

Почвоведение, 2021, № 11, стр. 1392-1406
Влияние типа землепользования на свойства почв и структуру экосистемных запасов углерода в среднетаежной подзоне Карелии
И. А. Дубровина a, *, Е. В. Мошкина b, В. А. Сидорова a, А. В. Туюнен b, А. Ю. Карпечко b, Н. В. Геникова b, М. В. Медведева b, А. В. Мамай b, О. В. Толстогузов c, Л. М. Кулакова c
a Институт биологии КарНЦ РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
b Институт леса КарНЦ РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
c Институт экономики КарНЦ РАН
185030 Петрозаводск, пр. А. Невского, 50, Россия
* E-mail: vorgo@mail.ru
Поступила в редакцию 01.06.2020
После доработки 20.04.2021
Принята к публикации 27.04.2021
Аннотация
Исследовано влияние типа землепользования на свойства легкосуглинистых подзолистых почв нормального увлажнения и экосистемные запасы углерода в среднетаежной подзоне Карелии. Проанализированы изменения в строении почвенных профилей, основные агрохимические и микробиологические показатели верхних горизонтов, запасы Сорг и Смик почвы в метровом слое и структура углеродных пулов участков. Исследовали 110-летний ельник в качестве контроля, пашню, сенокос, а также молодой березняк 20 лет и средневозрастной ельник 65 лет естественного лесовозобновления на бывших сельскохозяйственных угодьях. На пашне почва уплотнена до 1.5 г/см3, имеет величину рН 5.2 и высокое содержание элементов питания. В почвах пашни и сенокоса содержание Сорг находится в пределах 1.6–2.1%, Nобщ 0.2%. В лесных почвах зафиксировано уменьшение плотности до 1.1–0.9 г/см3, величины рН до 4.0–3.2, ухудшение общего агрохимического фона при увеличении содержания Сорг до 2.2–4.6%. Для почв пашни и спелого леса характерно пониженное содержание Смик (50–60 мг С/кг почвы). На участках сенокоса, молодого и средневозрастного лесов содержание Смик составляет около 250 мг С/кг почвы. Запасы Смик в метровом слое почвы максимальны на участке молодого леса и составляют 88.9 г С/м2, для пашни, сенокоса и средневозрастного леса запасы Смик находятся в пределах 52–61 г С/м2, в спелом лесу 23.6 г С/м2, при этом в лесной подстилке накапливается 14.5 г С/м2. Общие экосистемные запасы углерода максимальны на лесных участках и минимальны на сенокосе. Запасы Сорг почвы в метровом слое пашни составляют 138 т С/га, на участках молодого и средневозрастного лесов уменьшаются до 101–91 т С/га, минимальны под спелым лесом и сенокосом и составляют 67–68 т С/га.
ВВЕДЕНИЕ
Почва является неотъемлемым компонентом наземных экосистем, она обуславливает развитие организмов и обеспечивает связь между большим геологическим и малым биологическим круговоротом веществ [47]. Количественно оценить многофункциональность почв позволяет концепция экосистемных услуг, которые обусловлены почвенными функциями [59, 74], охватывающими все аспекты почвенно-экологических, социальных и экономических услуг [51]. Как элемент биосферы почва выступает в качестве фильтра и буфера на пути миграции химических веществ в водоемы, регулирует газовый состав атмосферы. Населяющие почву микроорганизмы активно преобразует верхнюю часть литосферы. Как элемент биогеоценоза, почва прежде всего является средой и местообитанием организмов, аккумулирует биофильные элементы питания, а также обладает функцией продуктивности [53, 66].
Способность почв предоставлять услуги определяется их свойствами и почвенными процессами, огромное влияние на которые оказывает практика использования земли и управления почвенными ресурсами. Известно, что система землепользования является одним из наиболее важных факторов, влияющих на ключевые свойства и экологические функции почв [69]. Поскольку деградация почвенных ресурсов ведет к общему нарушению экологического баланса биогеоценоза и ландшафта в целом, невозможно переоценить важность учета почвенных функций при смене землепользования [55]. Изменение режима землепользования оказывает влияние на растительный покров, количественные и качественные характеристики углеродного пула и состав микробных сообществ почв. Это в свою очередь влияет как на плодородие почв, так и на эмиссию углекислого газа в атмосферу. Основными факторами, влияющими на секвестрацию почвенного углерода при смене землепользования, считаются климатические условия местности, тип почв и растительности, схема обработки и удобрения почв, а также история участка [71, 81]. Считается, что основные потери углерода почвы происходят при переводе лесных угодий, лугов и пастбищ в пашню. Лесовозобновление на бывших сельскохозяйственных землях, напротив, предполагает накопление почвенного углерода [48, 56]. Несмотря на это мнение, многие исследователи сообщают о противоречивых последствиях облесения, когда лесовозобновление имеет недостоверный эффект или приводит к уменьшению запасов углерода почвы [60, 64, 72, 76].
Общие углеродные запасы экосистемы являются важной составляющей биогеохимического цикла углерода в биологическом круговороте веществ. В вопросе влияния смены землепользования на глобальный цикл углерода существует большая неопределенность. Считается, что изменения в землепользовании обеспечивают до 20% общих антропогенных выбросов CO2 и являются важным чистым источником углекислого газа [65]. Региональные оценки углеродного баланса имеют значительные различия, поскольку зависят от ряда факторов природного и антропогенного характера, поэтому исследования по интегральной оценке структуры запасов углерода разных землепользований чрезвычайно актуальны [44, 58].
Углерод микробной биомассы является важной составляющей почвенного органического углерода. Микроорганизмы прямо и косвенно воздействуют на многие ферментативные, энергетические и биогеохимические процессы, происходящие в почве, влияют на разложение органического вещества и круговорот питательных элементов. Содержание микробного компонента служит индикатором трансформации и деградации почв [67, 75]. При смене режима землепользования значительные изменения претерпевают агрохимические и физико-химические свойства почвы, обеспечивающие функцию почвенной продуктивности [22, 39, 57].
Изменение характера землепользования зависит как от географических особенностей, так и экономического положения территории. В России в контексте смены землепользования в последнее десятилетие активно изучались процессы гумусообразования при ослаблении и прекращении активного агротехнического воздействия и выведения пахотных угодий в залежь [5, 17, 34, 39]. Поскольку 2/3 залежных земель расположено в лесной зоне [26], то многочисленные исследования постагрогенных экосистем методом хронорядов проводились в зоне южной тайги [10, 32, 33, 38, 78]. Для подзоны средней тайги аналогичных исследований проведено гораздо меньше [21, 23]. Здесь основным трендом в изучении смены землепользования является выявление закономерностей изменения свойств почв в процессе естественного возобновления лесной растительности после рубки древостоя [8, 25]. В среднетаежной подзоне, являющейся границей активного земледелия, практически не проводилось комплексного анализа влияния разных типов землепользования на почвенные свойства и функции. В связи с глобальным изменением климата и проблемами обеспечения продовольственной безопасности, для развития северных регионов представляется актуальным изучение экологических последствий смены режимов землепользования.
Цель работы – изучение влияния разных типов землепользования на свойства и функции почв, а также структуру экосистемных запасов углерода. На генетически близких почвах среднетаежной подзоны европейской территории России исследованы изменения в строении почвенных профилей и основных агрохимических и микробиологических показателей. Рассчитаны запасы органического углерода и углерода микробной биомассы почвы в метровом слое, а также запасы углерода в подземной и надземной фитомассе, дебрисе и подстилке для каждого типа землепользования.
ОБЪЕКТЫ И МЕТОДЫ
Исследование проводили на Агробиологической станции Карельского научного центра РАН и прилегающих к ней территориях (61°45′ N, 34°20′ E) в пределах г. Петрозаводска (Южная Карелия). Климат региона умеренно-холодный, переходный от морского к континентальному. В южной части Карелии среднегодовая температура воздуха составляет 2°С при 650 мм осадков в год [1]. Естественная растительность относится к среднетаежной подзоне зеленомошных хвойных лесов. Почвообразование идет на четвертичных отложениях последнего оледенения. Рельеф района исследования представляет собой моренную холмистую равнину, почвообразующей породой является суглинистая морена разной степени завалуненности. В почвенном покрове преобладают подзолистые почвы (Retisols) различного уровня антропогенной трансформации и гидроморфизма.
Объектами изучения явились 5 характерных для данной климатической зоны типов землепользования как сельскохозяйственного, так и лесохозяйственного назначения (табл. 1). Участки выбраны на возвышенных элементах рельефа с легкосуглинистыми почвами нормального увлажнения. Агробиологическая станция основана в 1951 г., поэтому участки пашни и сенокоса находятся в длительном сельскохозяйственном использовании. До 1980-х годов сенокосный участок использовали под посевы многолетних трав, позже агротехнические мероприятия и подсев трав прекратили, и участок перешел в категорию старосеяного луга. Молодой и средневозрастный леса представляют собой участки естественного лесовозобновления на бывших сельскохозяйственных угодьях. Спелый лес произрастает на территории, прилегающей к Агробиологической станции, не испытывавшей значительной антропогенной нагрузки [31].
Таблица 1.
Характеристика участков
| Тип землепользования | Растительность | Почва |
|---|---|---|
| Пашня, двупольный севооборот, вспашка с отвалом пласта, внесение минеральных и зеленых удобрений | Картофель, злаково-бобовые травосмеси | Агрозем текстурно-дифференцированный типичный Umbric Retisol (Aric) |
| Сенокос, сенокошение 2 раза в год, агротехнические мероприятия не проводятся | Луг злаково-разнотравный с преобладанием ежи сборной (Dactylis glomerata L.) и манжетки обыкновенной (Alchemilla vulgaris L.) | Агродерново-подзолистая типичная Umbric Retisol (Cutanic) |
| 20-летний лес, молодой древостой, II класс бонитета | Березняк злаково-разнотравный, в напочвенном покрове доминирует вейник лесной (Calamagrostis arundinacea (L.) Roth.) | Агродерново-подзолистая типичная Umbric Retisol (Cutanic) |
| 65-летний лес, средневозрастный древостой, II класс бонитета | Ельник кисличный, в напочвенном покрове преобладают кислица обыкновенная (Oxalis acetosella L.) и щитовник картузианский (Dryopteris carthusiana (Vill.) H.P. Fuchs) | Дерново-подзолистая постагрогенная Albic Retisol (Ochric) |
| 110-летний лес, спелый древостой, II класс бонитета | Ельник чернично-кисличный, в напочвенном покрове преобладают черника обыкновенная (Vaccinium myrtillus L.) и кислица обыкновенная (Oxalis acetosella L.) | Подзолистая типичная Albic Retisol |
Отбор проб и анализ почв. На каждом участке был заложен почвенный разрез, проведено его морфологическое описание, определена классификационная принадлежность почв [45]. В генетических горизонтах была определена плотность сложения (ρ) весовым методом и отобраны образцы в трехкратной повторности для определения обменной кислотности, содержания органического углерода и углерода микробной биомассы. Для расчета запасов углерода в подстилках на участках 65- и 110-летнего леса в шестикратной повторности с каждого участка были отобраны образцы подстилки. Для анализа вариабельности общих агрохимических свойств и микробиологической активности в верхних минеральных горизонтах почв на каждом участке из прикопок были отобраны образцы с глубины 5–15 см из верхнего горизонта. В почвенных образцах определяли обменную кислотность (рНKCl) потенциометрически на иономере АНИОН 4100 “Инфраспак-Аналит”; содержание органического углерода (Сорг) методом высокотемпературного каталитического сжигания на анализаторе ТОС-L CPN “Shimadzu”; содержание общего азота (Nобщ) по Кьельдалю на анализаторе азота “Buchi”; соотношение С/N; содержание подвижного фосфора (P2O5) со спектрофотометрическим окончанием на спектрофотометре UV-1800 “Shimadzu” и калия (К2О) с атомно-эмиссионным окончанием на атомно-абсорбционном спектрофотометре АА-7000 “Shimadzu” по Кирсанову; сумму обменных оснований (S) по Каппену-Гильковицу [40].
Определение дыхательной активности почв и содержания в ней микробной биомассы проводили согласно рекомендациям [9, 20]. Содержание углерода микробной биомассы (Cмик) определено методом субстрат-индуцированного дыхания (СИД), которое оценивали по скорости начального максимального дыхания микроорганизмов после обогащения почвы глюкозой и инкубации в течение 1.5–2 ч при температуре 22°С. Изменение концентрации СО2 регистрировали газоанализатором на основе NDIR-сенсора “SenseAir”.
Величину СИД рассчитывали по формуле:
Углерод микробной биомассы рассчитывали по формуле:
где Cмик – углерод микробной биомассы, мкг С/г почвы; СИД – субстрат-индуцированное дыхание, мкг С–СО2/г почвы в час.Для верхних горизонтов почв определяли параметры экофизиологического статуса микробного сообщества – соотношение Смик/Сорг и микробный метаболический коэффициент QR, который расчитывали как отношение БД/СИД.
Запасы углерода в почве рассчитывали для каждого генетического горизонта, затем суммировали. Запасы углерода в слое определяли по общей формуле:
где Q – запас углерода, т С/га (г/м2); С – содержание углерода, %; h – мощность горизонта, см; ρ – плотность сложения, г/см3.Анализ растительности. Для оценки основных экосистемных пулов углерода на исследуемых участках руководствовались рекомендациями, разработанными для лесных экосистем [63], но также учитывали фитомассу напочвенной растительности. Определяли 5 основных пулов углерода: надземную фитомассу (древостой и напочвенный покров); подземную фитомассу; дебрис – сухостой и валеж; лесную подстилку; углерод почвы в слое 0–100 см. Геоботаническое описание растительности проводили на площадках 20 × 20 м2. Возраст древостоя определяли с использованием бурава Пресслера. Для расчета запасов фитомассы сельхозугодий использовали средние многолетние данные Федеральной службы государственной статистики по урожайности картофеля и луговых трав. Для расчета запасов фитомассы напочвенного покрова на лесных участках использовали данные для аналогичных сообществ среднетаежной подзоны [15, 24]. Запас фитомассы древостоя рассчитывали на основании таксационных измерений по ОСТ 56-69-83 и показателей плотности древесины [30]. Затем данные по общей фитомассе распределялись по отдельным фракциям [15, 16]. Запас углерода в древесных остатках рассчитывали согласно [41]. При пересчете запасов фитомассы в запасы углерода использовали коэффициент 0.5 для древесных фракций и хвои, 0.45 – для листьев и трав [42].
Статистическая обработка. Для анализа различий средних значений в исследованных показателях применяли однофакторный дисперсионный анализ (ANOVA) с апостериорным анализом по критерию Тьюки при уровне значимости p ≤ 0.05. В таблицах и диаграммах приведены средние значения ± ошибка среднего, значимые различия обозначены разными буквами. Статистическую обработку данных проводили в программах Microsoft Excel и PAST Statistics.
РЕЗУЛЬТАТЫ
Морфология профиля. Исследованные почвы сформированы на суглинистой морене и характеризуются сходным гранулометрическим составом, это крупно-пылеватые и пылевато-песчаные легкие суглинки [31], что предполагает их генетическую однородность (рис. 1). На пашне почва диагностирована как агрозем текстурно-дифференцированный типичный (Umbric Retisol (Aric)). Верхняя часть профиля представлена мощным агрогумусовым горизонтом 35 см, подразделяющимся на два подгоризонта по плотности. Далее идет текстурный горизонт, постепенно переходящий в породу. Элювиальный и субэлювиальный горизонты отсутствуют. Почвы под сенокосом и лесом 20 лет имеют принципиально сходное строение и диагностированы как агродерново-подзолистые типичные (Umbric Retisol (Cutanic)). Для них характерен агрогумусовый горизонт, в нижней части которого наблюдается зона деградации BEL небольшой мощности в виде комбинации осветленного бесструктурного материала и более плотных фрагментов с ореховатой структурой. Глубже расположен текстурный горизонт, резко переходящий в породу. Четко выраженный элювиальный горизонт отсутствует. Для почвы под сенокосом характерен агрогумусовый горизонт небольшой мощности до 15 см, в верхней части которого сформирована дернина. Почва под лесом 20 лет, напротив, имеет хорошо развитый гомогенный агрогумусовый горизонт без признаков дифференциации, травянистая дернина и лесная подстилка отсутствуют. Под 65-летним лесом почва по морфологическому строению близка к естественным зональным почвам, имеет отчетливо выраженный элювиальный горизонт небольшой мощности, который в отсутствие субэлювиального переходит в текстурный горизонт, плавно переходящий в породу. Серогумусовый горизонт мощностью 16 см в нижней части осветлен и менее оструктурен. Это позволяет выделить два субгоризонта и является признаком постагрогенной трансформации. Над серогумусовым горизонтом залегает маломощная лесная подстилка в начальной стадии формирования мощностью в среднем 1 см. Почва диагностирована как дерново-подзолистая постагрогенная (Albic Retisol (Ochric)). Под спелым 110-летним лесом развита типичная подзолистая почва (Albic Retisol) с характерной текстурной дифференциацией профиля и отсутствием гумусово-аккумулятивного горизонта. Элювиальный горизонт мощностью 10 см имеет ясный переход к текстурному по плотности и цвету, субэлювиальный горизонт отсутствует. Непосредственно над элювиальным горизонтом залегает хорошо развитая лесная подстилка мощностью в среднем 6 см.
Рис. 1.
Схематическое строение почвенных профилей. Обозначения здесь и далее: 1 – пашня; 2 – сенокос; 3 – 20-летний лес; 4 – 65-летний лес; 5 – 110-летний лес.

Агрофизические и агрохимические показатели почв. Плотность сложения почв значительно изменяется как по глубине, так и в зависимости от участка (рис. 2). Максимальные значения плотности характерны для пашни и достигают 1.5 г/см3 в верхней части агрогумусового горизонта. На участке сенокоса и в лесных почвах плотность верхних горизонтов уменьшается и составляет в среднем 1.0 г/см3. Максимальная плотность почв на всех участках приурочена к текстурным горизонтам и достигает 1.8 г/см3 на пашне, несколько уменьшаясь на участках сенокоса и молодого леса (1.7 г/см3) и в 65- и 110-летнемлесах (1.5–1.6 г/см3). Величина обменной кислотности минимальна на пашне и имеет значения от 5.3 ед. рН в верхних горизонтах до 4.5 в породе. На участке сенокоса показатели рН меньше и составляют 4.4–4.1 ед. рН. В почве пашни и сенокоса рН в профиле равномерно уменьшается с глубиной. На участках под лесной растительностью значения рН в верхних горизонтах (5–15 см) уменьшаются от молодого (рН 4.0) к спелому лесу (рН 3.2). При этом в лесах величина рН максимальна в текстурных горизонтах и достигает значений 4.2–4.7.
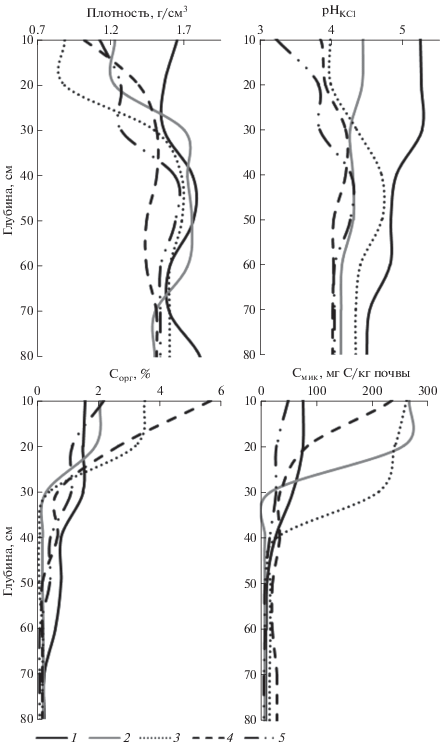
Распределение органического углерода в профиле имеет равномерно-аккумулятивный тип на пашне и регрессивно-аккумулятивный на остальных участках (рис. 2). Содержание углерода в верхних горизонтах (до 20 см) возрастает от почв пашни и сенокоса (1.6–2%) к участкам молодого и средневозрастного лесов (3–5%). Под лесом 110 лет содержание углерода в горизонте EL составляет около 2%, что близко по значению к почвам пашни и сенокоса. На участках сенокоса и леса 20 лет четко выражен элювиальный минимум, когда возросшее содержание углерода в агрогумусовом горизонте резко убывает в горизонте BEL. В почвах 65- и 110-летнего леса кривые распределения углерода более сглаженные за счет несколько повышенного его содержания в горизонтах EL и ВТ. Содержание органического углерода в верхних горизонтах растет в ряду почв от пашни (1.56%) к средневозрастному лесу (4.56%) (табл. 2). Содержание общего азота минимально в почве спелого леса (0.09%) и максимально на участках молодого и средневозрастного лесов (0.26–0.29%). Для почв пашни и сенокоса характерны средние значения показателя – 0.19%. Соотношение С/N узкое в почвах пашни и сенокоса (9.6–12.9), оно несколько возрастает на участках 20- и 65-летнего леса (14.1–18.7). Максимально широкое соотношение зафиксировано в спелом лесу – 27.1.
Таблица 2.
Показатели почвенного плодородия и микробиологической активности в верхних минеральных горизонтах почв (n = 6)
| Показатель | Пашня | Сенокос | Лес | ||
|---|---|---|---|---|---|
| 20 лет | 65 лет | 110 лет | |||
| Горизонт | Р | Р | Р | AY | EL |
| Глубина, см | 5–15 | 5–15 | 5–15 | 5–15 | 6–16 |
| ρ, г/см3 | 1.52 ± 0.04a | 1.17 ± 0.03b | 0.94 ± 0.04c | 0.98 ± 0.03cd | 1.13 ± 0.05bd |
| рНKCl | 5.17 ± 0.06a | 4.32 ± 0.05b | 4.05 ± 0.06c | 3.67 ± 0.08d | 3.23 ± 0.01e |
| P2O5, мг/кг | 306.3 ± 20.3a | 46.8 ± 5.4b | 103.0 ± 4.0c | 14.0 ± 1.2bd | 3.5 ± 0.0d |
| К2О, мг/кг | 59.3 ± 2.2a | 41.0 ± 2.4b | 44.5 ± 4.5ab | 39.8 ± 6.5b | 13.3 ± 0.5c |
| S, смоль(экв)/кг | 6.38 ± 0.65a | 6.00 ± 0.41ab | 4.69 ± 0.68b | 0.64 ± 0.12c | 0.26 ± 0.21c |
| Cорг, % | 1.56 ± 0.05a | 2.07 ± 0.08ab | 3.08 ± 0.18b | 4.56 ± 0.48c | 2.16 ± 0.02ab |
| Nобщ, % | 0.19 ± 0.01a | 0.19 ± 0.01a | 0.26 ± 0.01b | 0.29 ± 0.03b | 0.09 ± 0.01c |
| С/N | 9.6 ± 0.23a | 12.9 ± 0.31b | 14.1 ± 0.27c | 18.7 ± 0.16d | 27.1 ± 0.66e |
| Cмик, мг С/кг | 59.2 ± 5.3a | 258.5 ± 6.7b | 254.1 ± 16.0b | 246.3 ± 18.7b | 49.5 ± 1.3a |
| Cмик/Cорг, % | 0.38 ± 0.03a | 1.26 ± 0.02b | 0.81 ± 0.02c | 0.62± 0.09c | 0.23± 0.01a |
| QR | 0.68 ± 0.04ac | 0.45 ± 0.05b | 0.51 ± 0.09ab | 0.54 ± 0.05ab | 0.86 ± 0.01c |
Содержание доступного фосфора максимально на пашне (306.3 мг/кг) и на участке молодого леса (103 мг/кг). Значительное уменьшение содержания фосфора наблюдается в почве сенокоса (46.8 мг/кг) и средневозрастного леса (14 мг/кг), минимальное количество фосфора в почве отмечено на участке спелого леса – 3.5 мг/кг. Содержание доступного калия также максимально на пашне (59.3 мг/кг), а минимально в 110-летнемлесу (13.3 мг/кг). Для участков сенокоса, 20- и 65-летнего леса различия в содержании калия недостоверны и значения достаточно близки (39.8–44.5 мг/кг). Содержание суммы обменных оснований незначительно падает в ряду почв от пашни (6.38 смоль(экв)/кг) к молодому лесу (4.69 смоль(экв)/кг). Для почв 65- и 110-летнего леса характерны очень низкие значения суммы обменных оснований (0.64–0.26 смоль(экв)/кг).
Микробиологические показатели почв. Углерод микробной биомассы имеет тенденцию изменения, аналогичную изменению содержания общего углерода во всех изученных почвах кроме почвы под сенокосом (рис. 2). Самое низкое содержание Cмик в верхних горизонтах почв зафиксировано на пашне (59.2 мг С/кг) и в спелом лесу (49.5 мг С/кг). Для участков сенокоса, 20- и 65-летнего леса значения показателя Смик близки, составляют 246.3–258.5 мг С/кг и достоверно не различаются (табл. 2). Доля Cмик в общем Cорг минимальна в почве леса 110 лет (0.23%), несколько больше на пашне (0.38%), значительно возрастает в почвах 65- и 20-летнего леса (0.62–0.81%) и максимальна на участке сенокоса (1.26%). Микробный метаболический коэффициент QR максимален на участках спелого леса (0.86) и пашни (0.68). На остальных землепользованиях значения показателя близки и составляют 0.45–0.54.
Анализ структуры запасов углерода землепользований. Несмотря на то, что в почве пашни отмечено наименьшее содержание органического углерода в верхнем горизонте, запасы Cорг на данном участке максимальны и составляют 138 т С/га (рис. 3, А). В ряду молодой – средневозрастной – спелый лес запасы углерода в слое 0–100 см уменьшаются с 101.2 до 68.4 т С/га. Минимальное значение запасов Cорг почвы зафиксировано на участке сенокоса – 66.5 т С/га. Вклад верхних 50 см почвы в запасы углерода на пашне и сенокосе составляет 77%. На лесных участках в слое почвы 0–50 см сосредоточено до 85–89% органического углерода. Запасы Cорг в подстилке возрастают более чем в 4 раза – с 6.5 т/га в средневозрастном лесу до 27.2 т/га в спелом лесу. Так, в процентном соотношении к запасам углерода почвы, запасы Cорг в подстилке составляют 7% на участке леса 65 лет и 40% в лесу 110 лет. Если проанализировать сумму запасов органического углерода почвы в слое 0–100 см и подстилки (там где она есть), то общие запасы Cорг этих двух пулов близки по значению для всех лесных землепользований и составляют в среднем 98 т/га.
Рис. 3.
Вклад разных слоев (%) и общие запасы органического углерода (А) и углерода микробной биомассы (Б) в почве (n = 3) и подстилке (n = 6).
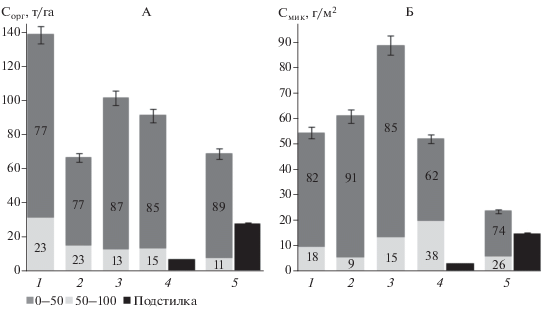
Структура запасов Cмик в почве и лесной подстилке заметно отличается от структуры запасов Cорг в данных пулах (рис. 3, Б). Запасы Cмик в слое почвы 0–100 см максимальны на участке молодого леса и составляют 88.9 г С/м2. Запасы Cмик убывают в ряду почв сенокос – пашня – 65-летний лес от 61 до 52 г С/м2. Минимальные значения содержания углерода микробной биомассы отмечены на участке спелого леса – 23.6 г С/м2. Значительная часть запасов Cмик сконцентрирована в верхней части профиля почв. Максимальная доля запасов Cмик в слое 0–50 см характерна для почв пашни, сенокоса и молодого леса – от 82 до 91%. В средневозрастном и спелом лесах вклад верхних 50 см почвы в запасы Cмик меньше и составляет 62 и 74%. Запасы Cмик в лесных подстилках изменяются пропорционально запасам Cорг и возрастают с 3.1 г С/м2 в средневозрастном лесу до 14.5 г С/м2 в спелом лесу. В процентном соотношении к запасам углерода микробной биомассы почвы, запасы Cмик в подстилках составляют 6% на участке 65-летнего леса и 61% в 110-летнем лесу. Несмотря на значительный вклад подстилки в запасы Cмик на участке спелого леса, суммарные запасы Cмик почвы и подстилки на данном участке низкое – 38.1 г С/м2. На участке средневозрастного леса запасы Cмик в этих двух пулах составляют 55.2 г С/м2.
Запасы углерода в фитомассе напочвенной растительности максимальны на участках сенокоса и молодого леса и составляют 9.3 и 7.7 т С/га (рис. 4, А). Данный показатель значительно уменьшается на участке спелого леса (3.6 т С/га), пашни (2.5 т С/га) и средневозрастного леса (2.2 т С/га). Доля углерода в подземной и надземной фитомассе напочвенного покрова на участке 20-летнего леса составляет 91%, на сенокосе и пашне она сокращается до 62 и 56%. В 65- и 110-летнем лесу в напочвенном покрове доля углерода надземной фитомассы больше и составляет 68 и 69% соответственно. Для лесных участков характерна линейная зависимость запасов углерода древесной растительности от возраста древостоя (рис. 4, Б). Запасы углерода в древесной фитомассе возрастают от 52.5 т С/га в 20-летнем лесу до 100 т С/га в 65-летнем лесу и составляют 159.5 т С/га в 110-летнем лесу. Последовательный рост запасов углерода древесной растительности происходит в основном за счет надземной фитомассы, доля которой составляет 88% в молодом лесу и 84–81% в средневозрастном и спелом лесах. Запасы углерода в дебрисе возрастают от 3.4 т С/га в 20-летнем лесу до почти 13 т С/га на остальных участках. Доля углерода мертвых органических остатков от углерода фитомассы древостоя составляет 7–13%.

Общие экосистемные запасы углерода (табл. 3) максимальны на лесных участках и возрастают от 164.8 т С/га в молодом лесу до 271.5 т С/га в спелом лесу. На участке сенокоса зафиксирован наименьший общий запас углерода – 75.8 т С/га, что почти в 2 раза меньше, чем на пашне. В исследуемом ряду землепользований соотношение углерода почвы к суммарным запасам углерода всех пулов участка последовательно уменьшается. На пашне запасы углерода почвы составляют 98%, на сенокосе – 88%, в молодом лесу – 61%, в 65-летнем лесу – 43%, и наконец, в 110-летнем лесу – 25%. Если на сельскохозяйственных угодьях превалирующим пулом углерода выступает Сорг почвы, то в зрелых лесных экосистемах 50–60% запасов углерода приходится на растительную фитомассу. Молодой лес занимает некоторое промежуточное положение, где на долю растительности приходится 37% от суммарных запасов углерода всех пулов участка. При этом доля подземной фитомассы на участках сенокос – спелый лес примерно одинакова и составляет 8–11% от общих запасов углерода. В лесных землепользованиях последовательно возрастает доля углерода мертвого органического вещества, сосредоточенного в дебрисе и подстилке. Так, если в 20-летнем лесу на него приходится лишь 2% от общих запасов углерода, то в 65-летнем – 9, а в 110-летнем – 15%.
Таблица 3.
Общие экосистемные запасы углерода (т C/га – над чертой) и доля каждого пула (% – под чертой)
| Пул | Пашня | Сенокос | Лес | ||
|---|---|---|---|---|---|
| 20 лет | 65 лет | 110 лет | |||
| Надземная фитомасса | 1.1/1 | 3.5/4 | 46.9/29 | 85.9/40 | 132.3/49 |
| Подземная фитомасса | 1.4/1 | 5.8/8 | 13.3/8 | 16.3/8 | 30.8/11 |
| Дебрис | 0 | 0 | 3.4/2 | 12.9/6 | 12.8/5 |
| Подстилка | 0 | 0 | 0 | 6.5/3 | 27.2/10 |
| Почва, 0–100 см | 138.0/98 | 66.5/88 | 101.2/61 | 90.9/43 | 68.4/25 |
| Сумма | 140.5/100 | 75.8/100 | 164.8/100 | 212.5/100 | 271.5/100 |
ОБСУЖДЕНИЕ
Почвы исследованных участков имеют различную степень агрогенной трансформации профиля, что влияет на их морфологическое строение, которое значительно отличается от естественной почвы (контроля) под спелым лесом. В изученном ряду почвообразованием охвачены лишь верхние 50–70 см, почвы имеют укороченный профиль, что типично для условий Карелии [43]. На пашне почва характеризуется предельной стадией агрогенной трансформации и утрачивает диагностические признаки, типичные для текстурно-дифференцированных почв, что позволяет выделить ее в отдел агроземов. На участках сенокоса и молодого леса происходит частичная постагрогенная трансформация профиля почв, в верхней части их текстурных горизонтов наблюдается возобновление элювиально-иллювиального процесса в виде зоны деградации горизонта BEL. При исследовании залежной сукцессии в южной тайге часто отмечается дифференциация верхнего гумусового горизонта в залежах 15–20 лет [5, 17, 38]. На изученных участках вспашка была прекращена около 30–40 лет назад, при этом морфологически агрогумусовые горизонты не претерпели значительных изменений и визуально сохраняют свою гомогенность [2]. Лишь на участке 65-летнеголеса серогумусовый горизонт подразделяется на 2 подгоризонта, а старопахотный горизонт имеет волнистую нижнюю границу. Следует отметить, что полноценные элювиальные горизонты сформированы только на участках с зональной растительностью, для них характерна четкая текстурная дифференциация при отсутствии переходного горизонта. Возможно, что в условиях среднетаежной подзоны морфологическое строение профиля агрогенно-нарушенных почв восстанавливается медленнее, чем в условиях южной тайги.
Изменение почвенных свойств с глубиной характеризует как агрогенное влияние на почвенный профиль, так и зональные особенности почв. Так, общая повышенная плотность текстурных горизонтов, низкие значения рН под лесной растительностью, а также выраженный элювиальный минимум углерода в профиле отражают генетические особенности текстурно-дифференцированных почв. На пашне почва характеризуется наибольшими показателями плотности и общим высоким уровнем рН в профиле, что типично для сельскохозяйственных почв подзолистого ряда [21, 22, 39]. Агрогенное влияние отражается также на содержании Cорг и типе распределения углерода по профилю. Отмечено значительное снижение содержания Cорг в верхней части профиля пашни и более равномерное распределение его по глубине. Наибольшее количество Cмик содержится в верхних органогенных горизонтах почв всех исследованных участков, содержание Cмик имеет тесную взаимосвязь с Cорг почвы [4, 79]. В целом для лесных почв и сенокоса отмечена более сильная дифференциация профиля по содержанию Cмик, по сравнению с пашней. В почвах всех категорий землепользования содержание Cмик глубже 50 см существенно уменьшается и имеет близкие значения. Под сенокосным лугом значительно сокращается мощность горизонта Р, нарастает элювиальный процесс [46] и по своим свойствам почва занимает промежуточное положение между пашней и лесом. В почвах лесных участков в целом уменьшается плотность, особенно значительно в гумусовых горизонтах, что сопряжено с растущим уровнем содержания углерода в верхних горизонтах почвы под лесной растительностью [34, 73]. Кислотность под лесными фитоценозами, напротив, растет. Считается, что смена растительности, кислая лесная подстилка и изменение состава гумуса играют ведущую роль в этом процессе [7, 39].
Наиболее заметное влияние смена землепользования оказывает на свойства верхних минеральных почвенных горизонтов. Почва под спелым лесом имеет самые низкие показатели рН, суммы обменных оснований, содержания подвижных фосфора и калия, а также общего азота в горизонте EL (табл. 2). Как известно, сельскохозяйственное освоение подзолистых почв ведет к значительному улучшению их агрохимического статуса [11, 12, 29], а, следовательно, функций продуктивности и депо элементов питания. По совокупности основных агрохимических показателей почву пашни можно охарактеризовать как сильноокультуренную, несмотря на экстенсивное использование. При длительном сенокосном использовании происходит повышение кислотности горизонта Р и некоторое уменьшение содержания обменных оснований, при этом содержание азота не меняется, а увеличение содержания Cорг статистически недостоверно. В ряду пашня – 65-летний лес содержание калия достаточно ровное, по-видимому, оно больше обусловлено минералогическим составом пород, чем агрогенным влиянием. Для содержания фосфора отмечен значительный разброс значений и низкое содержание в почвах под зональной растительностью, поскольку доступность данного элемента зависит как от остаточного фосфора удобрений, так и от кислотности почв [18, 27]. На участках пашни и сенокоса зафиксировано соотношение С/N, характерное для сельскохозяйственных дерново-подзолистых почв севера Европы [37]. В лесных почвах соотношение С/N увеличивается от молодого к спелому лесу, отражая возрастающее поступление органического вещества при уменьшении скорости его минерализации. В целом для лесных почв характерно ухудшение агрохимических свойств и функций продуктивности и депо элементов питания по сравнению с почвами сельскохозяйственных угодий [52].
Для почв изученного ряда отмечено изначально высокое содержание углерода. Содержание Cорг в горизонте EL спелого леса составляет около 2%, близкие значения получены для горизонтов Р на участках пашни и сенокоса. Увеличение содержания Cорг и азота в верхних горизонтах отмечено на участках 20- и 65-летних лесов. Также для этих участков и сенокоса характерны наибольшие показатели Cмик, возрастающие при восстановительной сукцессии под лесной растительностью [10]. На пашне содержание Cмик в верхнем горизонте низкое, как и в целом по профилю, что типично для нарушенных экосистем [13, 35, 82]. Минимальные значения содержания Cмик получены для спелого леса, что типично для элювиальных горизонтов по сравнению с органогенными [36]. Доля углерода микробной биомассы в общем содержании органического углерода почвы Cмик/Cорг является важным экологическим показателем, характеризующим состояние микробного сообщества и степень зрелости экосистемы [62, 68]. Для почв 20- и 65-летних лесов и особенно сенокоса установлено более высокое соотношение Cмик/Cорг в органогенных горизонтах и увеличение доли микробного углерода в общем углероде верхних горизонтов. Следовательно, трансформационная функция почв на данных участках повышена по сравнению с пашней и спелым лесом. Микробный метаболический коэффициент QR показывает физиологический статус микробного сообщества, он чувствителен к нарушениям в почве. Обычно QR увеличивается в верхних горизонтах пахотных и нарушенных почв по сравнению с естественными почвами [50, 68], что также отмечено в нашем исследовании. В целом близкие к 1 значения QR характерны для зоны дерново-подзолистых почв [20], для всех исследованных участков функционирование микробоценозов можно считать сбалансированным.
По запасам органического углерода в метровом слое почва пашни превосходит почву под спелым лесом в 2 раза (рис. 3, А). Многие исследователи отмечают рост запасов углерода почвы при восстановительной сукцессии лесной растительности [5, 26, 33]. Однако встречаются работы, отмечающие уменьшение запасов углерода в почвах при лесовосстановлении по сравнению с сельскохозяйственными угодьями [23, 32, 60, 64]. Такие данные зачастую получают для почв, сформированных под хвойной растительностью [52, 61], где возрастает количество трудноразлагаемого опада по сравнению с легкоразлагаемыми травянистыми растительными остатками и органическими удобрениями на пашне [70, 77]. Несмотря на увеличение содержания Cорг в верхних горизонтах лесных почв, общие запасы углерода в них могут сокращаться. В целом смена землепользования может оказывать существенное влияние на запасы углерода в верхней части почвенного профиля, а в более глубоких слоях запасы Cорг остаются неизменными [54, 80]. В нашем случае большие запасы углерода в почве пашни обеспечены за счет повышенной плотности и равномерного распределения Cорг в профиле почвы. В лесных почвах уменьшаются как общие запасы углерода, так и вклад слоя 50–100 см, Cорг концентрируется в верхней части почвенного профиля. Значительный вклад в запасы углерода зрелых лесов вносит Cорг подстилки. Для сенокосного участка получены самые низкие запасы углерода (66.5 т С/га), несколько больше запасы Cорг под спелым лесом (68.4 т С/га). Считается, что для каждого типа почв характерно свое стационарное состояние и запас углерода, в зависимости от природной зоны. Так, для суглинистых почв южной тайги запас углерода почвы в стационарном состоянии составляет около 70 т С/га [33]. По-видимому, запасы углерода почвы стремятся к стационарному состоянию под растительными сообществами, близкими к своему экологическому оптимуму не только под зональной растительностью (в нашем случае вторичный ельник чернично-кисличный в субклимаксной стадии сукцессии), но и под полуприродной системой, каковой является суходольный луг в климаксной стадии сукцессии [14].
Максимальные запасы углерода микробной биомассы получены для почвы в 20-летнем лесу (рис. 3, Б). Участкам сенокоса и молодого леса, соответствуют максимальные запасы углерода, сосредоточенного в фитомассе напочвенной растительности, особенно подземной ее части (рис. 4, А). Количество и качество поступающего на почву опада определяет интенсивность микробиологической активности и качества микробного сообщества данных землепользований [5, 78]. Запасы Cмик в слое 0–100 см в почвах пашни, сенокоса и средневозрастного леса имеют близкие значения, а минимальные значения запасов Cмик отмечены в почве спелого леса. На участках 65- и 110-летнего леса в напочвенном покрове преобладают кустарнички, и увеличивается доля трудногидролизуемого опада из надземной фитомассы. Для почвы спелого ельника в целом характерны низкие параметры микробиологической активности, а основной пул Cмик приурочен к лесной подстилке (рис. 3, Б). Вклад верхних 50 см в запасы Cмик в почвах сельхозугодий и молодого леса достаточно высокий (82–91%). В почвах средневозрастного и спелого лесов в отличие от Cорг возрастает вклад слоя 50–100 см в запасы Cмик.
Общие экосистемные запасы углерода, складывающиеся из пяти пулов, имеют структуру характерную для таежной зоны [33, 44]. Запас углерода максимален в лесных экосистемах, где наибольшая скорость накопления углерода приходится на время активного формирования древесного яруса [19] и составляет 165–272 т С/га. Древесная растительность лесных участков характеризуется высокой продуктивностью и классом бонитета [28], что обусловлено более благоприятными для лесовозобновления почвенными свойствами бывших сельхозугодий [3, 6]. В зрелых лесах 50–60% углерода участка приходится на растительную фитомассу (табл. 3). В целом зрелые лесные экосистемы обладают максимальной функцией секвестрации углерода за счет надземной биомассы. Доля связанных с фитомассой пулов мертвого органического вещества дебриса и подстилки возрастает с 2% в молодом до 15% в спелом лесу. В молодом лесу доля почвенного углерода достаточно велика и составляет 61% от общего баланса, под зональной растительностью доля Cорг почвы уменьшается и составляет 43–25%. Доля почвенного углерода для сельскохозяйственных угодий типично высока [33], Cорг почвы составляет 88% от общего баланса углерода на сенокосе и 98% на пашне.
ЗАКЛЮЧЕНИЕ
В среднетаежной подзоне Карелии на легкосуглинистых почвах нормального увлажнения смена землепользования приводит к значительным изменениям морфологического строения почвенного профиля, агрохимических и микробиологических свойств почв, запасов углерода почвы и экосистемы в целом. Почвы исследованных участков характеризуются различной степенью агрогенной и постагрогенной трансформации по сравнению с типичной подзолистой почвой под спелым лесом. На пашне отмечена максимальная степень трансформации профиля, который утрачивает диагностические признаки, типичные для текстурно-дифференцированных почв. Под средневозрастным лесом почва по морфологическому строению близка к естественным зональным почвам и имеет отчетливо выраженный элювиальный горизонт. Наиболее заметное влияние смена землепользования оказывает на свойства верхних минеральных почвенных горизонтов. Для пашни характерна наибольшая степень уплотнения почвы, пониженная кислотность и высокое содержание доступного фосфора, калия и суммы обменных оснований. В почве сенокоса отмечено некоторое ухудшение агрохимических свойств, уменьшение плотности и увеличение кислотности. В целом для сельскохозяйственных землепользований отмечен наиболее высокий уровень функции продуктивности и депо элементов питания. На лесных участках значительно увеличивается кислотность, уменьшается плотность и ухудшаются агрохимические свойства, особенно в почве спелого леса. По показателям экофизиологического статуса Cмик/Cорг и QR функционирование микробоценозов на всех исследованных участках можно считать сбалансированным. Для почв пашни и спелого леса характерно пониженное содержание углерода микробной биомассы, а на участках сенокоса, молодого и средневозрастного лесов зафиксирован максимальный уровень Смик и факторов микробного благополучия, что ассоциируется с растущим уровнем Сорг и трансформационной функцией почвы. Максимальные запасы Смик в метровом слое почвы отмечены под молодым лесом, что связано с поступлением большого количества легкогидролизуемого опада. В спелом лесу основной пул Cмик приурочен к лесной подстилке. Запасы Сорг почвы в метровом слое максимальны на пашне (138 т С/га) и сокращаются на участках молодого и средневозрастного лесов до 101–91 т С/га. Под растительными сообществами, близкими к своему экологическому оптимуму – спелым лесом и сенокосом, запасы Сорг минимальны и составляют 67–68 т С/га. Общие экосистемные запасы углерода, ассоциированные с функцией секвестрации углерода, максимальны для зрелых лесов, и минимальны на сенокосе. В молодом лесу доля почвенного углерода составляет 61%, а под зональной растительностью сокращается до 43–25% от общего баланса. До 50–60% углерода участка в зрелых лесах приходится на растительную фитомассу. Возрастает также доля связанных с ней пулов мертвого органического вещества дебриса и подстилки. Для сельскохозяйственных угодий ведущим пулом является Сорг почвы, он составляет 88% от общего баланса углерода на сенокосе и 98% на пашне.
Список литературы
Атлас Карельской АССР / Под ред. А.Г. Дурова. М.: ГУГК СССР, 1989. 40 с.
Баранова О.Ю., Номеров Г.Б., Строганова М.Я. Изменения свойств пахотных дерново-подзолистых почв при зарастании их лесом // Почвообразование в лесных биогеоценозах. М.: Наука, 1989. С. 60–78.
Беляева Н.В., Данилов Д.А., Кази И.А. Особенности естественного возобновления ели европейской на постагрогенных землях // Актуальные проблемы лесного комплекса. 2019. № 54. С. 6–10.
Богородская А.В., Шишикин А.С. Динамика микробной биомассы, ее структура и функциональная активность в почвах при лесовозобновлении на вырубках пихтарников Енисейского кряжа // Почвоведение. 2020. № 1. С. 119–130. https://doi.org/10.31857/S0032180X20010050
Владыченский А.С., Телеснина В.М., Румянцева К.А., Чалая Т.А. Органическое вещество и биологическая активность постагрогенных почв южной тайги на примере Костромской области // Почвоведение. 2013. № 5. С. 518–529. https://doi.org/10.7868/S0032180X1305016X
Голубева Л.В., Наквасина Е.Н., Минин Н.С. Продуктивность и качество древесины сосны обыкновенной (Pínus sylvestris L.) в постагрогенных насаждениях // Известия Санкт-Петербургской лесотехнической академии. 2016. № 215. С. 19–29. https://doi.org/10.21266/2079-4304.2016.215.19-29
Дричко В.Ф., Литвинович А.В., Павлова О.Ю., Чернов Д.В., Буре В.М. Скорости изменения кислотно-основных параметров, содержания общего углерода и состава гумуса в дерново-подзолистой песчаной почве при переходе от пашни к лесу при сукцессии залежных земель // Агрохимия. 2015. № 11. С. 19–29.
Дымов А.А., Милановский Е.Ю. Изменение органического вещества таежных почв в процессе естественного лесовозобновления растительности после рубок (средняя тайга республики Коми) // Почвоведение. 2014. № 1. С. 39–47. https://doi.org/10.7868/S0032180X14010043
Евдокимов И.В. Методы определения биомассы почвенных микроорганизмов // Rus. J. Ecosystem. Ecol. 2018. Т. 3. № 3. С. 1–20. https://doi.org/10.21685/2500-0578-2018-3-5
Ерохова А.А., Макаров М.И., Моргун Е.Г., Рыжова И.М. Изменение состава органического вещества дерново-подзолистых почв в результате естественного восстановления леса на пашне // Почвоведение. 2014. № 11. С. 1308–1314. https://doi.org/10.7868/S0032180X14110045
Завьялова Н.Е. Гумус и азот дерново-подзолистой почвы различных сельскохозяйственных угодий Пермского края // Почвоведение. 2016. № 11. С. 1347–1354. https://doi.org/10.7868/S0032180X16110113
Иванов И.А., Иванов А.И., Цыганова Н.А. Изменение свойств подзолистых и дерново-подзолистых почв на песчаных породах при окультуривании // Почвоведение. 2004. № 4. С. 489–499.
Иващенко К.В., Ананьева Н.Д., Васенев В.И., Кудеяров В.Н., Валентини Р. Биомасса и дыхательная активность почвенных микроорганизмов в антропогенно-измененных экосистемах (Московская область) // Почвоведение. 2014. № 9. С. 1077–1088. https://doi.org/10.7868/S0032180X14090056
Исаков Ю.А., Казанская Н.С., Тишков А.А. Зональные закономерности динамики экосистем. М.: Наука, 1986. 148 с.
Казимиров Н.И., Морозова Р.М. Биологический круговорот веществ в ельниках Карелии. Л.: Наука, 1973. 175 с.
Казимиров Н.И., Морозова Р.М., Куликова В.К. Органическая масса и потоки веществ в березняках средней тайги. Л.: Наука, 1979. 216 с.
Кечайкина И.О., Рюмин А.Г., Чуков С.Н. Постагрогенная трансформация органического вещества дерново-подзолистых почв // Почвоведение. 2011. № 10. С. 1178–1192.
Кирпичников Н.А., Адрианов С.Н. Действие и последействие фосфорных удобрений на дерново-подзолистой тяжелосуглинистой почве при различной степени известкования // Агрохимия. 2007. № 10. С. 14–23.
Кудеяров В.Н. Современное состояние углеродного баланса и предельная способность почв к поглощению углерода на территории России // Почвоведение. 2015. № 9. С. 1049–1060. https://doi.org/10.7868/S0032180X15090087
Курганова И.Н., Лопес де Гереню В.О., Мостовая А.С., Овсепян Л.А., Телеснина В.М., Личко В.И., Баева Ю.И. Влияние процессов естественного лесовосстановления на микробиологическую активность пост-агрогенных почв европейской части России // Лесоведение. 2018. № 1. С. 3–23. https://doi.org/10.7868/S0024114818010011
Литвинович А.В. Постагрогенная эволюция хорошо окультуренных дерново-подзолистых почв Северо-Запада Нечерноземной зоны // Агрохимия. 2009. № 7. С. 85–93.
Литвинович А.В., Дричко В.Ф., Павлова О.Ю., Чернов Д.В., Шабанов М.В. Изменение кислотно-основных свойств окультуренных дерново-подзолистых почв легкого гранулометрического состава в процессе постагрогенной трансформации // Почвоведение. 2009. № 6. С. 680–686.
Литвинович А.В., Павлова О.Ю. Изменение гумусного состояния дерново-подзолистой глееватой песчаной почвы на залежи // Почвоведение. 2007. № 11. С. 1323–1329.
Лиханова И.А., Лаптева Е.М., Ковалева В.А. Продуктивность растительных сообществ и формирование органогенных горизонтов почв в ходе самовосстановительной сукцессии на техногенных субстратах // Известия СамНЦ РАН. 2014. Т. 16. № 5. С. 182–189.
Лиханова Н.В., Бобкова К.С. Пулы и потоки углерода в экосистемах вырубки ельников средней тайги Республики Коми // Теорeтическая и прикладная экология. 2019. № 2. С. 91–100. https://doi.org/10.25750/1995-4301-2019-2-091-100
Люри Д.И., Горячкин С.В., Караваева Н.А., Щенисенко E.A., Нефедова Т.Т. Динамика сельскохозяйственных земель России в XX веке и постагрогенное восстановление растительности и почв. М.: ГЕОС, 2010. 416 с.
Митрофанова Е.М. Влияние длительного применения минеральных удобрений и последействия извести на фосфатный режим дерново-поверхностно-подзолистой почвы Предуралья // Агрохимия. 2016. № 7. С. 36–43.
Мошкина Е.В., Медведева М.В., Туюнен А.В., Карпечко А.Ю., Геникова Н.В., Дубровина И.А., Мамай А.В., Сидорова В.А., Толстогузов О.В., Кулакова Л.М. Особенности естественного восстановления лесных экосистем на бывших сельскохозяйственных землях (на примере Южного агроклиматического района Карелии) // Биосфера. 2019. Т. 11. № 3. С. 134–145. https://doi.org/10.24855/biosfera.v11i3.506
Муха В.Д. Естественно-антропогенная эволюция почв (общие закономерности и зональные особенности). М.: КолосС, 2004. 271 с.
Полубояринов О.И. Плотность древесины. М.: Лесная промышленность, 1976. 160 с.
Почвенная карта и объяснительная записка к почвенной карте агробиологической станции 1977–1980 гг. Институт биологии КарНЦ РАН. Петрозаводск, 1980. 153 с.
Романовская А.А., Коротков В.Н., Карабань Р.Т., Смирнов Н.С. Динамика элементов баланса углерода на неиспользуемых пахотных угодьях Валдайской возвышенности // Экология. 2012. № 5. С. 347–352.
Рыжова И.М., Ерохова А.А., Подвезенная М.А. Динамика и структура запасов углерода в постагрогенных экосистемах южной тайги // Почвоведение. 2014. № 12. С. 1426–1435. https://doi.org/10.7868/S0032180X14090111
Рыжова И.М., Ерохова А.А., Подвезенная М.А. Изменение запасов углерода в постагрогенных экосистемах в результате естественного восстановления лесов в Костромской области // Лесоведение. 2015. № 4. С. 307–317.
Семенюк О.В., Ильяшенко М.А., Бобрик А.А. Оценка экологических функций парковых почв на основе показателей их биологической активности // Проблемы агрохимии и экологии. 2013. № 3. С. 35–39.
Стольникова Е.В., Ананьева Н.Д., Чернова О.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов европейской территории России // Почвоведение. 2011. № 4. С. 479–494.
Сычев В.Г., Шевцова Л.К., Беличенко М.В., Рухович О.В., Иванова О.И. Взаимодействие циклов углерода и азота в основных типах почв при длительном применении различных систем удобрения // Проблемы агрохимии и экологии. 2018. № 4. С. 68–77. https://doi.org/10.26105/AE.2018.4.33.016
Телеснина В.М., Ваганов И.Е., Карлсен А.А., Иванова А.Е., Жуков М.А., Лебедев С.М. Особенности морфологии и химических свойств постагрогенных почв южной тайги на легких отложениях (Костромская область) // Почвоведение. 2016. № 1. С. 115–129. https://doi.org/10.7868/S0032180X16010111
Телеснина В.М., Курганова И.Н., Лопес де Гереню В.О., Овсепян Л.А., Личко В.И., Ермолаев А.М., Мирин Д.М. Динамика свойств почв и состава растительности в ходе постагрогенного развития в разных биоклиматических зонах // Почвоведение. 2017. № 12. С. 1514–1534. https://doi.org/10.7868/S0032180X17120115
Теория и практика химического анализа почв / Под ред. Л.А. Воробьевой. М.: ГЕОС, 2006. 400 с.
Трейфельд Р.Ф., Кранкина О.Н. Определение запасов и фитомассы древесного детрита на основе данных лесоустройства // Лесное хозяйство. 2001. № 4. С. 23–26.
Уткин А.И., Замолодчиков Д.Г., Честных О.В. Углеродные пулы фитомассы, почв и депонирование углерода в еловых лесах России // Хвойные бореальной зоны. 2004. Вып. 2. С. 21–30.
Федорец Н.Г., Бахмет О.Н. Особенности формирования почв и почвенного покрова Карело-Кольского региона // Тр. Карельского научного центра РАН. 2016. № 12. С. 39–51. https://doi.org/10.17076/eco358
Чернова О.В., Рыжова И.М., Подвезенная М.А. Влияние исторических и региональных особенностей землепользования на величину и структуру запасов углерода в южной тайге и лесостепи европейской России // Почвоведение. 2018. № 6. С. 747–758. https://doi.org/10.7868/S0032180X18060114
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Якименко Е.Ю. Сравнительная характеристика почвообразования в луговых и лесных биогеоценозах // Почвообразование в лесных биогеоценозах. М.: Наука, 1989. С. 79–102.
Adhikari K., Hartemink A.E. Linking soils to ecosystem services – A global review // Geoderma. 2016. V. 262. P. 101–111. https://doi.org/10.1016/j.geoderma.2015.08.009
Akujärvi A., Heikkinen J., Palosuo T., Liski J. Carbon budget of Finnish croplands – Effects of land use change from natural forest to cropland // Geoderma Regional. 2014. V. 2–3. P. 1–8. https://doi.org/10.1016/j.geodrs.2014.09.003
Anderson J., Domsch K.H. A physiological method for the quantitative measurement of microbial biomas in soils // Soil Biol. Biochem. 1978. № 10. P. 215–221.
Balota E.L., Filho A.C., Andrade D.S., Dick R.P. Long-term tillage and crop rotation effects on microbial biomass and C and N mineralization in a Brazilian Oxisol // Soil & Tillage Research. 2004. V. 77. P. 137–145. https://doi.org/10.1016/j.still.2003.12.003
Baveye P.C., Baveye J., Gowdy J. Soil “ecosystem” services and natural capital: critical appraisal of research on uncertain ground // Front. Environ. Sci. 2016. V. 4. P. 41. https://doi.org/10.3389/fenvs.2016.00041
Berthrong S.T., Jobbagy E.G., Jackson R.B. A global meta-analysis of soil exchangeable cations, pH, carbon and nitrogen with afforestation // Ecological Applications. 2009. V. 19. P. 2228–2241. https://doi.org/10.1890/08-1730.1
Blum W.E.H. Functions of soil for society and the environment // Environmental Science and Bio/Technology. 2005. V. 4(3). P. 75–79. https://doi.org/10.1007/s11157-005-2236-x
Borchard N., Bulusu M., Meyer N., Rodionov A., Herawati H., Blagodatsky S., Cadisch G., Welp G., Ame-lung W., Martius C. Deep soil carbon storage in tree-dominated land use systems in tropical lowlands of Kalimantan // Geoderma. 2019. V. 354. P. 113–864. https://doi.org/10.1016/j.geoderma.2019.07.022
Bouma J. Soil science contributions towards Sustainable Development Goalsand their implementation: linking soil functions with ecosystem services // J. Soil Sc. Plant Nutrition. 2014. V. 177(2). P. 111–120. https://doi.org/10.1002/jpln.201300646
Deng L., Zhu G.Y., Tang Z.S., Shangguan Z.P. Global patterns of the effects of land-use changes on soil carbon stocks // Global Ecology and Conservation. 2016. V. 5. P. 127–138. https://doi.org/10.1016/j.gecco.2015.12.004
Falkengen-Grerup U., ten Brink D.-J., Brunet J. Land use effects on soil N, P, C and pH persist over 40–80 years of forest growth an agricultural soils // Forest Ecol. Manag. 2006. V. 225. P. 74–81. https://doi.org/10.1016/j.foreco.2005.12.027
Fuchs R., Schulp C.J.E., Hengeveld G.M., Verburg P.H., Clevers J.G.P.W., Schelhaas M.J., Herold M. Assessing the influence of historic net and gross land changes on the carbon fluxes of Europe // Global Change Biology. 2016. V. 22. P. 2526–2539. https://doi.org/10.1111/gcb.13191
Greiner L., Keller A., Grêt-Regamey A., Paprit A. Soil function assessment: review of methods for quantifying the contributions of soils to ecosystem services // Land Use Policy. 2017. V. 69. P. 224–237. https://doi.org/10.1016/j.landusepol.2017.06.025
Guidi C., Vesterdal L., Gianelle D., Rodighero M. Changes in soil organic carbon and nitrogen following forest expansion on grassland in the Southern Alps // Forest Ecol. Manag. 2014. V. 328. P. 103–116.https://doi.org/10.1016/j.foreco.2014.05.025
Guo L.B., Gifford R.M. Soil carbon stocks and land use change: a meta analysis // Global Change Biology. 2002. V. 8. P. 345–360. https://doi.org/10.1046/j.1354-1013.2002.00486.x
Insam H., Domsch K.H. Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites // Microbial Ecology. 1988. V. 15. P. 177–188. https://doi.org/10.1007/BF02011711
IPCC Good Practice Guidance for Land Use, Land-Use Change and Forestry. IPCC National Greenhouse Gas Inventories Programme. IGES, Japan, 2003. 590 p.
La Mantia T., Gristina L., Rivaldo E., Pasta S., Novara A., Rühl J. The effect of post-pasture woody plant colonization on soil and aboveground litter carbon and nitrogen along bioclimatic transect // iForest. 2013. V. 6(4). P. 238–246. https://doi.org/10.3832/ifor0811-006
Le Quéré C., Raupach M.R., Canadell J.G. et al. Trends in the sources and sinks of carbon dioxide // Nature Geoscience. 2009. V. 2. P. 831–836. https://doi.org/10.1038/ngeo689
Lehmann A., Stahr K. The potential of soil functions and planner-oriented soil evaluation to achieve sustainable land use // J. Soils Sediments. 2010. V. 10. P. 1092–1102. https://doi.org/10.1007/s11368-010-0207-5
Nannipieri P., Eldor P. The chemical and functional characterization of soil N and its biotic components // Soil Biol. Biochem. 2009. V. 41. P. 2357–2369. https://doi.org/10.1016/j.soilbio.2009.07.013
Pabst H., Gerschlauer F., Kiese R., Kuzyakov Y. Land Use and Precipitation Affect Organic and Microbial Carbon Stocks and the Specific Metabolic Quotient in Soils of Eleven Ecosystems of Mt. Kilimanjaro, Tanzania // Land Degradation and Development. 2016. V. 27. P. 592–602. https://doi.org/10.1002/ldr.2406
Leal Pacheco F.A., Sanches Fernandes L.F., Valle Junior R.F., Valera C.A., Tarlé Pissarra T.C. Land degradation: multiple environmental consequences and routes to neutrality // Current Opinion in Environmental Science & Health. 2018. V. 5. P. 79–86. https://doi.org/10.1016/j.coesh.2018.07.002
Poeplau C., Don A., Vesterdal L., Leifeld J., van Wesemael B., Schumacher J., Gensior A. Temporal dynamics of soil organic carbon after land_use change in the temperate zone – carbon response functions as a model approach // Global Change Biology. 2011. V. 17. P. 2415–2427. https://doi.org/10.1111/j.1365-2486.2011.02408.x
Ramesh T, Bolan N. S., Kirkham M. B., Wijesekara H., Kanchikerimath M., Rao C.S., Sandeep S., Rinklebe J., Ok Y.S., Choudhury B.U., Wang H., Tang C., Wang X., Song Z., Freeman O.W. Soil organic carbon dynamics: Impact of land use changes and management practices: A review // Adv. Agronomy. 2019. V. 156. P. 1–107. https://doi.org/10.1016/bs.agron.2019.02.001
Risch A.C., Jurgensen M.F., Page-Dumroese D.S., Wildi O., Schütz M. Long-term development of above- and below-ground carbon stocks following land-use change in subalpine ecosystems of the Swiss National Park // Can. J. For. Res. 2008. V. 38. P. 1590–1602. https://doi.org/10.1139/X08-014
Šantrůčková H., Kaštovská E., Kozlov D., Kurbatova J., Livečková M., Shibistova O., Tatarinov F., Lloyd J. Vertical and horizontal variation of carbon pools and fluxes in soil profile of wet Southern taiga in European Russia // Boreal Environment Research. 2010. V. 15(3). P. 357–369. doi 10138/233132
Schwilch G., Bernet L., Fleskens L., Giannakis E., Leventon J., Maranón T., Mills J., Short C., Stolte J., van Delden H., Verzandvoort S. Operationalizing ecosystem services for the mitigation of soil threats: A proposed framework // Ecological Indicators. 2016. V. 67. P. 586–597. https://doi.org/10.1016/j.ecolind.2016.03.016
Shao P, Liang C, Rubert-Nason K, Li X, Xie H, Bao X. Secondary successional forests undergo tightly-coupled changes in soil microbial community structure and soil organic matter // Soil Biol. Biochem. 2019. V. 128. P. 56–65. https://doi.org/10.1016/j.soilbio.2018.10.004
Smal H., Olszewska M. The effect of afforestation with Scots pine (Pinus silvestris L.) of sandy post-arable soils on their selected properties. II. Reaction, carbon, nitrogen and phosphorus // Plant Soil. 2008. V. 305. P. 171–187. https://doi.org/10.1007/s11104-008-9538-z
Thuille A., Schulze E.-D. Carbon dynamics in successional and afforested spruce stands in Thuringia and in the Alps // Global Change Biology. 2006. V. 12. P. 325–342. https://doi.org/10.1111/j.1365-2486.2005.01078.x
Vladychensky A.S., Telesnina V.M., Chalaya T.A. Influence of fallen plant leaves on biological activity of post-agrogenic soils of southern taiga // Moscow Univ. Soil Sci. Bull. 2012. V. 67. P. 1–7. https://doi.org/10.3103/S0147687412010085
Wang Y., Liu X., Chen F., Huang R., Deng X., Jiang Y. Seasonal dynamics of soil microbial biomass C and N of Keteleeria fortunei var. cyclolepis forests with different ages // J. Forestry Research. 2020. V. 31. P. 2377–2384. https://doi.org/10.1007/s11676-019-01058-w
Wiesmeier M., von Lützow M., Sporlein P., Geuss U., Hangen E., Reischl A., Schilling B., Kögel-Knabner I. Land use effects on organic carbon storage in soils of Bavaria: the importance of soil types // Soil & Tillage Research. 2015. V. 146. P. 296–302. https://doi.org/10.1016/j.still.2014.10.003
Wiesmeier M., Urbanski L., Hobley E., Lang B., von Lützow M., Marin-Spiotta E., van Wesemael B., Rabot E., Ließ M., Garcia-Franco N., Wollschläger U., Vogel H.-J., Kögel-Knabner I. Soil organic carbon storage as a key function of soils – A review of drivers and indicators at various scales // Geoderma. 2019. V. 333. P. 149–162. https://doi.org/10.1016/j.geoderma.2018.07.026
Xu X., Thornton P.E., Post W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems // Global Ecology and Biogeography. 2013. V. 22. P. 737–749. https://doi.org/10.1111/geb.12029
Дополнительные материалы отсутствуют.