pt} --{\kern 1pt} 7.60} \right)}}$ | $\frac{{3.04}}{{\left( {0.96{\kern 1pt} --{\kern 1pt} 8.50} \right)}}$ | $\frac{{0.14}}{{\left( {0.07{\kern 1pt} --{\kern 1pt} 0.33} \right)}}$ | $\frac{{0.10}}{{\left( {0.05{\kern 1pt} --{\kern 1pt} 0.17} \right)}}$ | 6 | |
| Pínus sylvéstris L. | O | $\frac{{36.10}}{{\left( {23.50{\kern 1pt} --{\kern 1pt} 42.20} \right)}}$ | $\frac{{38.88}}{{\left( {32.90{\kern 1pt} --{\kern 1pt} 42.40} \right)}}$ | $\frac{{1.43}}{{\left( {0.98{\kern 1pt} --{\kern 1pt} 1.66} \right)}}$ | $\frac{{1.42}}{{\left( {1.24{\kern 1pt} --{\kern 1pt} 1.58} \right)}}$ | 4 |
| AY | $\frac{{2.33}}{{\left( {1.22{\kern 1pt} --{\kern 1pt} 3.00} \right)}}$ | $\frac{{2.27}}{{\left( {0.84{\kern 1pt} --{\kern 1pt} 4.40} \right)}}$ | $\frac{{0.10}}{{\left( {0.05{\kern 1pt} --{\kern 1pt} 0.15} \right)}}$ | $\frac{{0.10}}{{\left( {0.04{\kern 1pt} --{\kern 1pt} 0.16} \right)}}$ | 4 | |
Влияние доминантных видов напочвенного покрова на распределение исследуемых почвенных показателей анализировали по данным межкроновых участков, предполагая для них бóльший вклад растений напочвенного покрова в суммарный пул ежегодного опада по сравнению с ПК и КО. Содержание Сорг в горизонте О изменяется от 17.6 до 40.6%, что практически совпадает с диапазоном значений этого показателя в суммарной выборке, а варьирование содержания Nобщ от 0.84 до 1.79% полностью соответствует диапазону значений “минимальное–максимальное” для исследуемой ПП (табл. 2). В горизонте AY, напротив, варьирование значений минимально по сравнению с аналогичными данными для КО и ПК. Следует отметить, что максимальные значения Nобщ в обоих горизонтах получены для межкронового участка трансекты 91, где в напочвенном покрове доминирует ландыш – вид преимущественно эвтрофных местообитаний.
ОБСУЖДЕНИЕ
Пространственное распределение почвенных свойств как отражение структуры лесных фитоценозов. Принимая во внимание данные публикаций других авторов [2, 16, 17, 23, 24], мы ожидали получить для дерново-подбуров ПТЗ более явные связи между содержанием Сорг и Nобщ в почвенных горизонтах и пространственно-видовой структурой древесного полога. Результаты статистического анализа позволили выявить некоторые тенденции пространственной дифференциации содержания Сорг и Nобщ, которые отражаются в соотношении средних значений параметров в соответствующих выборках. Относительно небольшой размер выборок и высокая вариабельность анализируемых показателей не исключают гипотетического характера наших интерпретаций полученных результатов.
Данные о статистически значимом уменьшении мощности подстилочного горизонта от прикомлевых к межкроновым участкам (КО > ПК > МК) хорошо согласуются с данными других авторов [2, 3, 26]. Вариабельность мощности горизонта О отражает различия в количестве опада, поступающего на поверхность почвы, и разные условия его деструкции и минерализации под кронами деревьев и в межкроновых пространствах. Схожий характер изменений с бóльшей или меньшей значимостью различий повторяется в распределении по трансектам содержания Сорг и Nобщ, что может рассматриваться как подтверждение ведущей роли древостоя в поступлении растительного опада и формировании органического вещества почв в исследуемом БГЦ.
В межкроновых участках заметное влияние на почвенные показатели может оказывать видоспецифичность опада травяно-кустарничкого яруса, что, по-видимому, отражает отмеченный нами широкий диапазон значений содержания Сорг и Nобщ в горизонте О соответствующих местоположений. Хотя нельзя исключать, что в исследуемом фитоценозе вариабельность этих данных может быть связана и с различным соотношением опада хвои и листвы разных видов деревьев (в том числе широколиственных), растущих вблизи условной линии трансект, что требует дополнительного исследования.
С видами деревьев связь пространственного распределения содержания Сорг и Nобщ в дерново-подбурах ПТЗ слабее, чем с положением мест отбора проб, и она по-разному проявляется для подстилочного и серогумусового горизонтов. Характерное для горизонта О уменьшение содержания Сорг и мощности подстилки в ряду P. sylvéstris > > P. ábies > B. spp. согласуется с данными других авторов для этих видов [18, 26]. Но в нижележащем горизонте AY для Сорг наблюдается обратное распределение B. spp. > P. ábies > P. sylvéstris. Схожая инверсия между горизонтами характерна и для содержания Nобщ: в подстилочном горизонте максимальные средние значения определены под елью, в серогумусовом – под березой. Однако, учитывая разное содержание азота в хвое сосны и ели и в листьях березы [5], отмеченные различия между горизонтами не противоречат имеющимся представлениям и эмпирическим данным о процессах деструкции, гумификации и минерализации опада с разным соотношением С : N и перераспределения образующихся продуктов гумификации по профилю почв [19, 31, 33].
Возвращаясь к теме математических моделей, полагаем, что полученные данные подтверждают логичность пространственного моделирования биогенных циклов элементов в лесных БГЦ через подобие “мозаик” горизонтальной структуры древостоя и почвенного покрова. Но в горизонтальной структуре древостоя, формируемой взаимным расположением разных видов, отдельного рассмотрения и математического описания требует видоспецифичная архитектура корневых систем, на что указывают следующие результаты наших исследований.
Пространственная локализация фракций поверхностного и внутрипочвенного растительного опада в разных типах леса. Получена высокая вариабельность значений содержания Сорг и Nобщ в горизонте AY по сравнению с горизонтом О. Считаем, что эти различия могут быть объяснены разным характером условий формирования и локализации поверхностного и внутрипочвенного опада в многовидовых древостоях. Стволы деревьев разных видов в пространстве исследуемой ПП размещены нерегулярно [40], что способствует формированию в данном БГЦ поверхностного опада относительно схожего состава. Подобный эффект воздушного переноса опадающей листвы и хвои в БГЦ, показанный в работе [6], снижает пространственную контрастность показателей содержания Сорг и Nобщ в горизонте О, связанную с поступлением под кроны деревьев соответствующего им видоспецифичного поверхностного опада.
В горизонте AY, напротив, пространственная неоднородность поступления фракций опада больше. Помимо корней растений травяно-кустарничкового яруса (отличающихся разнообразием морфологических типов и соответствующего им пространственного расположения), в этом горизонте сосредоточены многочисленные тонкие корни деревьев. Повышенное содержание азота и высокая оборачиваемость фитомассы тонких корней сочетаются с их определенной локализацией в пространстве – с минимумом вблизи комля и увеличением их количества и суммарной массы к зоне, соответствующей периферии крон деревьев [27, 46, 51, 58]. Дополнительным фактором пространственного распределения корней в почвенных горизонтах служит корневая конкуренция между соседними растениями [43, 57]. Все это создает сложное внутрипочвенное распределение различных фракций корневого опада. Естественно, что формирование органического вещества горизонта AY тесно связано с перемещением растительных остатков и миграцией органических соединений из подстилочного горизонта [32]. Как следствие, пространственная неоднородность горизонта AY, связанная с локализацией фракций корневого опада, в изучаемых дерново-подбурах “сглаживается” поступлением на поверхность почвы относительно однородного по своим биохимическим характеристикам хвойно-листового опада и внутрипрофильной миграцией продуктов его трансформации. Но это предположение требует дополнительного анализа и подтверждения.
Следует подчеркнуть, что предлагаемая интерпретация данных, по нашему мнению, отражает ситуацию для лесов с многовидовым составом древостоев при отсутствии группового размещения деревьев определенного вида. В лесах, где древостой представлен одним или двумя видами, пространственная неоднородность характеристик подстилочного горизонта может быть более выраженной, например, за счет мозаики напочвенного покрова [26, 29]. Или наоборот – в условиях относительно разреженного древостоя и при однородном напочвенном покрове (например, в зеленомошных или лишайниковых типах леса в северной тайге), видоспецифичный опад, аккумулирующийся под кронами отдельных деревьев (или групп деревьев одного вида), формирует в пространстве БГЦ контрастность почвенных характеристик лесной подстилки и верхних горизонтов почв [22].
Учет пространственной неоднородности почвенного покрова в лесных фитоценозах разного видового состава имеет важное значение при анализе масс-баланса C и N и связанных с этим оценок депонирования элементов в лесных почвах [26]. Для лесов России в подобных оценках [39] используются результаты изучения почвенных разрезов, которые обычно закладываются в межкроновых пространствах, реже под кронами деревьев. Полученные нами данные для дерново-подбуров ПТЗ показывают, что в многовидовых сообществах хвойно-широколиственных лесов почвенный покров межкроновых участков может отличаться высокой пространственной вариабельностью показателей. И что более важно, запасы углерода и азота в поверхностных горизонтах почв межкроновых участков в среднем ниже, чем под кронами деревьев. С учетом соотношения подкроновых и межкроновых участков в пространстве лесных БГЦ, недооценка почвенного стока биогенных элементов может быть весьма существенной.
ЗАКЛЮЧЕНИЕ
Приведенные результаты исследований отражают частный случай пространственного распределения содержания Сорг и Nобщ в почвах конкретной лесной территории. Более того, полученные данные являются одномоментным “срезом” почвенно-биогеохимических условий, которые, как известно, определяются сложным (и часто случайным) сочетанием разнонаправленных процессов, характерных для лесных почв и фитоценозов. Тем не менее, анализ полученных данных позволяет обратить внимание на пространственные особенности почвенных циклов С и N в разных типах леса, которые ранее в литературе специально не акцентировались. Мы имеем в виду различия в пространственной локализации и соотношении фракций поверхностного и корневого опада в лесах разного видового состава и разных природно-климатических зон, которые определяют пространственную вариабельность почвенных характеристик.
В исследованных дерново-подбурах ПТЗ эти различия проявляются в сочетании относительно невысокой пространственной вариабельности содержания Сорг и Nобщ в горизонте лесной подстилки (формируемой поверхностным опадом с преобладанием в его составе хвои и листвы многих видов деревьев) и значительного варьирования показателей в нижележащем серогумусовом горизонте (для которого характерно сложное распределение корневых систем древостоя и растений травяно-кустарничкового яруса). На этом фоне выделяются межкроновые участки, где при относительно небольшой мощности подстилочного горизонта наблюдается широкий диапазон значений содержания в нем Сорг и Nобщ, а в серогумусовом горизонте, напротив, минимальный разброс значений тех же показателей.
Список литературы
Бирюков М.В., Рыжова И.М., Гунина А.А., Богатырев Л.Г., Погожева Е.А. Оценка уровня стабилизации органического вещества в условиях почвенных лизиметров // Вестник Моск. ун-та. Сер. 17, почвоведение. 2014. № 2. С. 9–15. https://doi.org/10.3103/S0147687414020021
Ведрова Э.Ф. Модельные опыты по выявлению роли сосны в формировании почвенной пестроты в лесу // Исследование и моделирование почвообразования в лесных биогеоценозах. Новосибирск: Наука, 1979. С. 30–42.
Воробейчик Е.Л., Пищулин П.Г. Промышленное загрязнение и формирование структуры полей концентраций тяжелых металлов в лесной подстилке // Экология. 2016. № 3. С. 323–334.
Грабарник П.Я., Шанин В.Н., Чертов О.Г., Припутина И.В., Быховец С.С., Петропавловский Б.С., Фролов П.В., Зубкова Е.В., Шашков М.П., Фролова Г.Г. Моделирование динамики лесных экосистем как инструмент прогнозирования и управления лесами // Лесоведение. 2019. № 6. С. 488–500.
Дымов А.А., Бобкова К.С., Тужилкина В.В., Ракина Д.А. Растительный опад в коренном ельнике и лиственно-хвойных насаждениях // Известия ВУЗов. Лесной журнал. 2012. № 3. С. 7–18.
Земсков Ф.И., Галкин В.С., Анохина Н.А., Богатырев Л.Г., Демидова А.Н., Прилепский Н.Г., Жилин Н.И., Бенедиктова А.И. Методические особенности исследования динамики поступления опада в условиях стационарных почвенных лизиметров // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 1. С. 9–15. https://doi.org/10.3103/S0147687417010082
Иванов И.В., Шадриков И.Г., Асаинова Ж.С., Дмитраков Л.М. Пространственно-временные аспекты соотношения почвенного и растительного покрова на границе южной тайги и смешанных лесов в условиях антропогенного воздействия // Почвенные процессы и пространственно-временная организация почв. М.: Наука, 2006. С. 78–97.
Иванова Е.А., Лукина Н.В. Варьирование массы и фракционного состава дрвесного опада в сосняках кустарничково-лишайниковых при аэротехногенном загрязнении // Лесоведение. 2017. № 5. С. 47–58.
Ипатов В.С., Кирикова Л.А. К характеристике фитогенного поля Picea abies (Pinaceae) в зеленомошных сосняках // Ботанический журн. 2001. Т. 86. № 5. С. 94–103.
Карпачевский Л.О. Лес и лесные почвы. М.: Лесная промышленность, 1981. 262 с.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Моск. ун-та, 1977. 312 с.
Карпачевский Л.О., Зубкова Т.А., Ташнинова Л.Н., Руденко Р.Н. Почвенный покров и парцеллярная структура лесного биогеоценоза // Лесоведение. 2007. № 6. С. 107–113.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Кузнецов М.А. Влияние условий разложения и состава опада на характеристики и запас подстилки в среднетаежном чернично-сфагновом ельнике // Лесоведение. 2010. № 6. С. 54–60.
Лебедева В.Х., Тиходеева М.Ю., Ипатов В.С. Влияние древесного полога на виды напочвенного покрова в ельнике чернично-зеленомошном // Ботанический журнал. 2005. Т. 90. № 3. С. 400–410.
Лукина Н.В., Орлова М.А. Питательный режим почв старовозрастных лесов Кольского полуострова // Лесоведение. 2008. № 1. С. 11–22.
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Мамай А.В., Федорец Н.Г., Степанов А.Л. Процессы азотфиксации и денитрификации в подзолистых почвах хвойных и мелколиственных лесов таежной подзоны Карелии // Лесоведение. 2013. № 1.C. 66–74.
Милановский Е.Ю., Шеин Е.В. Функциональная роль амфифильных компонентов гумусовых веществ в процессах гумусо-структурообразования и в генезисе почв // Почвоведение. 2002. № 10. С. 1201–1213.
Моделирование динамики органического вещества в лесных экосистемах / Отв. ред. Кудеяров В.Н. М.: Наука, 2007. 380 с.
Мониторинг биоразнообразия лесов: методология и методы / Отв. ред. Исаев А.С. М.: Наука, 2008. 453 с.
Орлова М.А., Лукина Н.В., Камаев И.О., Смирнов В.Э., Кравченко Т.В. Мозаичность лесных биогеоценозов и плодородие почв // Лесоведение. 2011. № 6. С. 39–48.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. Влияние ели на кислотность и содержание элементов питания в почвах северотаежных ельников кустарничково-зеленомошных // Почвоведение. 2016. № 11. С. 1355–1367. https://doi.org/10.1134/S1064229316110077
Орлова М.А., Лукина Н.В., Смирнов В.Э., Краснов Д.А., Камаев И.О. Плодородие еловых лесов Хибинских гор // Почвоведение. 2012. № 6. С. 682–694. https://doi.org/10.1134/S1064229312060087
Оценка и сохранение биоразнообразия лесного покрова в заповедниках европейской России. Гл. 4. Приокско-Террасный заповедник. М.: Научный мир, 2000. С. 73–103.
Подвезенная М.А., Рыжова И.М. Зависимость вариабельности запасов углерода в почве от пространственной структуры растительного покрова лесных биогеоценозов // Вестник Моск. ун-та. Сер. 17, почвоведение. 2010. № 4. С. 3–9. https://doi.org/10.3103/S0147687410040010
Разгулин С.М., Воронин Л.В. Роль тонких корней деревьев в бореальных лесах // Лесоведение. 2018. № 3. С. 225–235. https://doi.org/10.7868/S00241148030051
Разумовский С.М. Труды по экологии и биогеографии (полное собрание сочинений). Товарищество научных изданий. КМК. 2011. 722 с.
Семенюк О.В., Ильяшенко М.А. Пространственная изменчивость почвенных свойств разновозрастных сосняков пейзажной части паркового комплекса “Архангельское” // Вестник Моск. ун-та. Сер. 17, почвоведение. 2013. № 1. С. 23–29.
Смирнова О.В., Алейников А.А., Семиколенных А.А., Бовкунов А.Д., Запрудина М.В., Смирнов Н.С. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике // Лесоведение. 2011. № 6. С. 67–78.
Телеснина В.М., Семенюк О.В., Богатырёв Л.Г. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”) // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 4. С. 11–20. https://doi.org/10.3103/S0147687419040082
Титлянова А.А., Шибарева С.В. Подстилки в лесных и травяных экосистемах. Новосибирск: Изд-во СО РАН, 2012. 137 с.
Трофимов С.Я., Дорофеева Е.И., Тарко А.М., Фокин А.Д., Смолина Г.А., Соколова Т.А., Толпешта И.И., Лузиков А.В. Органическое вещество почв как характеристика функциональной организации почвенной системы // Регуляторная роль почвы в функционировании таежных экосистем. М.: Наука, 2002.
Учватов В.П. Ландшафты Приокско-Террасного биосферного заповедника // Ландшафтно-геохимические основы фонового мониторинга. М.: Наука, 1989. С. 103–117.
Фридланд В.М. Проблемы географии, генезиса и эволюции почвенного покрова. М.: Наука, 1986. 243 с.
Фролов П.В., Зубкова Е.В., Комаров А.С. Клеточно-автоматная модель сообщества двух видов растений разных жизненных форм // Известия РАН. Сер. биологическая. 2015. № 4. С. 341–349.
Ханина Л.Г., Грозовская И.С., Смирнов В.Э. Анализ базы данных по биомассе лесного напочвенного покрова для моделирования его динамики в круговоротных моделях лесных экосистем // Хвойные бореальной зоны. 2013. Т. 31. № 1–2. С. 22–29.
Чертов О.Г., Грабарник П.Я., Шанин В.Н., Быховец С.С., Петропавловский Б.С., Припутина И.В., Фролов П.В., Зубкова Е.В. Динамические модели наземных экосистем для количественной оценки продуктивности растительности // Растительные ресурсы. 2019. № 2. С. 151–169.
Честных О.В., Замолодчиков Д.Г., Уткин А.И. Общие запасы биологического углерода и азота в почвах лесного фонда России // Лесоведение. 2004. № 4. С. 30–42.
Шанин В.Н., Шашков М.П., Иванова Н.В., Быховец С.С., Грабарник П.Я. Исследования структуры древостоев и микроклиматических условий под пологом леса на постоянной пробной площади в Приокско-Террасном заповеднике // Труды Приокско-Террасного заповедника. Вып. 7. М., Товарищество научных изданий КМК, 2018. С. 72–85.
Augusto L., Ranger J., Binkley D., Rothe A. Impact of several common tree species of European temperate forests on soil fertility // Annals Forest Sci. 2002. V. 59. P. 233–253. https://doi.org/10.1051/forest:2002020
Bengston B., Falkengren-Grerup U., Bengtsson G. Spatial distributions of plants and gross N transformation rates in a forest soil // J. Ecology. 2006. V. 94(4). P. 754–764. https://doi.org/10.1111/j.1365-2745.2006.01143.x
Bolte A., Villanueva I. Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) // Europ. J. Forest Res. 2006. V. 125. P. 15–26. .https://doi.org/10.1007/s10342-005-0075-5
Bruckner A., Kandeler E., Kampichler C. Plot-scale spatial patterns of soil water content, pH, substrate-induced respiration and N mineralization in a temperate coniferous forest // Geoderma. 1999. V. 93. P. 207–223. https://doi.org/10.1016/S0016-7061(99)00059-2
Gömöryová E., Gregor J., Pichler V., Gömöry D. Spatial patterns of soil microbial characteristics and soil moisture in a natural beech forest // Biologia. 2006. V. 61. Suppl. 19. P. 329–333.
Gordon W.S., Jackson R.B. Nutrient concentrations in fine roots // Ecology. 2000. V. 81(1). P. 275–280.
IUSS Working Group WRB. World Reference Base for Soil Resources 2014. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome.
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing // Ecological Modelling. 2017. V. 345. P. 113–124. https://doi.org/10.1016/j.ecolmodel.2016.08.007
Kooch Y., Bayranvand M. Composition of tree species can mediate spatial variability of C and N cycles in mixed beech forests // Forest Ecology and Management. 2017. V. 401. P. 55–64. https://doi.org/10.1016/j.foreco.2017.07.001
Lamers M., Ingwersen J., Streck T. Modelling N2O emission from a forest upland soil: A procedure for an automatic calibration of the biogeochemical model Forest-DNDC // Ecological Modelling. 2007. V. 205. P. 52–58. https://doi.org/10.1016/j.ecolmodel.2007.02.007
Lawrence D.J., Luckai N., Meyer W.L., Shahi C., Fazekas A.J., Kesanakurti P., Newmasterb S. Distribution of white spruce lateral fine roots as affected by the presence of trembling aspen: root mapping using simple sequence repeat DNA profiling // Can. J. Forest Res. 2012. V. 42(8). P. 1566–1576. https://doi.org/10.1139/x2012-082
Levia D.F., Germer S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands // Rev. Geophys. 2015. V. 53. № 3. P. 673–714.
Levine J.M., HilleRisLambers J. The importance of niches for the maintenance of species diversity // Nature. 2009. V. 461. P. 254–257. https://doi.org/10.1038/nature08251
Martiny J.B., Eisen J.E., Penn K., Allison S.D., Horner-Devine M.C. Drivers of bacterial β-diversity depend on spatial scale // PNAS. 2011. V. 108. P. 7850–7854. https://doi.org/10.1073/pnas.1016308108
Pommerening A., Grabarnik P. Individual-based Methods in Forest Ecology and Management. Springer Nature, 2019. 411 p. https://doi.org/10.1007/978-3-030-24528-3
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2015.
Shanin V., Mäkipää R., Shashkov M., Ivanova N., Shestibratov K., Moskalenko S., Rocheva L., Grabarnik P., Bobkova K., Manov A., Osipov A., Burnasheva E., Bezrukova M. New procedure for the simulation of belowground competition can improve the performance of forest simulation models // Europ. J. Forest Res. 2015. V. 134. P. 1055–1074. https://doi.org/10.1007/s10342-015-0909-8
Strong W.L., La Roi G.H. Root-system morphology of common boreal forest trees in Alberta, Canada // Can. J. Forest Res. 1983. V. 13(6). P. 1164–1173. https://doi.org/10.1139/x83-155
Wallman P., Svensson M., Sverdrup H., Belyazid S. ForSAFE – an integrated process-oriented forest model for long-term sustainability assessments // Forest Ecol. Managem. 2005. V. 207(1–2). P. 19–36. https://doi.org/10.1016/j.foreco.2004.10.016
Yang F., Tian J., Fang H., Gaoa J., Zhang X., Yu G., Kuzyakov Y. Spatial heterogeneity of microbial community and enzyme activities in a broad-leaved Korean pine mixed forest // Europ. J. Soil Biol. 2018. V. 88. P. 65–72. https://doi.org/10.1016/j.ejsobi.2014.12.001
Дополнительные материалы
- скачать EMS.docx
- Рис. S1. - Рис. S4.