

Почвоведение, 2021, № 2, стр. 232-239
Влияние различных типов землепользования на прокариотные сообщества и стабилизацию органического вещества дерново-подзолистой почвы
Н. Е. Завьялова a, *, И. Г. Широких b, М. Т. Васбиева a, Д. С. Фомин a
a Пермский федеральный исследовательский центр УрО РАН
614532 Пермский край, Пермский район, Лобаново, ул. Культуры, 12, Россия
b Федеральный научный центр Северо-Востока им. Н.В. Рудницкого
610007 Киров, ул. Ленина, 166а, Россия
* E-mail: nezavyalova@gmail.com
Поступила в редакцию 13.02.2020
После доработки 01.06.2020
Принята к публикации 07.06.2020
Аннотация
В длительном стационарном опыте (1977–2018 гг.) на дерново-подзолистой почве (Eutric Albic Retisols (Abruptic, Loamic, Cutanic)) Пермского края изучали влияние севооборотов с различным насыщением бобовыми травами и бессменных посевов зерновых культур (озимая рожь, яровой ячмень) на структуру прокариотных сообществ и сохранность органического вещества (ОВ). В качестве эталонов сравнения использовали бессменный чистый пар и залежь. Минимальные значения коэффициентов минерализации (0.37) и педотрофности (0.28), найденные на основании учета численности при посеве на традиционные среды МПА, КАА и ПА, выявлены в залежной почве, максимальные (1.97 и 1.30 соответственно) – в парующей почве. Обнаружена обратная зависимость между содержанием в почве Cорг и коэффициентами минерализации (r = –0.67; p < 0.01); между соотношением СГК/СФК в составе ОВ почвы и коэффициентами ее педотрофности (r = –0.64; p < 0.02). Показано, что стабилизация ОВ дерново-подзолистой почвы зависит от типа использования, ухудшаясь в ряду: залежь > севообороты (0–28.6–42.9% бобовых) > бессменный посев зерновых > бессменный чистый пар. В возделываемых и парующей почвах, наряду с общим уменьшением запаса ОВ, повысилась его лабильность, о чем свидетельствуют более низкие, чем в залежной почве (0.96), соотношения СГК/СФК (0.55–0.79). Между коэффициентами минерализации и показателями СГК/СФК в почве длительного стационара прослеживалась наиболее тесная отрицательная корреляция (r = –0.81; p < 0.001). Показано также, что различное использование дерново-подзолистой почвы сопровождалось структурными перестройками комплекса актиномицетов, выбранных в качестве модельной группы почвенных микроорганизмов. В частности, регулярная механическая обработка почвы и внесение минеральных удобрений (N60P30K60) способствовали увеличению видового спектра стрептомицетов, изменению частоты встречаемости и долевого участия представителей отдельных секций и серий, смене доминантных форм. Полученные результаты указывают на возможность использования микробиологических показателей в качестве биомаркеров состояния почвенного ОВ.
ВВЕДЕНИЕ
Проблема рационального использования органического вещества (ОВ) приобрела большое научно-практическое значение в связи со значительными его потерями в пахотных почвах, как в нашей стране [2, 5], так и за рубежом [1, 9]. К числу основных факторов, контролирующих степень дегумификации почв, относят тип землепользования, структуру севооборотов, обработку почвы, системы удобрений и др.
Вопросы стабилизации ОВ в агроэкосистемах не могут быть решены без выявления роли микроорганизмов в формировании бездефицитного баланса. Результаты ранее выполненной оценки величины углерода микробной биомассы по вариантам варьируют от 1.50 до 3.24% от общих запасов почвенного ОВ дерново-подзолистой тяжелосуглинистой почвы [7]. Очевидно, это является причиной недостаточного внимания к важной роли почвенной микробиоты в динамике ОВ. Между тем для разработки мер, препятствующих развитию потерь ОВ, повсеместно проводятся исследования, направленные на выявление связей между динамикой ОВ почвы и другими микробиологическими показателями в разных почвенно-климатических зонах и типах почв [10–12, 14, 16].
Действие различных биологических факторов и агротехнических приемов на состояние ОВ и плодородие почвы становится очевидным по истечении десятков лет. Поэтому особую ценность представляют исследования, выполненные в длительных полевых стационарах, где на фоне изменения факторов окружающей среды действие изучаемого фактора накапливается во времени.
Цель работы – сравнительная оценка состояния почвенных микробных сообществ и ОВ, сформировавшихся за сорокалетний период при различных типах землепользования в тяжелосуглинистой дерново-подзолистой почве Предуралья.
ОБЪЕКТЫ И МЕТОДЫ
Исследования выполнены в условиях длительного стационарного опыта, заложенного в 1977 г. на дерново-подзолистой тяжелосуглинистой почве (Eutric Albic Retisols (Abruptic, Loamic, Cutanic)). Варианты опыта: 1 – чистый пар бессменно; 2 – ячмень бессменно; 3 – озимая рожь бессменно; 4 – зерновой пятипольный севооборот (0% бобовых культур) (ячмень, рожь озимая, пшеница яровая, ячмень, овес); 5 – полевой семипольный севооборот (типичный, 28.6% бобовых культур) (унавоженный чистый пар, рожь озимая, пшеница яровая с подсевом клевера, клевер 1 года пользования (г. п.), клевер 2 г. п., ячмень, овес); 6 – полевой семипольный севооборот (42.9% бобовых культур) (сидеральный пар (клевер 1 г. п.), рожь озимая, пшеница яровая с подсевом клевера, клевер 1 г. п., клевер 2 г. п., ячмень, овес с подсевом клевера); 7 – залежь. Представленные варианты изучали без применения минеральных удобрений и при внесении удобрений в дозе N по 60, Р по 30 и К по 60 кг д. в./га. Минеральные удобрения вносили под зерновые культуры, на клевере изучали последействие. В опыте использовали аммиачную селитру или мочевину, хлористый калий, суперфосфат. В паровом поле типичного для Предуралья полевого севооборота вносили подстилочный навоз в дозе 40 т/га пашни.
На момент закладки опыта почва имела следующие агрохимические показатели: рНKCl 5.2–5.3, гидролитическая кислотность 2.1–2.3 смоль (экв)/кг, сумма поглощенных оснований 14.0–15.5 смоль (экв)/кг, содержание органического углерода по Тюрину 1.10–1.12%, подвижного фосфора 225–240 мг/кг, подвижного калия 196–204 мг/кг (по Кирсанову). Почвенные образцы для исследований отбирали осенью 2018 г. с двух несмежных повторений в слое 0–20 см.
Для микробиологического анализа из каждого образца брали по две усредненных навески массой 1.0 г. Количество микроорганизмов определяли методом посева из разведений почвенных суспензий на мясо-пептонном (МПА), крахмало-аммиачном (КАА), почвенном (ПА) агаре. Количество колониеобразующих единиц различных групп микроорганизмов пересчитывали на 1 г воздушно-сухой почвы с учетом разведения суспензий в 104 раз при посеве на МПА и КАА, в 105 раз – при посеве на ПА. Для характеристики изменений в структуре почвенного микробного сообщества по вариантам рассчитывали коэффициент минерализации (КАА/МПА), отражающий интенсивность мобилизации азота, и коэффициент педотрофности (ПА/МПА), характеризующий развитие в почве автохтонной микрофлоры [6].
При выявлении структурных перестроек микробного сообщества в качестве модельной группы микроорганизмов использовали вырастающие на казеин-глицериновом (КГА) агаре актиномицеты, проводя дифференцированный учет колонии разных морфотипов. Для характеристики структуры комплексов определяли частоту встречаемости и долевое участие представителей отдельных секций и серий рода Streptomyces [1].
Морфологические признаки исследовали при помощи светового микроскопа Leica DM 2500 (Carl Zeiss, Германия). Принадлежность выделенных культур актиномицетов к роду Streptomyces определяли на основании характерных морфологических признаков: нефрагментированный мицелий, длинные цепочки спор – на воздушном и отсутствие спор – на субстратном мицелии.
Диагностические признаки природных изолятов стрептомицетов изучали на минеральном агаре 1, органическом агаре 11, глицерин-нитратной, овсяной и пептон-дрожжевой средах в соответствии с ключом Гаузе с соавт. [1].
Таксономическое положение выборочно отобранных штаммов (культуры с различными морфотипами колоний) определяли на основе анализа фрагментов 16S рРНК в НПК “Синтол” (г. Москва). Первичный сравнительный анализ полученных нуклеотидных последовательностей с последовательностями из базы данных GenBank проводили с помощью программы NCBI BLAST (http://www. n-cbi.nlm.nih.gov/blast). Парное выравнивание последовательностей осуществляли с помощью программы LALIGN (https://embnet.vital-it.ch/ software/LALIGN_form.html).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
За сорокалетний период использования дерново-подзолистой тяжелосуглинистой почвы в длительном полевом опыте выявлены существенные различия между вариантами с бессменными посевами зерновых культур, севооборотами с различным насыщением бобовыми травами, бессменным чистым паром и залежью. Так, залежная почва по агрохимическим характеристикам (по кислотности почвенного раствора, содержанию углерода и другим параметрам) стала соответствовать целинной [3]. В 2018 г. в ней отмечено характерное для дерново-подзолистых почв Предуралья содержание углерода – 1.36% (табл. 1). Наименьшие в опыте значения коэффициентов минерализации (0.37) и педотрофности (0.28), рассчитанные по данным учета численности прокариот, показывают, что в залежной почве процессы деструкции ОВ протекают гораздо медленнее, чем при других типах землепользования (табл. 2, рис. 1). Исходя из этих результатов, залежь можно охарактеризовать как климаксную экосистему, в которой процессы разложения и синтеза ОВ уравновешены и хорошо сбалансированы.
Таблица 1.
Агрохимические свойства дерново-подзолистой почвы
| Тип землепользования (фактор А) | Минеральные удобрения (фактор В) | Сорг, % | СГК/СФК | рНKCl | S | Hг | Са | Mg | Р2О5, мг/кг |
|---|---|---|---|---|---|---|---|---|---|
| смоль(экв)/кг | |||||||||
| Чистый пар бессменно | Без удобрений | 0.78 | 0.55 | 5.0 | 21.1 | 3.1 | 16.2 | 3.9 | 340 |
| Ячмень бессменно | 1.01 | 0.72 | 5.1 | 22.5 | 3.1 | 18.6 | 2.0 | 440 | |
| Озимая рожь бессменно | 0.95 | 0.70 | 5.1 | 19.8 | 3.2 | 18.4 | 2.7 | 294 | |
| Севооборот | |||||||||
| 0% бобовых | 1.07 | 0.72 | 5.2 | 20.1 | 3.2 | 17.9 | 3.6 | 420 | |
| 28.6% бобовых | 1.47 | 0.76 | 5.5 | 21.5 | 3.0 | 17.2 | 3.2 | 537 | |
| 42.9% бобовых | 1.21 | 0.78 | 4.9 | 21.7 | 3.5 | 18.2 | 2.9 | 314 | |
| Залежь | 1.36 | 0.96 | 4.8 | 19.8 | 4.8 | 17.2 | 2.8 | 473 | |
| Ячмень бессменно | NPK | 1.03 | 0.76 | 5.0 | 19.6 | 3.7 | 18.2 | 1.9 | 679 |
| Озимая рожь бессменно | 1.06 | 0.76 | 5.1 | 19.3 | 3.0 | 16.3 | 3.7 | 321 | |
| Севооборот | |||||||||
| 0% бобовых | 1.12 | 0.72 | 5.1 | 20.0 | 3.5 | 17.7 | 2.9 | 462 | |
| 28.6% бобовых | 1.56 | 0.78 | 5.3 | 21.0 | 3.2 | 17.9 | 2.7 | 732 | |
| 42.9% бобовых | 1.25 | 0.79 | 5.0 | 20.5 | 3.2 | 19.4 | 1.2 | 495 | |
| Главные эффекты | Фактора А | 0.06 | – | 0.1 | – | 0.3 | Fф < Fт | Fф < Fт | 70 |
| Фактора B и взаимодействия АВ | Fф < Fт | – | 0.1 | – | 0.2 | 1.1 | Fф < Fт | 53 | |
| Частные различия | I порядка | 0.10 | – | 0.1 | – | 0.4 | 4.5 | 4.6 | 99 |
| II порядка | 0.07 | – | 0.1 | – | 0.5 | 3.0 | 4.1 | 141 | |
Таблица 2.
Количество микроорганизмов, вырастающих на традиционных средах, в зависимости от способа хозяйственного использования почвы (среднее ± стандартное отклонение)
| Вариант | Общая численность, тыс./г почвы | ||
|---|---|---|---|
| МПА | КАА | ПА | |
| Чистый пар бессменно | 1733 ± 340 | 3467 ± 2721 | 2250 ± 989 |
| Ячмень бессменно | 4358 ± 860 | 3142 ± 807 | 3983 ± 2025 |
| без удобрений | |||
| NPK | 3883 ± 914 | 2842 ± 1124 | 3900 ± 1081 |
| Озимая рожь бессменно | 4800 ± 604 | 3892 ± 1408 | 6650 ± 1438 |
| без удобрений | |||
| NPK | 8300 ± 5166 | 5367 ± 4945 | 7417 ± 1459 |
| Севооборот (0% бобовых) | 6558 ± 2157 | 3700 ± 775 | 5417 ± 3032 |
| без удобрений | |||
| NPK | 4267 ± 1684 | 3500 ± 1341 | 8650 ± 1014 |
| Типичный семипольный севооборот (28.6% бобовых) | 10 542 ± 5851 | 5050 ± 1723 | 8483 ± 3540 |
| навоз | |||
| навоз + NPK | 5975 ± 1690 | 3733 ± 866 | 4117 ± 801 |
| Севооборот (42.9% бобовых) | 8792 ± 2716 | 3333 ± 2048 | 5150 ± 1716 |
| без удобрений | |||
| NPK | 10 592 ± 1320 | 3767 ± 1413 | 6950 ± 1858 |
| Залежь | 4308 ± 716 | 1717 ± 740 | 1200 ± 839 |
Рис. 1.
Коэффициенты минерализации (КАА/МПА) и педотрофности (ПА/МПА) по вариантам: 1 – чистый пар бессменно; 2, 8 – ячмень бессменно; 3, 9 – озимая рожь бессменно; 4, 10 – зерновой пятипольный севооборот (0% бобовых культур) (ячмень, рожь озимая, пшеница яровая, ячмень, овес); 5, 11 – полевой семипольный севооборот (типичный, 28.6% бобовых культур) (унавоженный чистый пар, рожь озимая, пшеница яровая с подсевом клевера, клевер 1 г. п., клевер 2 г. п., ячмень, овес); 6, 12 – полевой семипольный севооборот (42.9% бобовых культур) (сидеральный пар (клевер 1 г. п.), рожь озимая, пшеница яровая с подсевом клевера, клевер 1 г. п., клевер 2 г. п., ячмень, овес с подсевом клевера); 7 – залежь.
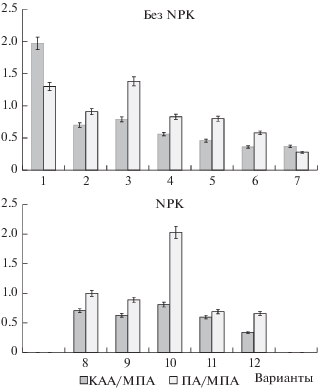
Минимальный уровень содержания углерода отмечен в бессменном чистом пару – 0.78%. Эта величина на 30% меньше, чем перед закладкой опыта. В условиях отсутствия растительности и связанного с ней поступления свежих растительных остатков, преобладали процессы разложения почвенного ОВ. Состояние микробной системы в длительно парующей почве типичное для поздних стадий микробных сукцессий: почти в два раза увеличилась численность бактерий, утилизирующих минеральные формы азота, возросло количество олиготрофных бактерий и актиномицетов, участвующих в разложении наиболее трудногидролизуемых органических соединений. По сравнению с залежью коэффициент минерализации в бессменном чистом пару выше в 5.3 раза, а коэффициент педотрофности – в 4.6 раза. Это свидетельствуют об интенсивных процессах разложения ОВ в пару. Уровень углерода, установившийся за 41 год в бессменном пару, условно принят за минимальное содержание углерода в дерново-подзолистой тяжелосуглинистой почве Предуралья [4].
Бессменное возделывание ячменя и озимой ржи без удобрений и при внесении N60P30K60 способствовало поддержанию агрохимических параметров почвы на уровне, близком к исходному. Пожнивные и корневые остатки, прижизненная корневая экссудация зерновых культур способствовали поддержанию численности микроорганизмов, участвующих в цикле углерода. Но почва в посевах зерновых культур, очевидно, вследствие дефицитного поступления азота, характеризовалась невысокими значениями коэффициентов минерализации (0.70–0.79) и близкими к единице индексами педотрофности (0.91–1.38).
В отличие от яровой культуры (ячменя), бессменное выращивание озимой ржи, даже без удобрений, привело к формированию в почве большего пула микроорганизмов. Вероятно, это связано с тем, что биомасса корневых и пожнивных остатков озимой культуры превосходит таковую для яровой культуры. На фоне внесения под озимую рожь минеральных удобрений коэффициенты минерализации (0.63) и педотрофности (0.89) имели более близкие значения, чем в вариантах без удобрений (0.79 и 1.38 соответственно), что говорит об оптимизирующем влиянии минеральных удобрений на процессы трансформации ОВ в посеве озимой ржи.
В севообороте без бобовых трав и минеральных удобрений коэффициенты минерализации и педотрофности по величине были сопоставимы со значениями в вариантах с посевами зерновых культур бессменно. На фоне NPK численность микроорганизмов, вырастающих на МПА и КАА, практически не изменилась, тогда как численность учтенных на ПА бактерий олиготрофного блока, довольствующихся незначительным количеством питательных веществ и завершающих сложный процесс разложения ОВ увеличилась в пределах порядка. В результате индекс педотрофности в этом варианте возрос до максимального в опыте значения – 2.03, а коэффициент минерализации составил 0.81.
Возделывание сельскохозяйственных культур в севообороте с долей бобовых 42.9% без внесения органических удобрений обеспечило стабилизацию исходного уровня ОВ в почве, не ухудшая ее другие агрохимические свойства. Бобовые травы в севообороте являются центрами формирования бобово-ризобиального комплекса, который обеспечивает биологическим азотом практически все микробное население почвы. Коэффициенты минерализации в почве севооборотов, включающих 42.9% бобовых (0.34 и 0.36), имели низкие значения, сопоставимые с коэффициентом минерализации залежной почвы (0.37), а в почве севооборотов с меньшим насыщением (28.6%) бобовыми травами увеличивались до 0.46 (при внесении навоза) и до 0.60 (при внесении навоза + NPK). Ведение традиционного севооборота с двумя полями клевера, применением органической и органо-минеральной систем удобрения привело к улучшению комплекса агрохимических параметров дерново-подзолистой почвы. Кислотность почвы изменилась от кислой (рНKCl 4.8) до слабокислой (рНKCl 5.3–5.4), содержание Сорг увеличилось от 1.10 до 1.47–1.56%, фосфора – до 537.5–732.5 мг/кг.
Приведенные результаты показывают, что стабилизация ОВ дерново-подзолистой почвы зависит от типа использования и снижается в ряду: залежь > > севообороты (0–28.6–42.9% бобовых) > бессменный посев зерновых > бессменный чистый пар. При сопоставлении полученных в опыте агрохимических данных с результатами микробиологических исследований обнаружена обратная зависимость между содержанием в почве Cорг и коэффициентами минерализации (r = –0.67; p < 0.01). Отрицательная корреляция выявлена также между соотношением гуминовых и фульвокислот (СГК/СФК) в составе ОВ почвы и коэффициентами ее педотрофности (r = –0.64; p < 0.02). Помимо количественных различий в бессменно парующей и возделываемых почвах произошли качественные изменения в составе ОВ: увеличилась его лабильность, о чем свидетельствуют меньшие (0.55–0.79), чем в залежной почве (0.96), соотношения СГК/СФК. Между показателями СГК/СФК и коэффициентами минерализации в почве длительного стационара установлена наиболее тесная отрицательная корреляция (r = –0.81; p < 0.001). Таким образом, по соотношению в почве микроорганизмов, вырастающих при посеве на средах с различными источниками азота – минеральными (КАА), органическими (МПА) и теми и другими одновременно, взятыми в минимальных концентрациях (ПА). Были рассчитаны экологические коэффициенты минерализации и педотрофности, величины которых тесно коррелируют с показателями гумусности почвы. Это не только позволяет судить о направленности микробных процессов трансформации ОВ дерново-подзолистой почвы за сорокалетний период ее различного использования, но и прогнозировать дальнейшую динамику содержания ОВ при различных типах землепользования.
Для выявления структурных перестроек микробного сообщества в результате различного использования дерново-подзолистой тяжелосуглинистой почвы в качестве модельной группы использовали актиномицеты – мицелиальные бактерии, которые играют ключевую роль в поддержании почвенного гомеостаза [15].
Дифференцированный учет в посевах на КГА колоний по морфотипам позволил установить, что во всех вариантах опыта на дерново-подзолистой почве преобладали колонии, микроскопия которых выявила типичные для рода Streptomyces морфологические признаки.
Выборочная идентификация выделенных штаммов, основанная на анализе фрагментов гена 16S рРНК, подтвердила, что изоляты с данным морфотипом являются представителями рода Streptomyces, семейства Streptomycetaceae, порядка Streptomycetales, класса Actinobacteria. В то же время видовая идентификация стрептомицетов, основанная на анализе гена 16s рРНК, была затруднена из-за высокого сходства нуклеотидных последовательностей данного гена для представителей всех таксонов внутри семейства Streptomycetaceae [8]. Действительно, все нуклеотидные фрагменты, выданные BLAST, для исследуемых штаммов более, чем на 98% соответствовали введенным последовательностям. В связи с этим помимо генетических данных в работе были изучены такие фенотипические признаки изолятов, как цвет воздушного и субстратного мицелия на диагностических средах. Это позволило в образцах почвы из различных вариантов опыта определить частоту встречаемости и долевое участие представителей отдельных цветовых секций и серий.
Анализ структуры комплексов актиномицетов в почвах сравниваемых вариантов показал, что залежную и бессменно парующую почву от других отличает узкий спектр выделяемых таксонов. С высокой частотой встречались виды серии Clnereus Achromogenes и секции Imperfectus, доминирующие в большинстве и других вариантов опыта. В парующей почве отмечены случайные представители Clnereus Violaceus и секции Albus (рис. 2). В бессменных посевах ячменя и озимой ржи видовой спектр актиномицетов расширился за счет видов из секций и серий Clnereus Chromogenes, Clnereus Aureus, Albus, выявляемых в различных соотношениях в зависимости от вида культуры (яровая или озимая) и внесения NPK. Сходный видовой состав имели комплексы актиномицетов в вариантах с севооборотами без включения бобовых трав, отличаясь лишь более значительной долей участия пигментированных видов серии Clnereus Chromogenes.
Рис. 2.
Долевое участие (%) в комплексах актиномицетов дерново-подзолистой почвы при разных способах ее использования видов: 1 – Clnereus Achromogenes, 2 – Clnoreus Chromogenes, 3 – Clnereus Aureus, 4 – Clnereus Violaceus, 5 – Albus, 6 – Helvolus, 7 – Imperfectus.
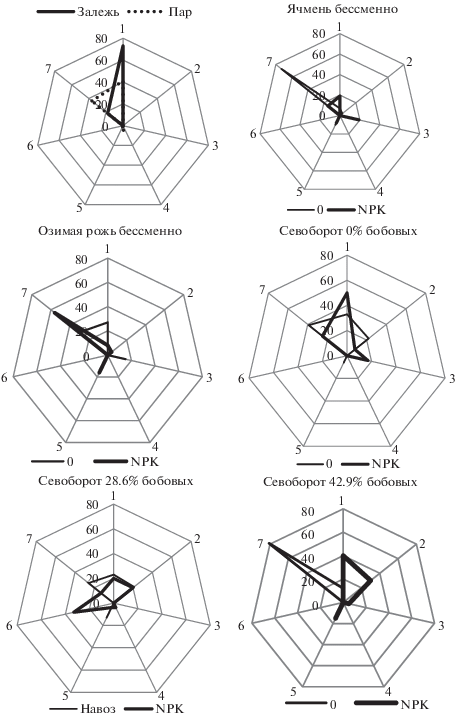
Насыщение севооборота на 28.6% бобовыми травами, при одновременном внесении в почву навоза отдельно и в сочетании с NPK, привело к формированию актиномицетных комплексов, в структуре которых все выявляемые секции и серии были представлены относительно равномерно, без выделения явных доминантов. Актиномицетный комплекс в почве севооборота с увеличенной до 42.9% долей бобовых трав, напротив, характеризовался сильной концентрацией доминирования. В его структуре на долю видов секции Imperfectus приходилось более 80% всего актиномицетного разнообразия. Внесение NPK в этом же варианте расширило видовой состав от двух до четырех различных секций и серий, при этом в комплексе произошла элиминация представителей секции Imperfectus. Таким образом, различный характер использования дерново-подзолистой почвы нашел отражение в специфической структуре комплекса почвенных актиномицетов по вариантам длительного полевого стационара.
ЗАКЛЮЧЕНИЕ
Различным типам землепользования соответствуют различные структуры сообществ почвенных микроорганизмов. Бессменные посевы зерновых культур, севообороты с различным насыщением бобовыми травами, бессменные чистый пар и залежь, внесение минеральных удобрений и навоза сопровождались структурными перестройками в комплексе мицелиальных прокариот, заключавшимися в изменении количества выявляемых секций и серий рода Streptomyces, частоте встречаемости отдельных представителей, их долевого участия в комплексе, состава доминантов. Главным фактором, ответственным за выявленные различия в структуре микробных комплексов, является, по-видимому, содержание органического углерода в почве и его качество. Механическая обработка почвы сама по себе и возделывание на ней сельскохозяйственных культур ведут к повышению интенсивности микробных процессов трансформации ОВ, что выразилось в увеличении коэффициентов минерализации и педотрофности почвы. Отказ от земледелия (залежь) привел к сдвигу в структуре микробного сообщества, первичным эффектом которого явилось снижение коэффициентов минерализации и педотрофности. Изменения структуры комплексов актиномицетов, взятых в качестве модельной группы почвенных микроорганизмов, согласуются с показателями численности других почвенных микроорганизмов, учитываемых на традиционных средах, и величинами коэффициентов минерализации и педотрофности по вариантам опыта.
Список литературы
Гаузе Г.Ф., Преображенская Т.П., Свешникова М.А., Терехова Л.П., Максимова Т.С. Определитель актиномицетов. Роды Streptomyces, Streptoverticillium, Chainia. М.: Наука, 1983. 248 с.
Добровольский Г.В., Куст Г.С., Чернов И.Ю., Добровольская Т.Г., Лысак Л.В. и др. Почвы в биосфере и жизни человека. М., 2012. 584 с.
Завьялова Н.Е. Органическое вещество дерново-подзолистых почв Предуралья. Пермь, 2014. 328 с.
Наумкин В.Н. Биологизация и экологизация земледелия Юго-Запада России // Агро XXI. 2001. № 3. С. 20–21.
Семенов В.М., Когут Б.М. Почвенное органическое вещество. М.: ГЕОС, 2015. 233 с.
Титова В.И., Козлов А.В. Методы оценки функционирования микробоценоза почвы, участвующего в трансформации органического вещества. Нижний Новгород: Нижегородская с.-х. акад., 2012. 64 с.
Ковалевская Н.П., Завьялова Н.Е., Шаравин Д.Ю., Фомин Д.С. Биологическая активность дерново-подзолистой почвы в длительном опыте с различными агротехническими приемами // Российская сельскохозяйственная наука. 2019. № 3. С. 38–41. https://doi.org/10.31857/S2500-26272019338-41
Labeda D.P., Goodfellow M., Brown R., Ward A.C., Lanoot B., Vanncanneyt M., Tamura T. Phylogenetic study of the speCles within the family Streptomycetaceae // Anton. Leeuw. Int. J. G. 2012. V. 1. 101. P. 73–104. https://doi.org/10.1007/s10482-011-9656-0
Lehmann J., Kleber M. The contentious nature of soil organic matter // Nature. 2015. V. 528. P. 60–68. https://doi.org/10.1038/nature16069
Manzoni S., Taylor P., Richter A., Porporato A., Ågren G.I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils // New Phytol. 2012. V. 196. P. 79–91. https://doi.org/10.1111/j.1469-8137.2012.04225.x
Martí-Roura M., Hagedorn F., Rovira P., Romanyà J. Effect of land use and carbonates on organic matter stabilization and microbial communities in Mediterranean soils // Geoderma. 2019. V. 351. P. 103–115. https://doi.org/10.1016/j.geoderma.2019.05.021
Miltner A., Bombach P., Schmidt-Brücken B., Kästner M. SOM genesis: microbial biomass as a significant source // Biogeochemistry. 2012. V. 111. P. 41–55. https://doi.org/10.1007/s10533-011-9658-z
Schmidt M.W.I., Torn M.S., Abiven S., Dittmar T., Guggenberger G., Janssens I.A., Kleber M., Kögel-Knabner I. et al. Persistence of soil organic matter as an ecosystem property // Nature. 2011. V. 478. № 7367. P. 49–56. https://doi.org/10.1038/nature10386
Soares M., Rousk J. Microbial growth and carbon use efficiency in soil: Links to fungal-bacteria dominance, SOC-quality and stoichiometry // Soil Biol. Biochem. 2019. V. 131. P. 195–205. https://doi.org/10.1016/j.soilbio.2019.01.010
Strohl W.R. Antimicrobials // Microbial Diversity and Bioprocessing / Ed. A.T. Bull. Washington DC: American Society for Microbiology, 2004. P. 336–355.
Zornoza C.G., Mataix-Solera J., Scow K.M., Arcenegui V.V., Mataix-Beneyto J. Changes in soil microbial community structure following the abandonment of agricultural terraces in mountainous areas of Eastern Spain // Appl. Soil Ecol. 2009. V. 42 P. 315–323. https://doi.org/10.1016/j.apsoil.2009.05.011
Дополнительные материалы отсутствуют.