

Почвоведение, 2021, № 4, стр. 465-474
Современные пойменно-старичные отложения среднего течения р. Камчатки (результаты комплексного группового биологического анализа)
О. Н. Успенская a, *, М. М. Певзнер b, Т. Д. Каримов b, c
a Всероссийский научно-исследовательский институт овощеводства
140153 Московская область, Раменский район, Верея, 500, Россия
b Геологический институт РАН
119017 Москва, Пыжевский пер., 7, Россия
c МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: usp-olga@yandex.ru
Поступила в редакцию 27.05.2020
После доработки 16.06.2020
Принята к публикации 27.07.2020
Аннотация
Впервые методом комплексного группового биоанализа проведено сравнительное исследование современных отложений весеннего паводка (пойменная фация аллювия) и начальной стадии зарастания старичного озера (старичная фация аллювия) в среднем течении р. Камчатки. Установлены различия процентного соотношения разных форм диатомовых водорослей и микроостатков вегетативных органов высших растений. Сравнение диатомового комплекса изученных проб с аналогичными данными по крупным озерам Камчатки позволило выявить принципиальные отличия в его составе, в том числе установлены два рода диатомей (Cyclotella и Stephanodiscus), которые либо не встречаются, либо присутствуют в незначительных количествах в отложениях паводка и начальной стадии зарастания старичного озера. Отложения полых вод, характеризующиеся по данным биоанализа как “водоем”, по составу захороненных биоостатков принципиально отличаются от таковых для долгоживущего крупного озера. Полученные результаты могут быть использованы для идентификации погребенных пойменно-старичных отложений в Центральной Камчатке.
ВВЕДЕНИЕ
Водные организмы (водоросли, беспозвоночные животные) и водно-болотные высшие растения имеют широкое распространение, а их остатки хорошо сохраняются в отложениях водоемов. Их можно встретить даже в самых малых биотопах, которые формируются в пределах пойменно-руслового комплекса рек. Являясь непосредственно связанными с водной средой, эти организмы представляют собой надежные индикаторы типа и характера осадконакопления, а также позволяют лучше распознать генезис некоторых осадочных толщ.
В последние годы были изучены и датированы позднеплейстоценовые отложения, вскрывающиеся в высоких береговых обрывах (ярах) р. Камчатки в ее среднем течении [16, 18, 19]. Новые данные изотопного (радиоуглеродного и уран-ториевого) датирования свидетельствуют о позднеплейстоценовом возрасте осадков. Ранее эти отложения рассматривались как опорные для среднего плейстоцена [3, 10]. Полевые наблюдения и первые результаты палеоботанического анализа заставляют усомниться и в ранее установленном генезисе некоторых толщ, слагающих яры. Для идентификации отложений и условий их образования применяли метод комплексного группового биологического анализа. Однако столкнулись с практически полным отсутствием данных о составе биоостатков в современных отложениях пойм и стариц в долине р. Камчатки. В связи с этим анализировали представительные пробы пойменного наилка и отложений зарастающей старицы, чтобы использовать этот сравнительный материал для определения генезиса пойменно-старичных отложений позднеплейстоцен-голоценового возраста.
Цель работы – детальное изучение современного пойменно-старичного аллювия р. Камчатки в ее среднем течении и определение ассоциаций водных организмов и высших растений для разных типов отложений.
ОБЪЕКТЫ И МЕТОДЫ
Для определения условий, в которых образовываются современные аллювиальные отложения в среднем течении р. Камчатки применяли комплексный групповой11 количественный биологический анализ (далее биоанализ) по [8, 20, 32], в частности, анализ диатомовых водорослей.
Метод комплексного группового количественного анализа предусматривает определение биоостатков в естественном материале водных осадков, так как любая температурная или химическая обработка пробы может уничтожить остатки некоторых из перечисленных выше групп организмов. Определение ведется до возможной степени детальности. Выводы основываются на количественных соотношениях групп разной экологии. Видовой анализ дополняет результаты группового.
Известно, что разные роды, виды и разновидности диатомовых водорослей имеют широкий спектр требований к окружающей среде. Среди них есть планктонные и донные, теплолюбивые и холодноводные, ацидофилы и алкалифилы, галофобы и галофилы, олигосапробы и мезосапробы и т. д. В то же время отдел (группа) диатомей в целом характерен для крупных, проточных, хорошо аэрируемых, умеренно холодных или теплых водоемов средней или высокой минерализации, с хорошо выраженным процессом нитрификации, с достаточным количеством силикатов, фосфатов и других биогенных элементов в воде. Поэтому массовое присутствие диатомей в донных осадках должно отвечать, как правило, периодам усиления обводненности водоема, когда он обеспечивается постоянным подтоком биогенных элементов, лучше аэрируется, становится чище благодаря усиливающемуся процессу разложения органического вещества.
Материалы настоящей публикации базируются, в первую очередь, на анализе некоторых жизненных форм диатомей, которые маркируют те или иные экологические обстановки. Бентосные формы приспособлены к существованию в осадках на дне водоемов. Доминирование донных диатомей может свидетельствовать о мелководных условиях образования осадков. Представители группы обрастателей ведут прикрепленный образ жизни на растениях, камнях и другом субстрате. Доминирование этой группы также может свидетельствовать о мелководности водоема, в котором отлагаются осадки, а также о наличии в нем водной или водно-болотной растительности. Реофильные диатомеи, входящие в группу обрастателей, способны вегетировать в быстротекущих водах и, следовательно, характеризуют речные отложения и свидетельствуют о режиме проточности. Планктонные формы обитают в толще воды и характеризуют глубоководность водоема.
Большинство золотистых водорослей – типично планктонные литоральные организмы, олигогалобы. В основном они встречаются в чистых прохладных водах, предпочитая зоны с гуминовым подтоком и чаще всего маркируют отложения верховых болот. Они свидетельствуют о слабой минерализации воды, о некотором обеднении ее биогенными элементами и железом, на возможную заболоченность водоема. В периоды заболачивания количество золотистых водорослей возрастает, а диатомовых – уменьшается.
Губки встречаются в водоемах разных типов, но основным фактором, определяющим возможность их обитания, является хорошая аэрация и отсутствие в воде взвешенных частиц, забивающих их циркуляционную систему. Поэтому наличие в осадках остатков губок указывает на проточность воды, но слабую, без взмучивания донных осадков.
Методика биоанализа заключалась в разведении 0.5–1.0 см3 пробы водой в соотношении 1 : 50 и просмотре капель взвеси при помощи светового микроскопа Микмед-5 при увеличении в 280–400 раз. Подсчитывали не менее 500 остатков всех отделов водорослей (диатомовые, синезеленые, золотистые, эвгленовые, желтозеленые, пирофитовые, харовые, хлорококковые, десмидиевые, зеленые нитчатые, вольвоксовые), животных (ветвистоусые, ракушковые рачки, простейшие, насекомые), вегетативных частей высших растений, пыльцы и спор высших растений. Затем рассчитывали процентные соотношения между группами организмов. Результаты анализа оформляли в виде таблиц и диаграмм.
Для определения состава высших растений усредненную пробу 20–30 г (для высокозольных образцов до 100 г и более) промывали на сите с диаметром ячеек 0.25 мм под струей воды с одновременным отмучиванием органической составляющей (сухую пробу предварительно замачивали горячей водой на сутки). Надситовый остаток помещали в стеклянную тару и заливали 2–5%-ной щелочью на несколько часов. Затем пробу снова промывали на сите с диаметром ячеек 0.25 мм с повторным отмучиванием органической составляющей. Освобожденный от гумусовых и минеральных компонентов растительный материал просматривали под микроскопом при увеличении в 56–80 раз. Процентные соотношения между видами растений устанавливали глазомерно в зависимости от площади, занимаемой остатками вегетативных частей данного вида в поле зрения микроскопа.
Идентификацию диатомовых водорослей проводили, согласно работам [28–31]. В более ранних публикациях, таких как [3], использовалась иная номенклатура диатомовых водорослей; например, Melosira italica var. valida (Grun.) = Melosira italica subsp. subarctica O. Müller 1906 = Aulacoseira subarctica (O. Müller) E.Y. Haworth [23].
РЕЗУЛЬТАТЫ ПРЕДШЕСТВУЮЩИХ ИССЛЕДОВАНИЙ
Ранее для Камчатки метод комплексного группового анализа применяли только для изучения голоценовых отложений некоторых торфяных болот [24].
Диатомовая флора в отложениях современных и голоценовых озер Камчатки хорошо изучена [1, 2, 4–7, 9, 11–15, 17, 21, 25–27]. Установлено, что для водоемов и водотоков Камчатки наиболее характерны такие виды, как Aulacoseira subarctica (O. Muller) Haworth, Stephanodiscus alpinus Hustedt, S. minutulus (Kutzing) Cleve et Muller. Эти виды широко распространены в регионе и характеризуются высокими показателями встречаемости и биомассы в пробах [9].
В фитопланктоне большинства вулканических озер Камчатки в значительном количестве (до 13% от общего количества диатомовых) были обнаружены Cyclotella ocellata Pant. [26], а также вновь выделенный вид Stephanodiscus popovskayae Genkal et Lepskaya [4]. Примечательно, что C. ocellata обитает в крупных глубоководных озерах (Ладожское, Байкал, Эльгыгытгын, Хубсугул), а также в средних по площади холодноводных озерах Камчатки (оз. Паланское) [4].
Для голоценовых осадков оз. Двухюрточного, наиболее распространенными видами являются: Stephanodiscus minutulus, Aulacoseira subarctica, Stephanodiscus medius Hakansson, Cyclotella ocellata и Stephanodiscus alpinus. Для наиболее молодых осадков характерны некоторые особенности: резкое увеличение содержания Stephanodiscus minutulus при все еще частой встречаемости Aulacoseira subarctica [25–27].
Принципиально другие виды характерны для оз. Нерпичьего, имеющего связь с Тихим океаном. Установлено большое разнообразие видов, как пресноводных, так и солоноватоводных. Однако доминирующую группу составляют солоноватоводные представители рода Melosira Ag. [21].
Таким образом, можно говорить о том, что при большом разнообразии форм, доминирующими видами диатомей для пресноводных озерных водоемов Камчатки являются планктонные Aulacoseira subarctica, Stephanodiscus alpinus и S. minutulus. Появление в осадках Cyclotella ocellata может свидетельствовать о значительных по площади и глубине холодноводных озерах.
Анализ современной диатомовой флоры долины р. Камчатки проводили в 60-е годы прошлого века [3]. Изучали планктонные пробы в окрестностях поселков Кирганик, Козыревск и Ключи. Было установлено, что подавляющее большинство форм – олигогалобы и принадлежат к категории индифферентов (80%), на долю галофилов приходится около 15%, а галофобы не превышают 5%. В группу индифферентов входит большая часть массовых и наиболее типичных видов. По географическому распространению большинство представителей современной аллювиальной диатомовой флоры космополиты (69%), северо-бореальные составляют около 17%, арктобореальные – 6%, северо-альпийские – 8%. При этом наиболее характерными представителями планктона р. Камчатки в летне-осенний период являются диатомовые водоросли рода Melosira Ag. Среди донных литоральных форм грунтов и обрастаний обычны Diatoma hiemale (Lyngb.) Heib. и Meridion circulare Ag.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В процессе изучения позднеплейстоценовых отложений, слагающих яры р. Камчатки и ее притоков, отмечено, что для наиболее тонких в гранулометрическом отношении осадков (тонкозернистые пески, суглинки, алевриты) крайне трудно установить условия их формирования. Поэтому для получения сравнительного материала анализировали современные отложения. Для идентификации наиболее проблематичных осадков (отложений паводка и начальной стадии зарастания старичного озера) отбирали контрольные пробы современных отложений вблизи яров Генералка и Большой (рис. 1).
Рис. 1.
Отложения паводка: А – положение точек опробования на карте Камчатки; Б – яр Генералка, стрелка показывает место отбора пробы; В – место отбора пробы Г, см. табл. 1 (Б, В – фото М.М. Певзнер); Г – весенний паводок в пос. Долиновка, фото из интернета А. Галдиной.

Отложения паводка. На берегу р. Камчатки около яра Генералка на поверхности низкой поймы, частично заросшей ольшаником с хвощем, в 0.6 м над урезом воды отобрали пробу влажного наноса, оставшегося после весеннего паводка. Материал пробы представлен опесчаненым суглинком коричневато-серого цвета со значительной примесью мелкой древесной трухи. По результатам биоанализа в пробе резко доминируют диатомовые водоросли (табл. 1, проба Г). Остатки высших растений крайне немногочисленны и представлены, главным образом, древесным детритом (кора и древесина лиственных).
Таблица 1.
Результаты комплексного группового биоанализа современных отложений паводка и начальной стадии зарастания старичного озера в среднем течении р. Камчатки
| Отложения паводка | % |
|---|---|
| Проба Б. Старичные отложения у яра Большого | |
| Высшие растения: травянистые (фрагменты корней) (80%), Cyperaceae (10%), Equisetum (10%), кора хвойных (ед.) |
10.5 |
| Золотистые водоросли (Chrysophyta) | 1.4 |
| Диатомовые водоросли: планктонные 44.0%: Aulacoseira subarctica (O. Mull.) Haworth (37.5%), A. varians Ag. (3.6%), A. italica (Ehr.) Simons. (1.6%), Cyclotella Kutz. (1.2%) обрастатели 35.6%: Fragilaria Lyndb. (10.4%), Denticula elegans Kutz. (5.2%), Synedra Ehr. (3.6%), Rhopalodia O. Mull. (2.8%), Gomphonema Ag. (2.8%), Epithemia Breb. (2.8%), Meridion circulare Ag. (2.0%), Eunotia Ehr., Rhoicosphenia curvata (Kutz.) Grun., Tabellaria Ehr., Tetracyclus rupestris (A. Br.) Grun., Cymbella Ag. донные 20.4%: Navicula Bory (10.0%), Pinnularia Ehr. (2.0%), Amphora Ehr. (2.0%) и др. (Cymatopleura W. Sm., Caloneis Cl., Gyrosigma Hass.) реофильные 2.0%: Meridion circulare Ag. |
87.7 |
| Spongia | 0.4 |
| Обстановка накопления: современная субаэральная растительность возникла на отложениях довольно глубокого, холодного, слабопроточного озера с обширной акваторией | |
| Проба Г. Отложения современной поймы у Яра Генералка | |
| Высшие растения: кора и древесина лиственных (100%), Sphagnum (ед.) |
14.0 |
| Золотистые водоросли (Chrysophyta) | 1.6 |
| Диатомовые водоросли: обрастатели 44.7%: Ceratoneis arcus (Ehr.) Kutz. (13.6%), Fragilaria Lyndb. (9.7%), Gomphonema Ag. (6.8%), Synedra Ehr. (3.9%), Meridion circulare Ag. (3.9%) и другие (Eunotia Ehr., Cocconeis Ehr., Tetracyclus Ralfs, Didymosphenia geminate (Lyndb.) M. Schmidt) донные 31.1%: Navicula Bory (11.7%), Amphora Ehr. (11.7%), Pinnularia Ehr. (4.9%), а также Hantzshia Grun., Caloneis Cl. планктонные литоральные и пелагические 12.6%: Fragilaria capucina Desm. (7.8%), Aulacosira subarctica (O. Mull.) Haworth (3.9%), Cyclotella Kutz. (1.0%) реофильные 17.5%: Ceratoneis arcus (Her.) Kutz., Meridion circulare Ag. |
84.4 |
| Обстановка накопления: проба сформирована в относительно неглубоком проточном холодноводном водоеме или это литоральные отложения проточного холодноводного водоема | |
Отложения идентифицируются как водные, поскольку 86.0% биоостатков в пробе составляют водоросли, главным образом, диатомовые (84.4%). О сравнительно небольшой глубине водоема свидетельствует преобладание в пробе диатомей-обрастателей (44.7% от суммы диатомей) и донных видов (31.1%), которые являются обитателями неглубоких водоемов (или литоральных зон глубоких). Проточность водоема определяется тем, что численность реофильных видов (Ceratoneis arcus (Ehr.) Kutz., Meridion circulare Ag.) составляет 17.5% от суммы всех диатомей и 39% от группы обрастателей (рис. 2а, круговая диаграмма). Вода, по-видимому, была холодной, так как численность холодолюбивых видов (Aulacoseira subarctica (O. Mull.) Haworth, Ceratoneis arcus (Ehr.) Kutz., Meridion circulare Ag., Tetracyclus Ralfs и Didymosphenia geminate (Lyndb.) M. Schmidt) составляет 22% от суммы диатомей. Остатки вегетативных органов высших растений составляют 14.0% от суммы всех биоостатков и представлены фрагментами коры и древесины березы (Betula sp.), единично – листочками сфагнового мха (Sphagnum sp.).
Рис. 2.
Гистограммы процентного содержания диатомовых водорослей и остатков высших растений в пробах из отложений паводка (А) и старичного озера (Б), согласно данных табл. 1, а также озера Двухюрточного по [26] (В). На круговых диаграммах показано процентное соотношение реофильных диатомей (светлый тон) и прочих обрастателей (темный тон) для отложений паводка (А) и старичного озера (Б): 1 – высшие растения; 2 – золотистые водоросли; 3‒5 – диатомовые водоросли: 3 – планктонные, 4 – обрастатели, 5 – донные; 6 – губки.
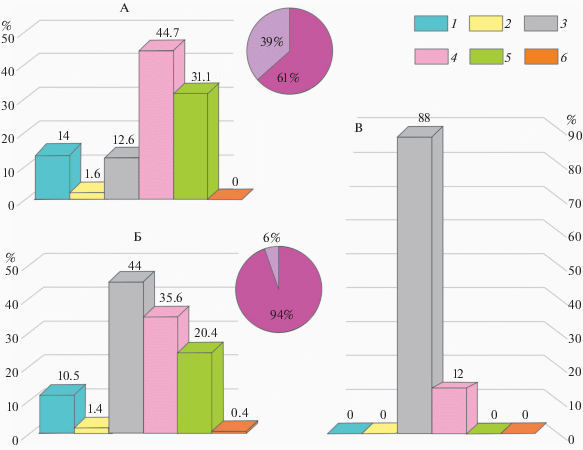
Можно предполагать, что проба, содержащая вышеописанные биоостатки, была сформирована в небольшом относительно неглубоком проточном холодноводном водоеме; либо в литорали более обширного проточного холодноводного водоема.
Поскольку анализировали материал весеннего паводка, оставшийся на берегу реки через 1–1.5 мес. после спада воды, то в данном случае понятие “водоем” характеризует не самостоятельно существующее озеро, но временный сезонный разлив реки (рис. 1, Г). Такие разливы могут существовать в среднем течении р. Камчатки до двух месяцев, когда образуются обширные мелководные озера, в которых, по мере падения уровня воды, начинается седиментация осадка. При этом полые воды, образованные при снеготаянии, характеризуются низкими температурами и высокой мутностью. Первые два месяца после вскрытия льда и разлива полых вод характеризуют раннюю (весеннюю) стадию развития фитопланктонного (диатомового, в частности) комплекса. За вегетационный период сменяется несколько поколений водорослей, поэтому весенние условия успевают отразиться и сохраниться (законсервироваться) в видовом составе изучаемых отложений. В этот период вода еще не прогрелась, относительно бедна кислородом и наименее прозрачная за весь вегетационный период. В результате формируется определенный набор видов, характерных для холодноводных неглубоководных озер, богатых биофильными элементами. При этом глубина изученного “водоема”, судя по высотному уровню поломанных и испачканных паводком деревьев, не превышала 2 м.
Отложения старичного озера. На берегу большого старичного озера, расположенного в частично закрытой излучине (имеет связь с рекой в половодье) у яра Большого (рис. 3) взята проба корнеобитаемого слоя. Отбор проводили в 1.5 м над урезом озера, в 0.3 м от границы воды, на глубине 15–20 см от дневной поверхности. Проба представляла собой сизо-черные полужидкие пески тонкозернистой размерности с характерным болотным запахом. После высыхания и растирания в пальцах пески приобретали голубовато-сизый цвет. Корнеобитаемый слой подстилался 25-сантиметровым горизонтом чуть более грубых нестратифицированных влажных песков темно-бежевого цвета, которые уходили под урез озера. Пробу отбирали у подножия осыпи на поверхности, заросшей хвощем, при этом на противоположном низком берегу озера преобладают осоки.
Рис. 3.
Старичное озеро у яра Большого: А – фрагмент космического снимка из Google Earth, видно, что вход в излучину уже закрыт, точкой указано место съемки фото В; Б – место отбора пробы Б; В – излучина превратилась в крупное озеро старичного типа, вид на юго-восток; Г – низкий берег активно зарастает водно-болотной растительностью (осока, белокрыльник). Фото М.М. Певзнер.

Биоанализ показал значительное доминирование диатомовых водорослей (табл. 1, проба Б). Среди высших растений преобладают травянистые, присутствуют осоки и хвощ. Кроме того, в незначительном количестве обнаружены спикулы губок. Отложения интерпретируются как водные, поскольку 89.1% биоостатков составляют водоросли, в основном, диатомовые (87.7%). Материнский водоем (озеро) должен быть довольно глубоким и обширным по площади, так как 44.0% от суммы диатомей приходится на планктонные виды (Aulacoseira subarctica (O. Mull.) Haworth, A. varians Ag., A. italica (Ehrenb.) Simons., Cyclotella Kutz.). При этом обрастатели (Fragilaria Lyndb., Denticula elegans Kutz., Synedra Ehr. и др.) и донные (Navicula Bory, Pinnularia Ehr., Amphora Ehr. и др.) занимают подчиненное положение, составляя 35.6 и 20.4% от суммы диатомей соответственно.
Проточность озера очень слабая: реофильные формы организмов (Meridion circulare Ag., Spongia) cоставляют всего 2.1% от суммы всех найденных биоостатков и 6% от всех обрастателей (рис. 2, Б, круговая диаграмма). Следовательно, для развития такой многочисленной и разнообразной диатомовой флоры (табл. 1) требуется обширная акватория, чтобы ветровое перемешивание воды обеспечивало для диатомей достаточные аэрацию и питание. Озеро холодное, что подтверждается численностью холодолюбивых видов, составляющей около половины всех диатомей. При этом заметно преобладает Aulacoseira subarctica (O. Mull.) Haworth (36.0% от суммы диатомей), представлены и другие виды холодолюбы: Meridion circulare Ag., Denticula elegans Kutz., Tetracyclus rupestris (A. Br.) Grun., Diatoma hiemale (Lyndb.) Heib. Лишь 10.5% биоостатков пробы приходится на фрагменты вегетативных органов высших растений. Доминируют корешки травянистых растений (80% от всех высших растений), также установлены хвощ (E-quisetum sp.) и осока (Carex sp.). Единично встретилась хвойная кора.
Данная ассоциация была идентифицирована как водные осадки с остатками высших растений, характерными для травяного луга. Можно сделать вывод, что современная субаэральная луговая растительность возникла на субстрате, который представляет собой продукт глубокого обширного водоема с холодной, слабо проточной водой. Действительно, в частично закрытой излучине у яра Большого сформировалось крупное озеро старичного типа (рис. 3, В). Оно глубокое и большую часть года застойное, в наши дни связь излучины с основным руслом реки осуществляется только в половодье через “нижнюю” протоку, так как вход в излучину давно зарос (рис. 3, А).
Таким образом, и в случае отложения паводка, и в старичных условиях осадки могут идентифицироваться как отложения водоемов со слабопроточной холодной водой. Обилие холодолюбивых организмов, по-видимому, обусловлено тем, что р. Камчатка питается притоками, истоки которых находятся в горах, а паводок в значительной степени связан с процессом снеготаяния. Для крупных стариц, таких как озеро в излучине у яра Большого, анализ диатомовых может свидетельствовать и о значительных глубинах водоема (например, до 10 м).
В части видового состава результаты наших исследований диатомовой флоры в современных пробах близко согласуются с данными [3], полученными по современным аллювиальным отложениям р. Камчатки.
ОБСУЖДЕНИЕ МАТЕРИАЛОВ
На гистограммах (рис. 2), построенных по данным табл. 1, показано, что в обоих случаях в комплексном составе проб доминируют диатомовые водоросли, то есть отложения существенно водные. Однако есть и заметные различия. Так, для старичного озера наиболее характерны планктонные диатомеи, их на 10% больше, чем обрастателей и в два раза больше, чем донных. Кроме того, присутствует небольшое количество реофильных форм, а также губки. Напротив, в отложениях паводка преобладают обрастатели и донные, планктонные содержатся в незначительном количестве. Губки, которые хорошо вегетируют в спокойных проточных водах, не обнаружены. Но много реофильных диатомей – обитателей проточных водоемов. Цист золотистых водорослей и в отложениях поймы, и в отложениях старицы очень немного. Это значит, что воды материнских водоемов имели среднюю или высокую минерализацию и содержали малое количество железа.
Характер содержащихся в пробах остатков высших растений также примечателен. Процентное содержание макроостатков высших растений оказалось в 1.5 раза больше в отложениях паводка, но практически все они представлены древесной трухой, то есть мусором, который переносился рекой. А в отложениях старичного озера присутствуют хвощ, осоки и фрагменты корней травянистых растений. При этом кора хвойных, которые обильно произрастают на поверхности яра и начинают появляться на осыпях (рис. 3), встречена в единичных экземплярах. То есть на берегу спокойного водоема создаются условия для произрастания водно-болотной растительности, а мусорный занос минимален.
Выше указано, что для отложений холодноводных озер со значительной глубиной и большой площадью зеркала воды характерны заметные количества (10–15%) Cyclotella ocellata и Stephanodiscus minutulus. C. ocellata отмечена и в отложениях современной поймы, и в старичном озере, но в обоих случаях в незначительном количестве (1–1.2%); а представители Stephanodiscus не обнаружены.
На рис. 2, В представлена гистограмма процентного содержания диатомовых водорослей для верхнего слоя осадков озера Двухюрточного, построенная по данным из [26, fig. 6]. Хорошо видно, что подавляющее большинство диатомовых представлено планктоном (включая небольшое количество бентосных форм), а обрастатели составляют чуть более 10% пробы.
ЗАКЛЮЧЕНИЕ
Впервые методом комплексного группового биоанализа проведено сравнительное исследование современных отложений паводка и начальной стадии зарастания старичного озера (пойменной и старичной фаций аллювия) в среднем течении р. Камчатки. Выявлены некоторые особенности состава биоты для литологически плохо различимых отложений. Для отложений паводка характерно доминирование диатомей-обрастателей (в том числе много реофильных форм) и донных; губки не обнаружены; остатки высших растений представлены занесенным мусором (древесная труха). В отложениях начальной фазы зарастания старичного озера преобладают планктонные диатомеи, при этом много обрастателей, а около 20% приходится на долю донных видов. В небольших количествах отмечаются реофильные диатомеи и губки; остатки высших растений представлены водно-болотной растительностью (хвощ, осока). Cyclotella ocellata, которая, по данным литературных источников, является индикатором крупных и довольно глубоких озер с холодной водой, обнаружена и в отложениях паводка, и в старичном озере, но в незначительном количестве (менее 1.5%). Представители рода Stephanodiscus, часто встречающиеся в больших озерах, в исследованных пробах не установлены. В условиях кратковременного затопления поймы формируются особые биотические сообщества, состав которых заметно отличается от сообществ, развивающихся в долгоживущих глубоководных озерах. По данным биоанализа комплекс биоостатков в обоих рассматриваемых случаях идентифицируется как отложения водоема. Представляется важным подчеркнуть, что понятие “водоем” не всегда означает долгоживущее глубоководное озеро.
В настоящей публикации рассмотрены лишь две представительные пробы современных осадков из долины р. Камчатки. Для выработки уверенных критериев для распознавания отложений различного генезиса этого не достаточно. Однако самые яркие различия между отложениями паводка, локального старичного озера и глубокого озера с обширным зеркалом воды удалось установить. Это важная информация, которая позволит в дальнейшем лучше распознавать похожие с литологической точки зрения погребенные отложения позднеплейстоценового возраста.
Список литературы
Бонк Т.В., Лепская Е.В., Маркевич Г.Н. Планктонное сообщество озера Чаша (Камчатка) // Сохранение биоразнообразия Камчатки и прилегающих морей. Петропавловск-Камчатский, 2007. С. 22–24.
Бонк Т.В., Лупикина Е.Г. Современное состояние планктона озера Карымское (Камчатка) // Чтения памяти Владимира Яковлевича Леванидова. Владивосток: Дальнаука, 2011. Вып. 5. С. 69–72.
Брайцева О.А., Мелекесцев И.В., Евтеева И.С., Лупикина Е.Г. Стратиграфия четвертичных отложений и оледенения Камчатки. М.: Наука, 1968. 226 с.
Генкал С.И., Лепская Е.В. Центрические диатомовые водоросли вулканических Верхнеавачинских озер (Камчатка) // Биология внутренних вод. 2014. № 1. С. 5–13. https://doi.org/10.7868/S0320965214010069
Генкал С.И., Лупикина Е.Г., Лепская Е.В. Cyclotella tripartita (Bacillariophyta) из озер Камчатки и Забайкалья // Ботанический журн. 2004. Т. 89. № 3. С. 426–435.
Генкал С.И., Лепская Е.В., Лупикина Е.Г. Диатомовые водоросли озера Хангар (Камчатка) // Ботанический журн. 2007. Т. 92. № 10. С. 1500–1507.
Генкал С.И., Лупикина Е.Г. Новые и редкие виды Aulacosira (Bacillariophyta) из кальдерных озер Камчатки // Ботанический журн. 1998. Т. 83. № 2. С. 104–110.
Кордэ Н.В. Биостратификация и типология русских сапропелей. М.: Изд-во АН СССР, 1960. 219 с.
Куликовский М.С., Шкурина Н.А. Видовой cостав и особенности флоры центрических диатомовых водорослей (Bacillariophyta) водоемов и водотоков Камчатки // Поволжский экологический журн. 2009. № 4. С. 305–318.
Куприна Н.П. Стратиграфия и история осадконакопления плейстоценовых отложений Центральной Камчатки. М.: Наука, 1970. 148 с.
Лепская Е.В. Фитопланктон оз. Ажабачье и его роль в питании массовых видов зоопланктона // Исследование биологии и динамики численности промысловых рыб камчатского шельфа. Петропавловск-Камчатский: Камчатский печатный двор, 2000. Вып. 5. С. 152–160.
Лепская Е.В. Многолетняя динамика численности и биомассы фитопланктона озера Курильское и определяющие ее факторы // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Петропавловск-Камчатский, 2004. Вып. 7. С. 79–87.
Лепская Е.В. Фитопланктон озера Паланского. Таксономия, экология, динамика количественных характеристик // Исследование водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Петропавловск-Камчатский, 2007. Вып. 9. С. 10–20.
Лепская Е.В., Рассел Ч. Доминантная флора лососевых озер юга Камчатки (Россия) // Альгология. 1999. Т. 9. № 2. С. 73.
Лепская Е.В., Лупикина Е.Г., Маслов А.В., Уколова Т.К., Свириденко В.Д. К характеристике альгофлоры пелагиали некоторых озер Камчатки // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука, 2002. Вып. 2. С. 272–286.
Максимов Ф.Е., Певзнер М.М., Петров А.Ю., Левченко С.Б., Григорьев В.А., Баранова Н.Г., Кузнецов В.Ю. Возраст толщи “косослоистых песков” опорного разреза яр Средний (Центральная Камчатка) по данным комплексного 230Th/238U и 14С датирования торфа // Доклады РАН. 2019. Т. 488. № 3. С. 288–293. https://doi.org/10.31857/S0869-56524883288-293
Маркевич Г.Н., Лепская Е.В., Исаев В.А., Бонк Т.В., Свириденко В.Д., Чебанова В.В., Генкал С.И., Введенская Т.Л. Природные условия, микрофлора и фауна Верхнеавачинских озер (Камчатка) // Известия ТИНРО. 2011. Т. 164. С. 312–329.
Певзнер М.М., Яшина О.В., Смышляева О.И., Нечушкин Р.И., Каримов Т.Д., Рождественский О.Ю. Опорные разрезы рыхлых отложений позднего-среднего плейстоцена Камчатки: современное состояние, проблема возраста и перспективы изучения // Литология и полезные ископаемые. 2019. № 1. С. 60–89. https://doi.org/10.31857/S0024-497X2019160-89
Певзнер М.М., Каримов Т.Д., Нечушкин Р.И. Толща “синих глин” Центральной Камчатки: возраст и проблема происхождения // Вулканизм и связанные с ним процессы. Петропавловск-Камчатский, 2019. С. 102–105.
Успенская О.Н. Другие водоросли // Общие закономерности возникновения и развития озер. Методы изучения истории озер. Сер. История озер СССР. Л.: Наука, 1986. С. 146–151.
Черепанова М.В., Лепская Е.В., Андерсон П., Ложкин А.В. Диатомовые водоросли из голоценовых осадков оз. Нерпичье (Камчатка) // Исследования водных биологических ресурсов Камчатки и Северо-Западной части Тихого океана. Петропавловск-Камчатский, 2013. Вып. 31. С. 89–106.
Шкурина Н.А., Лепская Е.В., Белякова Г.А. Диатомовые водоросли озера Дальнее (Камчатка) // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Петропавловск-Камчатский, 2004. Вып. 7. С. 88–93.
Algae Base. https://www.algaebase.org/
Dirksen V.G., Uspenskaya O.N. The Holocene climate and vegetation changes in Eastern Kamchatka based on pollen, macrofossil and tephra’s data // Second Scientific Congress of East asian Federation of Ecological Societies “Global Environmental Change and Ecosystems in East Asia”. March 25–28, 2006. Niigata, Japan. Abstract ES9-06.
Hoff U., Dirksen O., Dirksen V., Herzschuh U., Hubberten H.-W., Meyer H., van den Bogaard Ch., Diekmann B. Late Holocene diatom assemblages in a lake-sediment core from Central Kamchatka, Russia // J. Paleolimnol. 2012. V. 47(4). P. 549–560. https://doi.org/10.1007/s10933-012-9580-y
Hoff U., Biskaborn B., Dirksen V., Dirksen O., Kuhn G., Meyer H., Nazarova L., Roth A., Diekmann B. Holocene environment of Central Kamchatka, Russia: Implication from a multi-proxy record of Two-Yurts Lake // Global and Planetary Change. 2015. V. 134. P. 101–117. https://doi.org/10.1016/j.gloplacha.2015.07.011
Hoff U., Dirksen O., Dirksen V., Kuhn G., Meyer H., Diekmann B. Holocene freshwater diatoms: palaeoenvironmental implications from south Kamchatka, Russia // Boreas. 2014. V. 43. P. 22–41. https://doi.org/10.1111/bor.12019
Krammer K., Lange-Bertalot H. Bacillariophyceae. 1: Naviculaceae // Sußwasserflora von Mitteleuropa. Stuttgard. N.Y.: VEB Gustav Fischer Verlag, 1986. Bd. 2. 876 p. (in German).
Krammer K., Lange-Bertalot H. Bacillariophyceae. 2: Bacillariaceae, Epithemiaceae, Surirellaceae // Sußwasserflora von Mitteleuropa. Jena: VEB Gustav Fischer Verlag, 1988. Bd. 2. 536 p. (in German).
Krammer K., Lange-Bertalot H. Bacillariophyceae. 3: Centrales; Fragilariaceae, Eunotiaceae // Suwasserflora von Mitteleuropa. Stuttgard, Jena: VEB Gustav Fischer Verlag, 1991a. Bd. 2. 576 p. (in German).
Krammer K., Lange-Bertalot H. Bacillariophyceae. 4: Ahnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverreichnis // Sußwasserflora von Mitteleuropa. Stuttgard, Jena: VEB Gustav Fischer Verlag, 1991b. Bd. 2. 437 p. (in German).
Skrypnikova M., Uspenskaya O., Khokhlova O. Paleoclimate study of mountain ecosystems by multiple group biological analysis // J. Mountain Sci. 2011. V. 43. № 1. P. 24–36. https://doi.org/10.1007/s11629-011-1033-y
Дополнительные материалы отсутствуют.